鄱阳湖湿地灰化苔草固碳能力及固碳量研究
2017-12-29周云凯白秀玲宁立新
周云凯,白秀玲*,宁立新
1. 河南大学环境与规划学院,河南 开封 475004;2. 北京师范大学减灾与应急管理研究院,北京 100875
鄱阳湖湿地灰化苔草固碳能力及固碳量研究
周云凯1,白秀玲1*,宁立新2
1. 河南大学环境与规划学院,河南 开封 475004;2. 北京师范大学减灾与应急管理研究院,北京 100875
湿地植物具有较强的固碳功能,在全球碳循环过程中扮演重要角色。利用实地测量数据,结合遥感(RS)与地理信息系统(GIS)技术研究鄱阳湖湿地洲滩优势植物——灰化苔草(Carex cinerascens)的固碳功能及其空间分异,以期为正确认识鄱阳湖湿地植物在区域生态系统碳循环中的作用,制定合理有效的湿地生态管理与保护对策提供科学依据。结果表明:在灰化苔草年内两个生长季,春草的平均固碳能力(579.46±294.19) g∙m-2略低于秋草(629.65±155.85) g∙m-2,不同高程带的固碳能力均以13.5~14.5 m高程带最大,而12.5 m以下高程带最小,且各高程带间(除秋草的12.5~13.5 m与>14.5 m高程带间)固碳能力均达到了极显著差异(P<0.001);灰化苔草春草和秋草的固碳量基本相当,分别为44.05×104t和44.10×104t;但由于受不同生长季各高程带分布面积和生物量变动的影响,灰化苔草春草生长季内最大固碳量出现在 13.5~14.5 m 高程带(19.84×104t),最小固碳量出现在<12.5 m 高程带(4.41×104t),而秋草生长季内最大固碳量则出现 12.5~13.5 m 高程带(17.29×104t),最小固碳量出现在>14.5 m高程带(4.51×104t);从全年来看,灰化苔草年固碳量为88.15×104t,年均固碳能力可达到1103~1180 g∙m-2,远高于其生长季内的碳释放能力,是鄱阳湖湿地的一个重要碳汇。
固碳能力;固碳量;空间分异;灰化苔草;鄱阳湖湿地
湿地是陆地生态系统的重要组成部分,在全球碳循环中扮演着重要角色(孟伟庆等,2011)。一方面湿地通过储存有机碳和无机碳而具有碳汇特征,而另一方面湿地又通过释放CO2、CH4等温室气体而具有碳源特征(宋洪涛等,2011),因此,湿地具有碳汇和碳源的双重功能。从短时间(<60年)来看,湿地有可能会是温室气体的源,但长期(>100年)来看,湿地则是温室气体的汇(王淑琼等,2014;Joiner et al.,1999;Brix et al.,2001;王德宣,2003)。在全球陆地生态系统中,湿地面积虽然仅占陆地总面积的 4%~6%,但碳储量却占到了全球陆地生态系统的12%~24%(IPCC,2001;吕铭志等,2013),其碳汇能力远高于农业、草原、温带森林和热带雨林等生态系统,是全球陆地生态系统最大的碳库(Mitsch et al.,2015;段晓男等,2006;杨平等,2011)。
植物作为湿地的主要初级生产者,可通过光合作用固定大气中的CO2形成植物有机体,是湿地碳积累的主要来源之一,发挥着重要的储碳、固碳功能(李博等,2009;宋洪涛等,2011)。然而,不同的湿地植物类型,其固碳能力存在着一定差异,而同种湿地植物生长在不同的环境中或不同的环境梯度下(如不同的水分条件、盐分条件、养分条件等),其生长发育过程也会有所不同,导致植物的生物量和固碳功能出现内部差异(Liu et al.,2015;Lou et al.,2016)。当前研究更多地关注植被类型间固碳功能的差别(梅雪英等,2007;张绪良等,2012;邵学新等,2013;潘宝宝等,2014),而忽视了植物群落或种群内部因微环境变化(尤其是高程形成的水分梯度)引起的固碳功能的空间分异,这在一定程度上降低了湿地植物固碳能力和固碳量的估算精度。因此,有效获取湿地植物空间分布以及分异信息对于准确评估湿地植物的固碳功能至关重要。近些年来,遥感(RS)与地理信息系统(GIS)技术的发展为植被空间信息获取提供了有力的技术支撑,结合实测数据,可实现对特定植物固碳功能更为精确的评估。
鄱阳湖湿地是中国最大的淡水湖泊湿地,由于其优越的水热条件及与长江复杂的水情关系,发育了区系成分复杂、多样性丰富的湿地植物群落(张全军等,2013;谭志强等,2016)。其中,灰化苔草(Carex cinerascens)是鄱阳湖湿地洲滩最主要的建群种之一(冯文娟等,2016),在湿地植物群落建造和生态系统功能维持方面起着重要作用(刘扬等,2016)。本文选取鄱阳湖湿地灰化苔草作为研究对象,基于RS与GIS技术,同时结合野外实测数据,研究灰化苔草的固碳能力、固碳量及其空间分异特征,旨在为正确认识鄱阳湖湿地植物在区域生态系统碳循环中的作用,制定合理有效的湿地生态管理与保护对策提供科学依据。
1 材料与方法
1.1 研究区概况
鄱阳湖湿地位于江西省北部,长江中下游南岸,地理坐标为 24°29′14″~30°04′41″N、113°34′36″~118°28′58″E。受长江和流域“五河”(赣江、抚河、饶河、修水、信江)来水的共同影响,鄱阳湖水位年内变化剧烈,呈现出丰水期与枯水期交替出现的独特水文节律(戴雪等,2014)。该区在气候上属于亚热带季风气候,夏季高温多雨,冬季温和少雨,多年平均气温 16.5~17.8 ℃,平均降水量 1350~1700 mm(刘肖利等,2013)。由于具有较为充足的水热资源,鄱阳湖湿地成为我国生物多样性最为丰富的地区之一(游海林等,2013)。随着水位的周期性涨落变化,鄱阳湖湿地洲滩各高程带“淹没—出露”过程不一,这种不同的水文经历使洲滩植物沿高程梯度呈现出较为明显的带状分布格局(胡振鹏等,2010),从湖岸向湖心方向依次分布着芦苇(Phragmites australis)、南荻(Triarrhena lutarioriparia)、灰化苔草(Carex cinerascens)和虉草(Phalaris arundinacea)等植物群落(周云凯等,2017),其中灰化苔草是鄱阳湖湿地分布最广的洲滩优势植物种(吴琴等,2012),是多年生根茎型湿生植物,一年中有两个生长季,分为春草和秋草。春草一般在每年 2—5月萌发生长,丰水期被湖水淹没,大量死亡或进入休眠状态,秋草则在秋季退水期洲滩出露后开始萌芽,一般9—12月为萌发生长期,次年1—2月枯萎(胡振鹏等,2010)。灰化苔草占据着一定的洲滩高程区间,各高程带由于水分条件等环境梯度的影响,其生长发育状况不尽相同。
1.2 研究方法
1.2.1 生物量测定
本研究选取灰化苔草分布集中的蚌湖东侧修水入湖口洲滩作为野外观测样地,洲滩上植物沿高程呈现出较为明显的带状分布,由低到高依次分布着虉草、灰化苔草、南荻等植物群落,其中灰化苔草面积最大,生长也较为茂盛。分别于 2013年 4月下旬(春草生长季)和10月下旬(秋草生长季)在洲滩两侧与湖岸垂直方向(沿高程或水分梯度)各布设1条样带,在样带灰化苔草分布区内按不同高程(<12.5m、12.5~13.5 m、13.5~14.5 m 和>14.5 m)布设采样点,在每条样带的同一高程带内,按等距离设立3个面积为1 m×1 m的样方,因此,每个高程带共布设6个样方,总计24个样方,用GPS-RTK(南方测绘,灵锐S86)确定各样方的高程和地理位置。在每个样方内采用计数法确定灰化苔草的种群密度,然后按对角线法选取9株灰化苔草植株,用剪刀齐地面剪下其地上部分(包括已枯叶片),并用小铁锹挖掘其地下根系,分离出新生根,将其装入密封袋中,共采集 24个植物样品。将采集到的灰化苔草植物样品带回实验室后用清水冲洗干净,并用吸水纸吸干,之后将其置于鼓风干燥箱中于105 ℃下杀青10 min,再在80 ℃下烘干至恒重,用精度为0.01 g的电子天平分别称量地上与地下部分干重,两者相加计算出总生物量,并换算成单位面积生物量,最后求各高程带及总体的平均值和标准差,以平均值±标准差作为灰化苔草生物量的统计值。
1.2.2 分布面积提取
从地理空间数据云网站(http://www.gscloud.cn/)收集获取2013年5月14日(春草生长季)和2013年10月5日(秋草生长季)两景Landsat8-OLI遥感影像。借助ENVI 5.0软件对遥感影像进行波段合成;以1∶50000地形图作为参考图像对遥感影像进行几何精校正;利用鄱阳湖湿地范围图作为掩膜对合成后的遥感影像进行裁剪;依据影像的光谱特征及计算得到的NDVI值,采用决策树分类法对遥感影像进行解译,对解译结果进行人机交互式目视检查,并纠正错分区域,确保遥感影像解译精度达到80%以上,从而得到灰化苔草春草和秋草的空间分布信息。使用解译后得到的两期灰化苔草分布范围矢量数据作为掩模,借助ARCGIS 10软件裁切鄱阳湖湿地DEM栅格数据,利用ARCGIS空间分析(Spatial Analyst)模块中的重分类(Reclassify)功能对裁切后的DEM按高程进行分级,将不同高程级别的栅格个数乘以像元大小,即可获得灰化苔草春草和秋草不同高程带的分布面积。
1.2.3 固碳能力计算
2013年4月下旬和10月下旬灰化苔草春草和秋草分别接近成熟期,其现存生物量基本可视为相应生长季生物量的累积量,可将其作为植物的净初级生产力(索安宁等,2010)。在生物量调查的基础上,根据光合作用反应式,草本植物每生成 1 g干物质平均需要固定1.62 g CO2,在此基础上乘以碳的质量比 0.273,可得到碳的转换系数(王淑琼等,2014)。利用各高程带实地测量得到的生物量乘以碳转换系数,可求得不同高程带灰化苔草的固碳能力(以平均值±标准差表示);然后再乘以遥感影像解译后得到的各高程带灰化苔草分布面积,可进一步估算出不同高程带灰化苔草的固碳量。计算公式为(索安宁等,2010;Daniel et al.,2002):
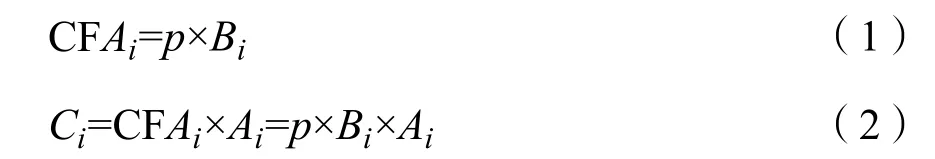
式中,CFAi为第i高程带灰化苔草的固碳能力(g∙m-2);Ci为第 i高程带灰化苔草的固碳量(t);p为碳转换系数(0.442),Bi为第 i高程带灰化苔草平均净初级生产力(以平均生物量代替)(g∙m-2);Ai为第i高程带灰化苔草的分布面积(km2)。
1.2.4 数据统计分析
运用SPSS 19.0对测量与计算数据进行统计分析,采用单因素方差分析(One-way ANOVA)中的LSD多重比较法进行不同高程带生物量、固碳能力的差异显著性检验。其中,P>0.05表示差异未达显著水平;P<0.05为差异达显著水平;而 P<0.01为差异达极显著水平。
2 结果与分析
2.1 灰化苔草空间分布
遥感影像解译结果显示(图 1),2013年灰化苔草主要分布在鄱阳湖湿地的西部和南部区域,而在鄱阳湖湿地的北部与东部区域灰化苔草分布极少,只在部分出露的洲滩上有少量分布。其中,春草主要分布在湿地的西部和西南部区域,而秋草更多地分布在湿地的南部区域。从分布面积来看(表1),2013年鄱阳湖湿地灰化苔草春草分布面积为798.80 km2,占鄱阳湖湿地总面积的26.3%,14.5 m以下高程带是其主要分布区间,占总分布面积的86.3%;秋草分布面积略少于春草,为746.55 km2,占鄱阳湖湿地总面积的24.6%。就不同高程带分布而言,12.5 m以下高程带的分布面积较春草有所增加,而14.5 m以上高程带的分布面积较春草则有所减少,整体分布高程略有下移。

表1 灰化苔草年内不同生长季各高程带分布面积Table1 The distribution area of Carex cinerascens in different elevation zones of Poyang Lake wetland during the spring and autumn growth seasons km2
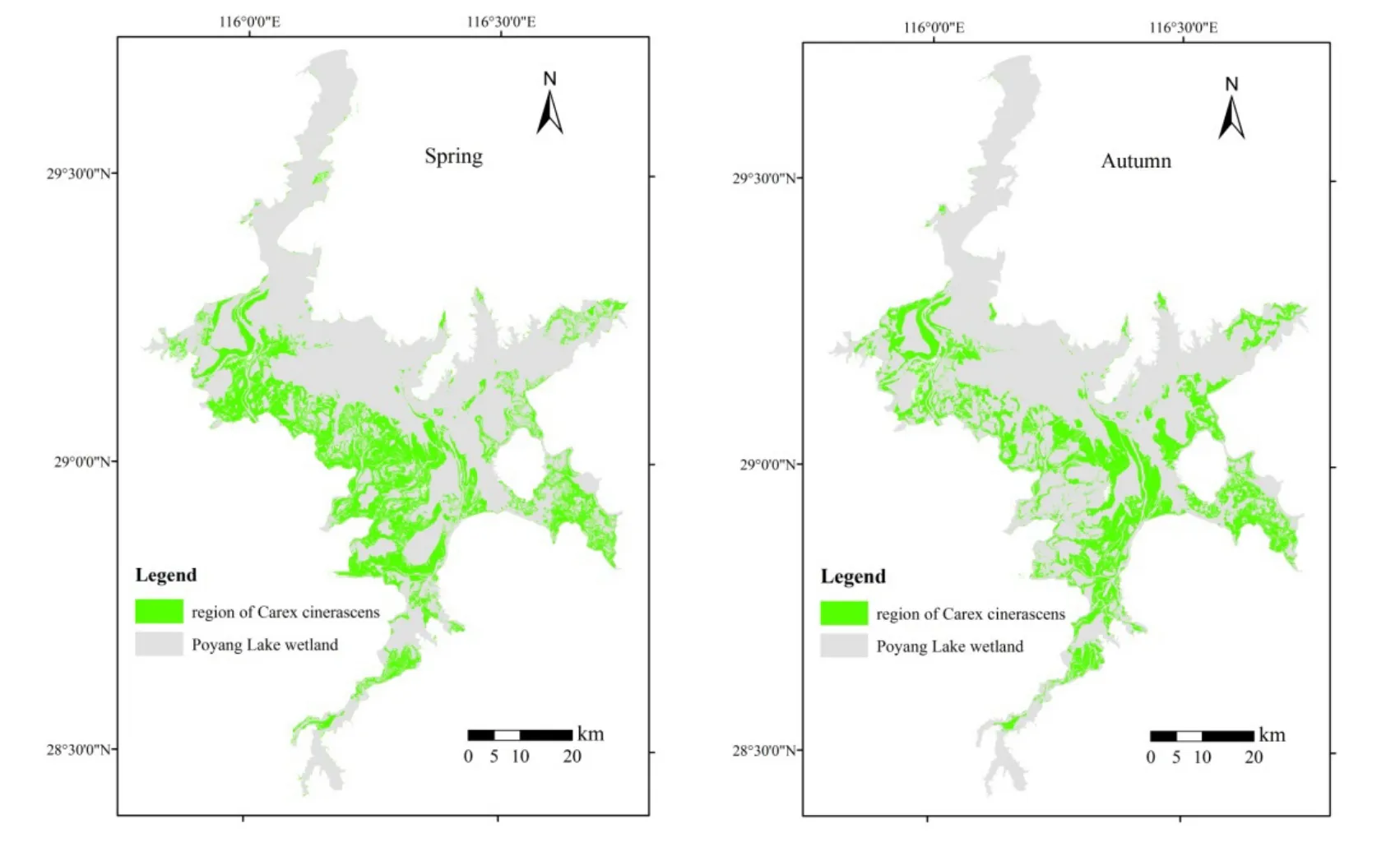
图1 2013年鄱阳湖湿地灰化苔草春草与秋草分布Fig.1 The spatial distribution maps of Carex cinerascens in Poyang Lake wetland in spring and autumn growth seasons in 2013
2.2 灰化苔草生物量
从年内两个生长季相同生长期生物量(干质量)测定结果来看(图2),鄱阳湖湿地灰化苔草春草的平均生物量(1311.0±665.59) g∙m-2略低于秋草(1424.5±352.59) g∙m-2,两个生长季内均以 13.5~14.5 m 高程带生物量为最高,春草为(2240.00±49.05)g∙m-2,秋草为(1760.00±35.66) g∙m-2;而 12.5 m 以下高程带生物量最低,春草为(483.80±20.03) g∙m-2,秋草为(848.80±30.50) g∙m-2。但不同生长季各高程带间生物量差异有所不同,在春草生长季内,不同高程带间生物量差异都达到了极显著检验水平(P<0.001);而在秋草生长季内,除了12.5~13.5 m与>14.5 m高程带间生物量差异不显著(P=0.169)外,其他高程带间生物量也都达到了极显著差异(P<0.001)。
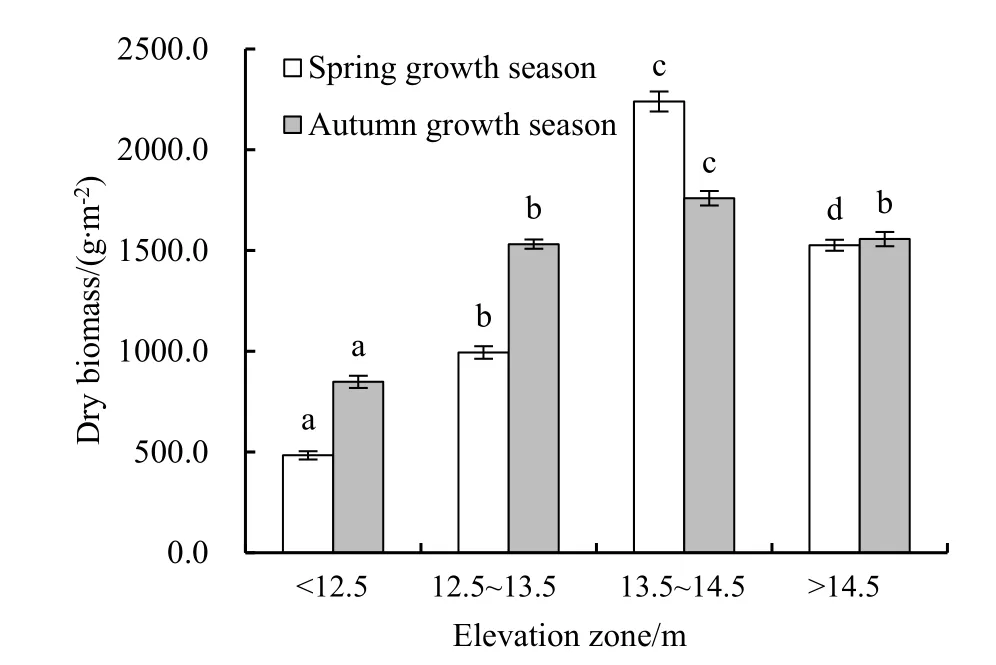
图2 不同高程带灰化苔草生物量(干重)Fig.2 The dry biomass of Carex cinerascens in different elevation zones of Poyang Lake wetland
2.3 灰化苔草固碳能力与固碳量
根据式(1)、(2)计算得到灰化苔草各高程带春草与秋草的固碳能力与固碳量(表2)。由表2可知,鄱阳湖湿地灰化苔草春草平均固碳能力(579.46±294.19) g∙m-2略低于秋草(629.65±155.85)g∙m-2,两个生长季内灰化苔草的固碳能力均以13.5~14.5 m高程带最大,而12.5 m以下高程带最小;但在春草生长季,各高程带间固碳能力均达到了极显著差异(P<0.001),而在秋草生长季,除12.5~13.5 m 与>14.5 m 高程带间差异不显著(P=0.169)外,其他高程带间也均达到了极显著差异(P<0.001)。
固碳能力与分布面积决定着灰化苔草的固碳量。虽然春草固碳能力略低于秋草,但由于春草的分布面积(798.80 km2)大于秋草(746.55 km2),尤其在固碳能力较强的13.5~14.5 m和>14.5 m高程带,春草的分布面积分别是秋草的 1.27倍和 1.67倍。因此,两个生长季内灰化苔草的固碳量基本相当,分别为 44.05×104t和 44.10×104t(表 2)。而各高程带的固碳量在不同生长季发生一定变动,春草生长季内最大固碳量出现在 13.5~14.5 m 高程带(19.84×104t),最小固碳量出现在<12.5 m 高程带(4.41×104t);而秋草生长季内最大固碳量则出现12.5~13.5 m 高程带(17.29×104t),最小固碳量出现在>14.5 m高程带(4.51×104t)(表 2)。此外,高程带间固碳能力和固碳量变幅均表现为春草>秋草。从全年来看,灰化苔草年固碳量为88.15×104t,年均固碳能力可达到 1103~1180 g∙m-2。
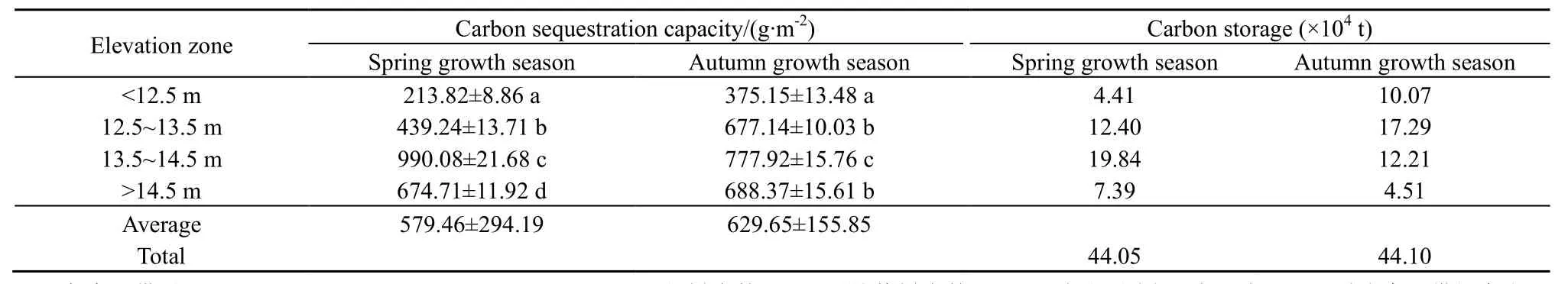
表2 不同高程带灰化苔草固碳能力与固碳量Table2 The carbon sequestration capacity and carbon storage of Carex cinerascens in different elevation zones of Poyang Lake wetland
3 讨论
鄱阳湖湿地灰化苔草的年均固碳能力明显高于全球陆地植被的平均固碳能力(表3),也高于中国陆地植被的平均固碳能力,分别是中国湖泊、河流、沼泽等生态系统年均固碳能力的7.5、5.03、1.82倍以上。与中国其他地区典型湿地植物相比,鄱阳湖湿地灰化苔草的年均固碳能力高于洪湖湿地菰(Zizania latifolia)和黄河三角洲湿地芦苇,也高于杭州湾和崇明东滩湿地的海三棱藨草(Scirpus mariqueter)的固碳能力,而与白洋淀湿地芦苇固碳能力相当,但低于杭州湾和崇明东滩湿地芦苇的年均固碳能力。可见,鄱阳湖湿地灰化苔草具有较强的固碳能力。值得注意的是,灰化苔草固定的碳并不能全部保存下来,一部分会通过植物的呼吸作用、微生物的分解作用以CO2、CH4等温室气体的形式释放到环境中,表现出碳源功能。已有研究发现,鄱阳湖苔草湿地在非淹水期 CO2释放量为205.99 g∙m-2,而 CH4释放量约为 0.12 g∙m-2(胡启武等,2011),其总体的碳释放能力远小于灰化苔草各生长季的碳固定能力。综合来看,灰化苔草在生长季内是鄱阳湖湿地的一个重要碳汇。
在鄱阳湖湿地,受水文条件等环境因素的影响,植物沿高程呈较为明显的带状分布特点,不同的湿地植物占据着一定的高程区间(胡振鹏等,2010),而在不同的生长季,由于环境条件的改变,植物分布的高程区间也会发生变动。在同一植物分布带中,由于各高程带的水分条件和水文经历不同,植物的生长发育状况不尽相同,表现出不同的生态和生理特征(王丽等,2008)。灰化苔草是鄱阳湖湿地典型的洲滩优势植物,2013年灰化苔草春草和秋草主要分布在12~16 m高程带上,但由于鄱阳湖水位的节律性涨落变化,不同高程带水文条件差异明显,这对灰化苔草生长发育状况具有重要影响。13.5 m以下高程带在灰化苔草生长季内长期处于过度湿润或淹水状态,使得灰化苔草生长受到抑制,其生物量和固碳能力相对较低,14.5 m以上高程带则相对干旱,灰化苔草生长同样会受到水分相对缺乏的限制,而13.5~14.5 m高程带在生长季内处于中度湿润或间歇性半淹水状态,形成有利于灰化苔草生长的水文条件,其生物量和固碳能力达到最高。灰化苔草生物量和固碳能力出现的这种空间分异是灰化苔草对鄱阳湖水位年内节律性变化长期适应的结果。
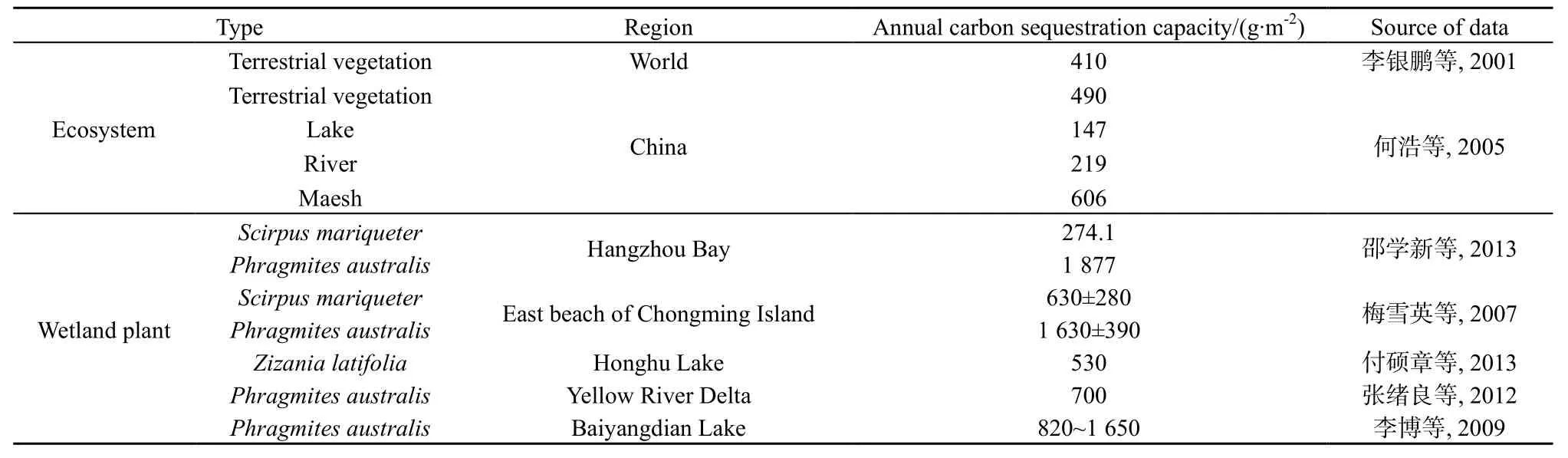
表3 不同生态系统与湿地植物固碳能力Table3 Carbon sequestration capacity of different ecosystems and wetland plants
4 结论
鄱阳湖湿地灰化苔草作为洲滩优势植物种,具有较强的固碳能力,通过对比发现,灰化苔草固碳能力高于全球和中国陆地植被的平均固碳能力,也高于许多地区典型湿地植物的固碳能力。在两个生长季内,灰化苔草春草的平均固碳能力(579.46±294.19) g∙m-2略低于秋草(629.65±155.85) g∙m-2,不同高程带的固碳能力均以13.5~14.5 m高程带最大,而12.5 m以下高程带最小;在春草生长季,各高程带间固碳能力均达到了极显著差异(P<0.001),而在秋草生长季,除12.5~13.5 m与>14.5 m高程带间差异不显著(P=0.169)外,其他高程带间也均达到了极显著差异(P<0.001)。
灰化苔草春草和秋草的固碳量基本相当,分别为44.05×104t和44.10×104t。但由于受灰化苔草分布面积和生物量变动的影响,各高程带固碳量在不同生长季间存在一定变动,春草生长季内最大固碳量出现在 13.5~14.5 m高程带(19.84×104t),最小固碳量出现在<12.5 m高程带(4.41×104t);而秋草生长季内最大固碳量则出现 12.5~13.5 m 高程带(17.29×104t),最小固碳量出现在>14.5 m 高程带(4.51×104t)。从全年来看,灰化苔草年固碳量为88.15×104t,年均固碳能力可达到 1103~1180 g∙m-2,远高于其生长季内的碳释放能力,是鄱阳湖湿地的一个重要碳汇,在区域碳循环过程中发挥重要作用。
BRIX H, SORRELL B K, LORENZEN B. 2001. Are Phragmites-dominated wetlands a net sink of greenhouse gases? [J]. Aquatic Botany, 69(2-4):313-324.
DANIEL P R, STAVROULA S, GARY P, et al. 2002. Patterns of canopy-air CO2concentration in a brackish wetland: analysis of a decade of measurements and the simulated effects on the vegetation[J]. Agricultural and Forest Meteorology, 114(1-2): 59-73.
IPCC. 2001. Climate Change 2001: Synthesis report: Third assessment report of the intergovernmental panel on climate change [R].Cambridge: Cambridge University Press.
JOINER D W, LAFLEUR P M, MCCAUGHEY J H, et al. 1999.Interannual variability in carbon dioxide exchanges at a boreal wetland in the BOREAS northern study area [J]. Journal of Geophysical Research: Atmospheres, 104(D22): 27663-27672.
LIU Y, WANG L, LIU H, et al. 2015. Comparison of carbon sequestration ability and effect of elevation in fenced wetland plant communities of the Xilin River floodplains: a model case study [J]. River Research &Application, 31(7): 858-866.
LOU Y J, PAN Y W, GAO C Y, et al. 2016. Response of plant height,species richness and aboveground biomass to flooding gradient along vegetation zones in floodplain wetlands, Northeast China [J]. Plos One,11(4): e0153972.
MITSCH W J, GOSSELINK J G. 2015. Wetlands (5thEdition) [M].Hoboken (NJ): John Wiley & Sons.
戴雪, 万荣荣, 杨桂山, 等. 2014. 鄱阳湖水文节律变化及其与江湖水量交换的关系[J]. 地理科学, 34(12): 1488-1496.
段晓男, 王效科, 尹弢, 等. 2006. 湿地生态系统固碳潜力研究进展[J].生态环境, 15(5): 1091-1095.
冯文娟, 徐力刚, 王晓龙, 等. 2016. 鄱阳湖洲滩湿地地下水位对灰化薹草种群的影响[J]. 生态学报, 36(16): 5109-5115.
付硕章, 柯文山, 陈世俭. 2013. 洪湖湿地野菰群落储碳、固碳功能研究[J]. 湖北大学学报(自然科学版), 35(3): 393-396.
何浩, 潘耀忠, 朱文泉, 等. 2005. 中国陆地生态系统服务价值测量[J].应用生态学报, 16(6): 1122-1127.
胡启武, 幸瑞新, 朱丽丽, 等. 2011. 鄱阳湖苔草湿地非淹水期CO2释放特征[J]. 应用生态学报, 22(6): 1431-1436.
胡振鹏, 葛刚, 刘成林, 等. 2010. 鄱阳湖湿地植物生态系统结构及湖水位对其影响研究[J]. 长江流域资源与环境, 19(6): 597-605.
李博, 刘存歧, 王军霞, 等. 2009. 白洋淀湿地典型植被芦苇储碳固碳功能研究[J]. 农业环境科学学报, 28(12): 2603-2607.
李银鹏, 季劲钧. 2001. 全球陆地生态系统与大气之间碳交换的模拟研究[J]. 地理学报, 56(4): 379-389.
刘肖利, 丁明军, 李贵才, 等. 2013. 鄱阳湖湿地植物群落沿高程梯度变化特征研究[J]. 人民长江, 44(5): 82-86.
刘扬, 施建敏, 边子星, 等. 2016. 鄱阳湖湿地灰化苔草群落物种多度分布格局沿水分梯度的变化[J]. 草业科学, 33(1): 19-26.
吕铭志, 盛连喜, 张立. 2013. 中国典型湿地生态系统碳汇功能比较[J].湿地科学, 11(1): 114-120.
梅雪英, 张修峰. 2007. 崇明东滩湿地自然植被演替过程中储碳及固碳功能变化[J]. 应用生态学报, 18(4): 933-936.
孟伟庆, 吴绽蕾, 王中良. 2011. 湿地生态系统碳汇与碳源过程的控制因子和临界条件[J]. 生态环境学报, 20(8-9): 1359-1366.
潘宝宝, 张金池, 冯开宇, 等. 2014. 洪泽湖典型水生植物群落碳储量[J]. 湿地科学, 12(4): 471-476.
邵学新, 李文华, 吴明, 等. 2013. 杭州湾潮滩湿地3种优势植物碳氮磷储量特征研究[J]. 环境科学, 34(9): 3451-3457.
宋洪涛, 崔丽娟, 栾军伟, 等. 2011. 湿地固碳功能与潜力[J]. 世界林业研究, 24(6): 6-11.
索安宁, 赵冬至, 张丰收. 2010. 我国北方河口湿地植物储碳、固碳功能研究——以辽河三角洲盘锦地区为例[J]. 海洋学研究, 28(3): 67-71.
谭志强, 张奇, 李云良, 等. 2016. 鄱阳湖湿地典型植物群落沿高程分布特征[J]. 湿地科学, 14(4): 506-515.
王德宣. 2003. 若尔盖高原与三江平原沼泽湿地CH4排放差异的主要环境影响因素[J]. 湿地科学, 1(1): 63-67.
王丽, 胡金明, 宋长青, 等. 2008. 水分梯度对三江平原典型湿地植物小叶章地上生物量的影响[J]. 草业学报, 17(4): 19-25.
王淑琼, 王瀚强, 方燕, 等. 2014. 崇明岛滨海湿地植物群落固碳能力[J]. 生态学杂志, 33(4): 915-921.
吴琴, 尧波, 朱丽丽, 等. 2012. 鄱阳湖典型苔草湿地生物量季节变化及固碳功能评价[J]. 长江流域资源与环境, 21(2): 215-219.
杨平, 仝川. 2011. LUCC对湿地碳储量及碳排放的影响[J]. 湿地科学与管理, 7(3): 56-59.
游海林, 徐力刚, 姜加虎, 等. 2013. 鄱阳湖典型洲滩湿地植物根系生长对极端水情变化的响应[J]. 生态学杂志, 32(12): 3125-3130.
张全军, 于秀波, 胡斌华. 2013. 鄱阳湖南矶湿地植物群落分布特征研究[J]. 资源科学, 35(1): 42-49.
张绪良, 张朝晖, 徐宗军, 等. 2012. 黄河三角洲滨海湿地植被的碳储量和固碳能力[J]. 安全与环境学报, 12(6): 145-149.
周云凯, 白秀玲, 宁立新. 2017. 鄱阳湖湿地苔草(Carex)景观变化及其水文响应[J].湖泊科学, 29(4): 870-879.
Carbon Sequestration Capacity and Carbon Storage of Carex cinerascens in Poyang Lake Wetland
ZHOU Yunkai1, BAI Xiuling1*, NING Lixin2
1. College of Environment and Planning, Henan University, Kaifeng 475004, China;2. Academy of Disaster Reduction and Emergency Management, Beijing Normal University, Beijing 100875, China
Wetland plants have strong ecological function in carbon fixation, and play an important role in the global carbon cycle.The main objective of the study is to understand the role of wetland plants in the regional ecosystem carbon cycle, which can provide scientific basis for wetland ecosystem management and reasonable protection in Poyang Lake wetland. Based on the field observation data and the technologies of remote sensing (RS) and geographic information system (GIS), this study investigates the carbon sequestration capacity, carbon storage and the spatial heterogeneity of Carex cinerascens in the Poyang Lake wetland. The results showed that the average carbon sequestration capacity of Carex cinerascens in spring growth season (579.46±294.19) g∙m-2was slightly lower than that in autumn growth season (629.65±155.85) g∙m-2. The carbon sequestration capacity showed great spatial heterogeneities in different growth seasons due to the influence of the environmental conditions caused by elevation. The elevation zone of 13.5~14.5 m had the strongest carbon sequestration capacity, whereas the elevation zone of <12.5 m was the weakest. In 2013, the carbon sequestration capacity among different elevation zones showed significant difference (P<0.001) except for two elevation zones between 12.5~13.5 m and >14.5m in autumn growth season (P=0.169). The total amount of carbon storage of Carex cinerascens was almost equal in spring and autumn growth seasons, but the elevation zones of the highest and lowest carbon storage had transferred from 13.5~14.5 m in spring growth season (19.84×104t) to 12.5~13.5 m in autumn growth season (17.29×104t) and from <12.5 m in spring growth season (4.41×104t) to >14.5 m in autumn growth season (4.51×104t), respectively. In the whole year,the carbon storage amount of Carex cinerascens can reach 88.15×104t, and the annual carbon sequestration capacity of Carex cinerascens was about 1103~1180 g∙m-2which is much higher than the carbon release capacity of Carex-dominated wetland. This implied that Carex cinerascens is an important carbon sink in Poyang Lake wetland in Jiangxi Province.
carbon sequestration capacity; carbon storage; spatial differentiation; Carex cinerascens; Poyang Lake wetland
10.16258/j.cnki.1674-5906.2017.12.005
X14
A
1674-5906(2017)12-2030-06
周云凯, 白秀玲, 宁立新. 2017. 鄱阳湖湿地灰化苔草固碳能力及固碳量研究[J]. 生态环境学报, 26(12): 2030-2035.
ZHOU Yunkai, BAI Xiuling, NING Lixin. 2017. Carbon sequestration capacity and carbon storage of Carex cinerascens in Poyang Lake wetland [J]. Ecology and Environmental Sciences, 26(12): 2030-2035.
国家自然科学基金项目(41371450;41101089);河南省高等学校青年骨干教师项目(2014GGJS-025)
周云凯(1980年生),男,副教授,博士,研究方向为湿地生态学。E-mail: ykzhou2009@126.com
*通信作者:白秀玲(1980年生),女,副教授,博士,研究方向为环境污染与生态修复。E-mail: bxling@163.com
2017-09-08
