橘小实蝇4种寄生蜂腹部和产卵器感器的扫描电镜观察
2017-12-25谷小红蔡普默杨燕川杨晴阳季清娥陈家骅
谷小红, 蔡普默, 杨燕川, 王 聪, 杨晴阳, 季清娥, 陈家骅
(福建农林大学植物保护学院益虫研究所,福建 福州 350002)
橘小实蝇4种寄生蜂腹部和产卵器感器的扫描电镜观察
谷小红, 蔡普默, 杨燕川, 王 聪, 杨晴阳, 季清娥, 陈家骅
(福建农林大学植物保护学院益虫研究所,福建 福州 350002)
应用扫描电镜对橘小实蝇的4种寄生蜂(阿里山潜蝇茧蜂、切割潜蝇茧蜂、长尾潜蝇茧蜂和布氏潜蝇茧蜂)的产卵器和腹部感器的形态和超微结构进行了观察和研究.共发现8种感器,分别为毛形感器、蒲姆氏鬃、钟形感器、浅凹状感器、栓锥感器、腔锥感器、刺形感器和分泌毛孔;同时发现阿里山潜蝇茧蜂和布氏潜蝇茧蜂的产卵针端部尖细,而长尾潜蝇茧蜂和切割潜蝇茧蜂的产卵针端部呈倒钩状.
寄生蜂; 橘小实蝇; 感器; 扫描电镜
橘小实蝇[Bactroceradorsalis(Hendel)],又名东方果实蝇,俗称果蛆,属双翅目Diptera实蝇科Tetriphitidae寡鬃实蝇亚科Dacinae果实蝇属BactroceraMacquart,是一种世界性检疫害虫[1-3],为害包括柑橘、芒果在内的46个科250多种果树、蔬菜和花卉,给我国果蔬业造成了巨大损失[4-5].寄生蜂是一类重要的寄生性天敌,其中许多种类是农林生态系统害虫综合治理中的重要力量.通过释放橘小实蝇的不同寄生蜂,可达到持续控制的目的[6-7].寄生蜂在生物防治中的成功应用依赖于其在复杂的化学与物理环境中寻找寄主的能力,有过学习经历的寄生蜂在田间释放后,可以通过气味更准确、快速地找到寄主和寄主植物,搜索效率和对寄主的寄生率较高,并且可以在田间停留更长的时间[8-9].
昆虫的感器在昆虫寻找寄主并决定是否在寄主上产卵等一系列行为中起着重要的作用.昆虫的感器分布在触角、足、腹部、产卵器等部位,其功能和形态各有不同[10].目前,我国学者已经对昆虫触角上的感受器超微结构进行了研究,但对寄生蜂产卵器感器的研究甚少.橘小实蝇等寄主通常隐藏在植物基质内,寄生蜂触角一般不易接触寄主,因此,产卵器的感受能力显得更为重要.由于寄生蜂种类及其寄主的不同,产卵器的外部形态及其感受器的结构也不相同,这与寄主利用和适应趋同现象有关[11].阿里山潜蝇茧蜂[Fopiusarisanus(Sonan)]、切割潜蝇茧蜂[Psyttaliaincise(Silvestri)]、长尾潜蝇茧蜂[Diachasmimorphalongicaudata(Ashmead)]和布氏潜蝇茧蜂[Fopiusvandenboschi(Fullaway)]是世界范围内防治橘小实蝇的重要寄生蜂.这4种寄生蜂对橘小实蝇不同虫期的寄生倾向性不同[12]:阿里山潜蝇茧蜂倾向于寄生橘小实蝇卵和初孵幼虫,切割潜蝇茧蜂倾向于寄生低龄幼虫,布氏潜蝇茧蜂多寄生于低、中龄幼虫,长尾潜蝇茧蜂则多寄生于高龄幼虫.已有学者对其触角感受器的超微结构进行了研究[13-14];但考虑到产卵器和腹部的感受器在寄生蜂对寄主的搜寻定位以及识别与接受过程中的重要作用,运用扫描电镜对其超微结构进行研究,以期为进一步了解不同寄生蜂对寄主选择差异性的机制提供依据,并为优化寄生蜂对橘小实蝇的生物防治效果提供参考.
1 材料与方法
1.1 供试虫源
阿里山潜蝇茧蜂和切割潜蝇茧蜂于2005年引进,长尾潜蝇茧蜂和布氏潜蝇茧蜂采自福建漳州,均在室内建立相对稳定的种群,约120代.切割潜蝇茧蜂、布氏潜蝇茧蜂和长尾潜蝇茧蜂分别寄生于橘小实蝇低、中、高龄幼虫,阿里山潜蝇茧蜂寄生于新鲜橘小实蝇卵或初孵幼虫.
寄生蜂幼虫饲养条件:温度(26±1) ℃,相对湿度为80%±5%,光周期L∶D=10 h∶14 h.用粗蔗糖、麦麸、废酵母等按一定比例配制的人工饲料饲养.
寄生蜂成虫饲养条件:饲养于25 cm×25 cm×25 cm木制饲养笼中,前侧中间有一个直径约15 cm的操作袖口,后侧安装活动玻璃门,其余各面用80目网纱封住.温度(25±1) ℃,相对湿度为75%±5%,光周期L∶D=14 h∶10 h.用蜂蜜和凉开水饲养.
1.2 样本制备和扫描电镜观察
取刚羽化的寄生蜂雌蜂各10头,置于100 Hz超声波中清洗30 s,除去表面黏附物.置于4 ℃冰箱内的2.5%戊二醛中固定5 h,用0.1 mol·L-1磷酸缓冲液和蒸馏水漂洗多次.在体视镜下,用解剖工具小心分离出产卵器和腹部,拍照并测量整体长度大小.
依次用30%、50%、70%、80%、90%、95%、100%的乙醇溶液脱水,置于干燥塔中干燥.将干燥好的样品贴于铜台上,真空环境下干燥喷金,在JSM6380LV扫描电子显微镜下观察、拍照.加速电压为15 kV.
1.3 试验仪器
JSM6380LV扫描电子显微镜(日本电子株式会社JEOL),SMZ-B4连续变倍体视显微镜(重庆奥特光学仪器有限公司),KQ2200DE型数控超声波清洗器(昆山市超声仪器有限公司),解剖刀和镊子.
2 结果与分析
2.1 产卵器和腹部的一般形态
4种潜蝇茧蜂的产卵器和腹部形态极为相似,产卵器皆由2片产卵鞘和内部的产卵针组成,由产卵鞘包裹着内部的产卵针,产卵时两边产卵鞘拄在寄主体外,产卵针伸长并插入寄主体内产卵(图1A).长尾潜蝇茧蜂产卵器比其他寄生蜂略长,切割潜蝇茧蜂的产卵器在这4种寄生蜂中最短.产卵鞘呈黑褐色、丝状,具有横纹(图1F),端部呈箭头状,具有锯齿状凸起(图1B),这结构能够加大摩擦力,有利于产卵针的固定;另有研究表明,其可能与寄主识别和避免过寄生现象发生有关[15].产卵针比产卵鞘细长,呈黄褐色且光滑.产卵针由产卵瓣组成,并由背产卵瓣和腹产卵瓣包围形成产卵道,背产卵瓣上具脊(图1C);不同的是,阿里山潜蝇茧蜂和布氏潜蝇茧蜂的产卵针端部尖细(图1D),长尾潜蝇茧蜂和切割潜蝇茧蜂的端部呈倒钩状(图1E).腹部一般为椭圆形.4个种间差异见表1.
2.2 产卵器和腹部感器的种类
2.2.1 毛形感器(sensilla trichoidea, ST) 毛形感器是4种茧蜂产卵器和腹部分布最广、数量最多的感受器.毛形感器主要分布于产卵鞘和腹部,产卵针并未发现毛形感器.按照形状和大小将该感器分为毛形感器Ⅰ和毛形感器Ⅱ(图2A).
毛形感器Ⅰ是产卵鞘和腹部分布数量最多的感器.产卵鞘的毛形感器Ⅰ向着产卵器端部方向着生,与产卵鞘呈约30°角,着生在凸起的基窝内,由基部向端部逐渐变细变尖,端部略弯曲,表面有明显纵纹.腹部的毛形感器Ⅰ分布于背面,呈有规则的横向排列,向产卵器方向着生,有基窝,由基部向端部逐渐变细变尖,端部略弯曲,表面有浅纵纹.毛形感器Ⅰ长23.02~106.18 mm,平均直径约3.02 mm.
毛形感器Ⅱ与毛形感器Ⅰ形状相似,但更短、细、小,分布于产卵鞘内侧边缘,较柔软,表面光滑,无纵纹.

A.产卵器(SS为产卵针,OS为产卵鞘);B.产卵鞘端部;C.产卵针(VV为腹产卵瓣,DV为背产卵瓣,R为背产卵瓣上的脊);D.阿里山潜蝇茧蜂和布氏潜蝇茧蜂的产卵针端部;E.长尾潜蝇茧蜂和切割潜蝇茧蜂的产卵针端部;F.横纹.图1 产卵器的基本形态结构Fig.1 General morphology of ovipositor
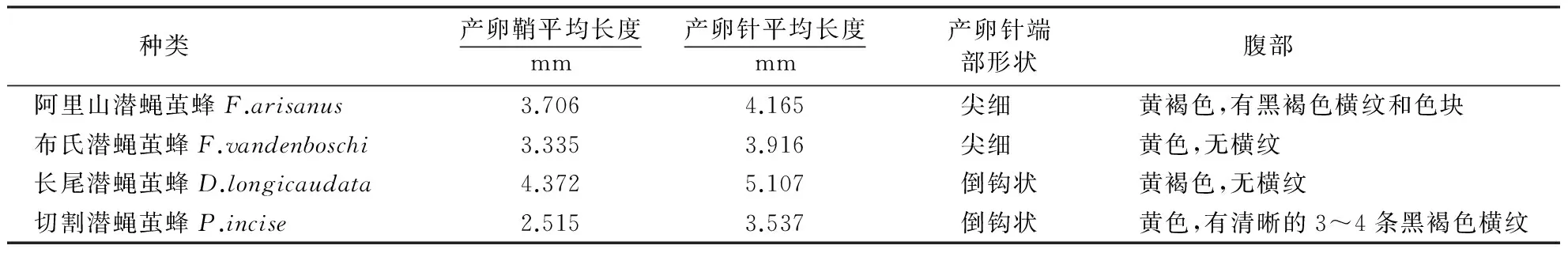
种类产卵鞘平均长度mm产卵针平均长度mm产卵针端部形状腹部阿里山潜蝇茧蜂F.arisanus3.7064.165尖细 黄褐色,有黑褐色横纹和色块布氏潜蝇茧蜂F.vandenboschi3.3353.916尖细 黄色,无横纹长尾潜蝇茧蜂D.longicaudata4.3725.107倒钩状黄褐色,无横纹切割潜蝇茧蜂P.incise2.5153.537倒钩状黄色,有清晰的3~4条黑褐色横纹
2.2.2 蒲姆氏鬃(böhm′s bristles, BB) 蒲姆氏鬃分布于产卵鞘内侧边缘和基部(图2B、2D).长短不一,长度为2.23~13.22 mm,直径0.63~1.44 mm.不具基窝,通常垂直于体表,短而尖.
2.2.3 钟形感器(sensilla campaniformia, SCa) 钟形感器在产卵鞘基部和端部有分布(图2C、2D).按照形状、大小可以将其分为2种类型:钟形感器Ⅰ分布在产卵鞘基部,外圈圆形凹陷,边缘光滑,中间部分凸出;钟形感器Ⅱ分布在产卵器端部倒钩状组织处,呈圆形,中心部分稍稍凸起.

A.毛形感器(STⅠ为毛形感器Ⅰ,STⅡ为毛形感器Ⅱ);B.蒲姆氏鬃(BB);C.浅凹状感器(SD)和钟形感器Ⅱ(SCaⅡ);D.栓锥感器(SS)和钟形感器Ⅰ(SCaⅠ);E-F.腔锥感器(SC);G.刺形感器(SCh);H.分泌毛孔(SP).图2 产卵器和腹部的感器Fig.2 Sensilla on the ovipositor and abdomen
2.2.4 浅凹状感器(slight surface depression, SD) 该感器在产卵针端部排列分布(图2C),凹陷较浅,呈椭圆形,腔中央平坦无凸起.
2.2.5 栓锥感器(sensilla styloconica, SS) 仅在产卵器基部发现该感器,在凹陷中心,表皮凸起成栓状,类似拇指状,其顶端着生一个锥形感器,感器中部具有环纹(图2D).
2.2.6 腔锥感器(sensilla coeloconical, SC) 该感器在产卵针端部有较多分布(图2E、2F).乳头状腔锥感器的凸起较小,位于相对较大的凹陷中,凹陷与中心凸出形成腔穴,直径为0.82~1.12 mm.
2.2.7 刺形感器(sensilla chaetica, SCh) 刺形感器分布在产卵鞘端部,其外形刚直如刺(图2G),比毛形感器硬,长度比蒲姆氏鬃长,顶端较圆钝,直立于表面,不具基窝.长为20.12~52.56 mm,直径1.42~2.85 mm.
2.2.8 分泌毛孔(secretory pore, SP) 分泌毛孔在腹部、产卵鞘和产卵针上都有存在,呈网状分布(图2H),没有表皮腺.
3 讨论与结论
橘小实蝇主要以幼虫钻蛀取食为害和成虫产卵为害为主,其藏于果实组织内,十分隐蔽,寄生蜂的触角一般不易接触到寄主,因此,寄生蜂在识别和接受寄主的过程中,腹部和产卵器是寄生蜂与寄主最直接的接触和感受部位,对其能否成功寄生具有重要的作用[15].例如,有些寄生蜂不在已死的寄主上产卵,这可能是因为死亡的寄主体液不流动,或在寄生蜂产卵时死亡寄主体壁纵肌不收缩,产卵器感受不到寄主体液流动或挤压,从而不进行产卵行为[16];某些单寄生种类的寄生蜂,能够识别已经寄生和未被寄生的寄主,从而保证后代能够在未被寄生的寄主上存活[17],这种能力也是通过产卵器上的感器获得.但这些行为是哪些感器所决定的,目前尚未报道.
本研究中的4种潜蝇茧蜂的产卵器和腹部形态极为相似,但在色块、横纹和大小上有所差异.另外,阿里山潜蝇茧蜂和布氏潜蝇茧蜂的产卵针端部尖细,而长尾潜蝇茧蜂和切割潜蝇茧蜂的产卵针端部呈倒钩状,这也在小菜蛾(PlutellaxylostellaL.)寄生蜂产卵器[11]中被发现,其主要起到刺穿寄主表皮和固定产卵器的作用.陈佳[18]对这4种寄生蜂的种间竞争研究发现,切割潜蝇茧蜂和长尾潜蝇茧蜂都不能识别先前被阿里山潜蝇茧蜂寄生过的幼虫,而布氏潜蝇茧蜂却能识别先前被阿里山潜蝇茧蜂寄生过的幼虫,这种差异性可能与寄生蜂产卵器中的某些化学感器差异相关.
在4种潜蝇茧蜂的腹部和产卵器上,共发现8种感器,分别为毛形感器、蒲姆氏鬃、钟形感器、浅凹状感器、栓锥感器、腔锥感器、刺形感器和分泌毛孔.不同类型的感受器具有不同的功能.一般认为,蒲姆氏鬃和刺形感器是机械感器,当遇到机械刺激时,能够起到机械感触的功能.蒲姆氏鬃多分布于触角,在产卵鞘上出现的报道较少,而本研究中的4种寄生蜂的产卵鞘都发现了蒲姆氏鬃.毛形感器不仅具有感受机械刺激的功能,还是性信息素的主要感器,同时具有味觉功能,能探测湿度变化和吸附空气中的气味因子[19];此外,在寄生蜂进行产卵时,毛形感器和刺形感器最先接触产卵基质,因此它们在寄主选择中还具有定位和刺探位点的功能[20].毛形感器普遍被认为是昆虫触角上分布最广、数量最多的感器;本研究发现,毛形感器在4种茧蜂产卵器和腹部也分布最广、数量最多.此前已经有许多类似的发现,如在白蛾周氏啮小蜂(ChouioiacuneaYang)、甘蓝夜蛾赤眼蜂(TrichogrammabrassicaeBezdenko)和麦蛾柔茧蜂(HabrobraconhebetorSay)产卵鞘都发现许多毛形感器[19].钟形感器和腔锥感器是一种化学感器,有助于寄生蜂搜寻合适的产卵位点[21].有关钟形感器形态的描述有2种:(1)钟形感器无孔,表皮呈蘑菇状凸起,插入表皮圆柱状凹陷中,推测其可能是一种温湿度感器[22];(2)钟形感器有孔,从表皮伸出,该感受器能感受温湿度、二氧化碳和气味[23].4种橘小实蝇寄生蜂的钟形感器形态与前者的描述相符.分泌毛孔是典型的化学感受器,具有分泌功能[24].浅凹状感器在小菜蛾寄生蜂[11]的产卵器上也有分布,但关于其功能还未见报道.
本试验仅观察了各种感器的外部形态和分布,感器的类型及功能主要是根据其形态、分布及寄生蜂的一些生物学特性推测而来,存在一定的局限性.对于感器的内部构造和功能,还可利用单细胞电位仪和透射电镜进行深入研究.
[1] 梁光红,陈家骅,杨建全,等.桔小实蝇国内研究概况[J].华东昆虫学报,2003,12(2):90-98.
[2] 黄可辉.检获检疫性害虫──桔小实蝇[J].华东昆虫学报,1994,3(2):104-107.
[3] 张清源,林振基,刘金耀.橘小实蝇生物学特性[J].华东昆虫学报,1998,7(2):65-68.
[4] PAN Z, LI D, ZENG L. Effects of environmental factors on the pathogenicity ofBeauveriabassianatoBactroceradorsalis(Hendel)[J]. Journal of Environmental Entomology, 2008,30(1):13-17.
[5] WHITE I M, ELSON-HARRIS M M. Fruit flies of economic significance: their identification and bionomics[J]. Environmental Entomology, 1992,22(22):1 408.
[6] 陈景芸,蔡平,张国彪,等.橘小实蝇发生与综合防治研究进展[J].安徽农业科学,2011,39(28):17 324-17 326.
[7] BARCLAY H J, MCINNIS D, HENDRICHS J. Modeling the area-wide integration of male annihilation and the simultaneous release of methyl eugenol-exposedBactroceraspp. sterile males[J]. Annals of the Entomological Society of America, 2014,107(1):97-112.
[8] 郭义.植物挥发物及学习经历在丽蚜小蜂EncarsiaformosaGahan寄主寻找行为中的作用[D].重庆:西南大学,2007.
[9] 刘万学.植物挥发性信息化合物在棉铃虫齿唇姬蜂寄主寻找行为中的作用[D].北京:中国农业科学院,2002.
[10] 尹新明,高艳,王高平,等.寄生蜂感受器的形态与功能[J].河南农业大学学报,2003,37(2):129-133.
[11] 王世贵,蒋芸芸.两种小菜蛾寄生蜂产卵器感器的形态和超微结构[J].动物分类学报,2007,32(2):369-375.
[12] CAI P, GU X, YAO M, et al. The optimal age and radiation dose forBactroceradorsalis(Hendel) (Diptera: Tephritidae) eggs as hosts for mass-rearedFopiusarisanus(Sonan) (Hymenoptera: Braconidae)[J]. Biological Control, 2017,108:89-97.
[13] 毕康.阿里山潜蝇茧蜂触角感受器超微观察及其对寄主卵表化合物的反应[D].福州:福建农林大学,2012.
[14] 涂蓉.橘小实蝇及其幼虫寄生蜂的嗅觉行为研究[D].福州:福建农林大学,2012.
[15] 黄蓬英,徐梅,林玲玲,等.桉树枝瘿姬小蜂成虫感器超微结构[J].应用昆虫学报,2010,47(4):752-758.
[16] CHEAH C A, COAKER T H. Host finding and discrimination inDiglyphusisaea, a parasitoid of the chrysanthemum leaf miner,Chromatomyiasyngenesiae[J]. Biocontrol Science and Technology, 1992,2(2):109-118.
[17] VINSON S B, IWANTSCH G F. Host suitability for insect parasitoids[J]. Annual Review of Entomology, 1980,25(25):397-419.
[18] 陈佳.橘小实蝇几种寄生蜂种间竞争的研究[D].福州:福建农林大学,2010.
[19] 徐晓蕊,张旭臣,祈金玉,等.白蛾周氏啮小蜂产卵器感器的形态和超微结构[J].环境昆虫学报,2012,34(4):459-465.
[20] 章玉苹,范一霖,郑苑,等.橘小实蝇本地寄生蜂长尾全裂茧蜂雌成虫产卵器感器的扫描电镜观察[J].环境昆虫学报,2012,34(4):466-471.
[21] PETER W L. Identification of Danish adult females ofLype(Trichoptera; Psychomyiidae), with notes on reproduction[J]. Aquatic Insects, 1995,17(2):65-70.
[22] YOKOHARI F. The sensillum capitulum, an antennal hygro- and thermoreceptive sensillum of the cockroach,PeriplanetaamericanaL.[J]. Cell and Tissue Research, 1981,216(3):525-543.
[23] DIETZ A, HUMPHREYS W J. Scanning electron microscopic studies of antennal receptors of the worker honey bee, including sensilla campaniformia[J]. Annals of the Entomological Society of America, 1971,64(4):919-925.
[24] NENON J P, LANNIC J L E, KACEM N, et al. Micromorphologie de l′ovipositeur des hyménoptères et évolution des symphytes phytophages aux apocrites parasitoïdes[J]. Comptes Rendus De Lacadémie Des Sciences Série Sciences De La Vie, 1995,318:1 045-1 051.
Scanningelectronmicroscopyobservationsontheabdominalandovipositor′ssensillaof4parasitoidsofBactroceradorsalis
GU Xiaohong, CAI Pumo, YANG Yanchuan, WANG Cong, YANG Qingyang, JI Qing′e, CHEN Jiahua
(Institute of Beneficial Insects, College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China)
The morphology and ultrastructure of ovipositor and abdominal sensilla of four species of parasitoids, includingFopiusarisanus(Sonan),Psytalliaincise(Silvestri),Diachasmimorphalongicaudata(Ashmead) andFopiusvandenboschi(Fullaway), were observed and studied by scanning electron microscopy (SEM). Eight kinds of sensilla were found, including sensilla trichoidea, böhm′s bristles, sensilla campaniformia, slight surface depression, sensilla styloconica, sensilla coeloconical, sensilla chaetica and secretory pore. Besides, the ovipositor speculas ofF.arisanusandF.vandenboschiwere tapering, while the ovipositor speculas ofP.inciseandD.longicaudatawere barb-shaped.
parasitoid;Bactroceradorsalis; sensilla; scanning electron microscopy
2017-03-25
2017-06-05
福建农林大学科技发展基金项目(KFA17456A).
谷小红(1990-),女,硕士研究生.研究方向:农业昆虫与害虫防治.Email:409383559@qq.com.通讯作者陈家骅(1938-),男,教授,博士生导师.研究方向:茧蜂分类与害虫防治.Email:jhchen@163.com.
S436.5
A
1671-5470(2017)06-0601-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.06.001
(责任编辑:杨郁霞)
