祁连山北坡草甸草原地上生物量与土壤理化性质的关系
2017-12-23贾文雄陈京华
王 洁,贾文雄,赵 珍,陈京华,丁 丹
(西北师范大学 地理与环境科学学院,兰州730070)
祁连山北坡草甸草原地上生物量与土壤理化性质的关系
王 洁,贾文雄,赵 珍,陈京华,丁 丹
(西北师范大学 地理与环境科学学院,兰州730070)
为了揭示祁连山北坡草甸草原的地上生物量与土壤理化性质的关系,在祁连山北坡东、中、西部选择了三个典型样地,连续收集了两个生长季的野外采样数据,开展植物地上生物量与土壤理化性质的关系研究。结果表明:(1)2014年生长季每个样地的地上总生物量均大于2013年生长季的,且两个生长季的地上总生物量从东到西均依次增加;(2)土壤含水量、土壤温度、土壤有机质、有效钾、p H值在不同样地、不同生长季、不同土层差异性明显,而土壤碱解氮、有效磷相对稳定,差异性不明显;(3)地上生物量与土壤温度呈显著正相关,与土壤水分相关性不明显,与有效磷呈极显著正相关,与p H值呈显著正相关,对地上生物量影响较大的因子是地温、有效磷以及p H值。土壤养分主要通过影响土壤的理化性质来决定植被的生长,进而影响植物的生物量。
祁连山北坡;草甸草原;地上生物量;土壤理化性质
草地生态系统是全球分布最广泛的陆地生态系统之一,草原面积约占全球陆地面积的1/5[1]。土壤是陆地生态系统的重要组成部分,是环境与生物相互作用的产物,储藏着碳、氮、磷等大量的营养物质[2],也是陆地生态系统中物质与能量相互交换的重要场所。土壤养分对于植物的生长起着关键性的作用,会直接影响植物群落的组成与生理活动,决定生态系统的结构、功能以及生产力水平[2]。但是,不合理的开发利用使草地生态系统逐渐退化,会导致土壤性质发生变化。因此,对草地土壤理化性质的研究有助于全面认识土壤养分状况,对草地的维护和管理有着重要的意义。Li等研究了不同管理模式下牧草土壤碳库的变化受管理方式的影响[3-4];张俊华等研究了不同土地利用类型下的土壤有机碳时空分布及其与区域环境的关系[5];盛海彦研究了放牧干扰对高寒金露梅灌丛草甸群落和土壤因子的影响[6];孙旭生研究了库布齐沙漠不同沙地类型植物群落与土壤的关系[7];李恩宇等研究了青藏高原高寒草甸不同季节土壤理化性质及酶活性对施肥处理的响应[8]。这些研究侧重于土壤碳库与生态环境的关系、植物群落与土壤环境的关系以及土壤理化性质与施肥的关系,为全面认识土壤环境与生态系统的关系奠定了基础。
祁连山地处青藏高寒区、西北半干旱区和东部湿润区的交界带,对气候变化非常敏感[9]。目前,由于全球气候变化和人类活动影响日渐加剧,祁连山的生态系统正发生着剧烈的变化,不仅表现为植被的退化,还表现出土壤环境的变化。祁连山土壤环境的变化是对生态系统变化的一种响应,它不仅与土壤本身的性质有关,而且受到外界强烈的限制与干扰,并与地理位置、气候环境、地质结构和成土过程等因素密切相关。在祁连山开展了一些植被与土壤理化性质关系方面的研究,如Li等研究了祁连山土壤碳库变化与土壤颗粒性质的关系[10];高超研究了东祁连山不同退化程度高寒草甸草原土壤有机质及其对草地生产力的影响[11];杨成德等研究了东祁连山高寒草甸土壤微生物量及其与土壤物理因子相关性特征[12]等。以往研究主要侧重于土壤有机质及其对草地生产能力的影响,微生物量与土壤因子的关系等,而对生物量与土壤因子之间的关系尚未深入研究。开展土壤环境变化对生态系统变化的响应十分重要,本研究通过调查祁连山北坡草甸草原的土壤理化性质对地上生物量的影响,旨在探讨不同植物群落在大尺度空间上的变化,分析不同植物群落的地上生物量与土壤理化性质的关系,进而为合理利用土地资源及草地生态系统的恢复与重建奠定基础。
1 研究区概况
祁连山(95°—104°E,35.5°—40°N)位于青藏高原东北边缘,横跨甘肃、青海两省,全长约1 000 km,最宽处约300 km,东起乌鞘岭,西至当金山口,北临河西走廊,南接柴达木盆地,由多条西北—东南走向的平行山脉和宽谷组成,山势由西向东逐渐降低[13]。祁连山年均温0.6℃,年降水400~700 mm,降水主要集中在6—8月,属于典型的大陆性气候,同时又具有水热垂直地带性变化的山地气候特点[14],土壤以山地森林灰褐土、山地褐土、荒漠棕钙土为主,对应形成了草甸草原、典型草原和荒漠草原[14],其中草甸草原是祁连山分布最广和最主要的植被类型。
2 研究方法
2.1 地上生物量和土壤样品的采集
本文在祁连山北坡的青阳山岔(QS,101°55′59″E,37°43′31″N)、民乐卧马山(MS,100°39′25″E,38°24′18″N)、肃南鄂博台子(SS,99°35′40″E,38°47′10″N)三个典型的草甸草原样地开展研究,三个样地的海拔分别为2 772 m,2 711 m,2 756 m,坡度分别为62°,67°,56°,坡向均为半阳坡。青阳山岔与民乐卧马山之间的空间距离大约为135 km,民乐卧马山与肃南鄂博台子之间的空间距离大约为101 km。在2013年、2014年两个连续生长季的5—9月,在青阳山岔、民乐卧马山、肃南鄂博台子布设植被样方,具体方法是将20 m的卷尺拉成一条直线,在0~3 m,10~13 m,20~23 m处做一个3 m×3 m的样方,在样方四角和中心设置5个1 m×1 m的小样方作为重复,采用针刺法统计物种的盖度,用卷尺测量物种的高度(每个物种测量15次),用圆圈法统计物种的频度(每个样地抛掷30次),统计样方中出现的物种数,按不同物种贴地面剪取植物的地上部分,装入密封塑料袋中带回。在实验室,将采集的草样在恒温65℃的烘箱内连续烘干至恒重,称量干重。同时,在剪完植被后的样方上采用对角线法,用土钻分层采集土壤样品,分层测至30 cm,每层10 cm,重复3次,将3次重复分层混和为一个混合样,然后将土样用塑料袋密封,带回实验室后取出一部分用来测定其含水量;剩余土样风干后制成过0.15 mm和1 mm的分样,进一步进行土壤理化性质的测定。
2.2 土壤理化性质的测定及数据处理
土壤含水量用烘干法、土壤地温用地温计、p H值用酸度计法、有效N,P,K通过TFC1B型速测仪来比色测定、土壤有机质用重铬酸钾容量法测定。
2.3 统计分析
用Excel 2010对数据进行预处理,SPSS 16.0软件进行单因素方差分析和Pearson相关性分析。
3 结果与分析
3.1 祁连山北坡草甸草原的地上生物量变化
从图1中可以看出,2013年,样地QS呈先增加后减小的趋势,其值在8月达最大,为118.40 g/m2,样地QS的地上总生物量为357.87 g/m2;样地 MS与样地SS的变化趋势一致,均是先增加后减小再增加,且9月值均达最大,分别为158.84 g/m2,135.87 g/m2,两个样地的地上总生物量分别为364.88 g/m2,389.37 g/m2。2014年,样地 QS的变化趋势同2013年的,最大值出现在8月且为151.74 g/m2,样地QS的地上总生物量为397.28 g/m2;样地 MS的变化趋势不同于2013年的,2014年在整个生长季呈增加趋势,最大值出现于9月且为144.51 g/m2,其地上总生物量为398.65 g/m2;样地SS的变化趋势在5—7月同2013年的,均呈增加趋势,2013年7—9月呈“V”型,而2014年呈倒“V”型,且最大值出现于8月,其值为136.61 g/m2,该样地的地上总生物量为427.02 g/m2。
综上所述,在时间尺度上,祁连山草甸草原三个样地在2014年的地上总生物量均大于2013年的,在空间尺度上,从东到西,地上总生物量依次增加。不同样地两年的地上生物量在生长季前期和后期波动较小,在8月份波动较大。通过对比可知连续两个生长季三个样地间的差异不大(p>0.05)。

图1 祁连山北坡不同草甸草原群落的地上生物量
3.2 祁连山北坡草甸草原的土壤理化性质变化
3.2.1 土壤含水量的季节变化特征 从图2中可以看出,三个样地在2013年的变化趋势比较一致,均呈“W”型,在生长季前期和后期波动较小,中期波动较大,在7月份均达最大值,而样地QS在2014年的土壤含水量变化呈缓慢上升趋势,样地MS在2014年的土壤含水量变化在生长季(5—9月)期间变化不是很大,总体呈先增加后减小的趋势,样地SS在2014年的土壤含水量变化在生长季期间也不是很大,总体呈减小趋势。在时间上,样地QS,SS在2013年的土壤含水量总量大于2014年的;在空间上,从东到西(QS→MS→SS),两年的土壤含水量总量均依次减小。通过对比可知,2013年三个样地间的差异性不显著(p>0.05),而2014年样地QS,MS之间的差异性不显著(p>0.05),但样地QS,MS与样地SS之间的差异性均极其明显(p<0.01)。

图2 不同样地两年的0—30 cm土层土壤水分季节变化
3.2.2 土壤含水量及地温在土壤剖面上的变化 由表1可以看出,2014年,样地土壤含水量为10.32%~33.84%,样地QS的土壤含水量在不同深度差异不显著,样地MS的第一层与第三层差异性比较显著(p<0.05),样地SS的第一层与第二层、第三层的差异均极其显著(p<0.01),样地QS,MS与样地SS之间的差异极其显著(p<0.01)。2013年,样地土壤含水量为15.84%~34.31%,样地QS,MS,SS在不同深度之间差异分别不显著,在土壤剖面方向上从上到下基本遵循土壤水分含量依次减少的趋势,三个样地之间的差异并不明显。三个样地在土壤剖面方向上从上到下土壤含水量分别依次减少。综上所述,在时间上,样地QS,SS在2013年的土壤含水量总量大于2014年的;在空间上,从东到西(QS→MS→SS)两年的土壤含水量总量均依次减少。
2014年,样地土壤地温为10.36~16.00℃,三个样地之间的差异不明显,平均地温SS>QS>MS,不同深度的差异也不明显,在土壤剖面方向上分别呈0—10 cm>20—30 cm>10—20 cm的趋势。2013年,样地土壤地温为11.20~17.50℃,平均地温QS>MS>SS,在土壤部面方向上样地MS,SS分别呈0—10 cm>10—20 cm>20—30 cm,样地QS的地温在第一层与第二层之间的差异比较显著(p<0.05),样地MS的地温在第一层与第三层之间的差异比较显著(p<0.05),样地SS的地温在不同深度差异不明显。三个样地之间的差异也不显著,在空间上从东到西地温依次减小,三个样地的地温分别在土壤剖面方向上从上到下总体呈递减的趋势。

表1 不同样地两年的土壤含水量及地温在土壤剖面上的变化
3.2.3 有机质的变化 由表2可知,2014年,样地有机质为35.31~93.02 g/kg,三个样地的有机质含量在不同深度差异很显著,在样地QS中,第一层与第二层、第三层的差异性极其显著(p<0.01),在MS样地中,第一层与第二层、第三层差异显著(p<0.05),在样地SS中,第一层与第二层呈显著性差异(p<0.05),与第三层呈极显著性差异(p<0.01),三个样地的有机质含量主要集中于第一层,并且从上到下依次递减。三个样地之间的差异性也很明显,样地SS与样地MS,QS呈极显著差异(p<0.01),样地 MS与样地QS呈显著差异(p<0.05)。2013年,样地有机质为37.25~93.87 g/kg,三个样地的有机质含量在不同深度差异是显著的,在样地QS中,第一层与第二层、第三层的差异性极其显著(p<0.01),在MS样地中,第一层与第三层差异显著(p<0.05),在样地SS中,第一层与第二、第三层差异显著(p<0.05),三个样地的有机质含量同2013年的主要集中于第一层,沿土壤剖面方向从上到下分别依次递减。三个样地之间的差异性也是非常显著的,样地SS与样地MS,QS呈极显著差异(p<0.01),样地MS与样地QS呈显著差异(p<0.05)。综上所述,三个样地两年的变化规律基本一致,在时间上,2013年的有机质含量在相对应的样地及相对应的深度其值均大于2014年的;在空间上,从东到西(QS→MS→SS)三个样地两年的有机质含量分别是递减的。
3.2.4 碱解氮、有效磷、有效钾的变化 碱解氮、有效磷在两年相对应的样地及相对深度之间的差异均不明显。2014年,样地碱解氮、有效磷、有效钾分别为8.47~30.95,44.40~79.60,34.50~216.30 mg/kg,碱解氮的平均值QS>MS>SS,有效磷的平均值SS>MS>QS,且有效磷主要集中于第三层。有效钾在样地QS中,第一层与第三层呈显著性差异(p<0.05),样地MS,SS在不同深度差异均不明显。三个样地之间的差异也比较明显,样地SS与样地MS呈极显著差异(p<0.01),与样地 QS呈显著差异(p<0.05)。三个样地两年的平均值均是SS>QS>MS,且相对应的值变化不大。2013年,样地碱解氮、有效磷、有效钾分别为12.58~56.08,38.15~210.80,21.80~206.60 mg/kg,碱解氮的平均值QS>SS>MS,有效磷的平均值QS>MS>SS,且有效磷主要集中于第二层。有效钾在样地QS中,第一层与第三层之间的差异性较明显(p<0.05),在样地 MS中,第一层与第二、第三层均呈极显著差异(p<0.01),在样地SS中,不同深度差异不明显。三个样地之间的差异并不明显。2013年,有效钾主要集中于第一层,2014年不太明显。
3.2.5 p H值的变化 2014年,样地p H值为8.10~8.54,样地QS的p H值在第一层与第三层之间的差异较显著,样地MS,SS在不同深度差异不明显。2013年,样地土壤p H值为8.04~8.54,在样地QS中,其土壤的p H值在不同深度差异不明显,在样地MS,SS中其p H值在第一层与第三层之间的差异性较显著(p<0.05)。三个样地之间的差异性较明显,样地SS与样地QS呈极显著差异(p<0.01),与样地 MS呈显著差异(p<0.05)。三个样地两年的p H值均偏碱性且分别在土壤剖面方向上从上到下呈增加趋势。
3.3 祁连山北坡草甸草原土壤理化性质对地上生物量的影响
由表3可以看出,2014年,在样地QS中碱解氮与p H值呈极显著正相关(p<0.01),有效钾与有机质呈显著负相关(p<0.05)。在MS样地中地上生物量与p H值呈显著正相关(p<0.05),有效磷与地温呈显著负相关(p<0.05)。在SS样地中有效钾与地温呈显著负相关(p<0.05)。2013年,在样地QS中地上生物量与地温呈显著正相关(p<0.05),碱解氮与p H值呈显著正相关(p<0.05)。在MS样地中地上生物量与有效磷呈极显著正相关(p<0.01),碱解氮与有机质呈显著负相关(p<0.05)。在SS样地中有效磷与有机质呈极显著正相关(p<0.01),与地温呈显著负相关(p<0.05),有机质与地温呈显著负相关(p<0.05)。综上可见,对地上生物量影响较大的因子分别是地温、有效磷以及p H值。
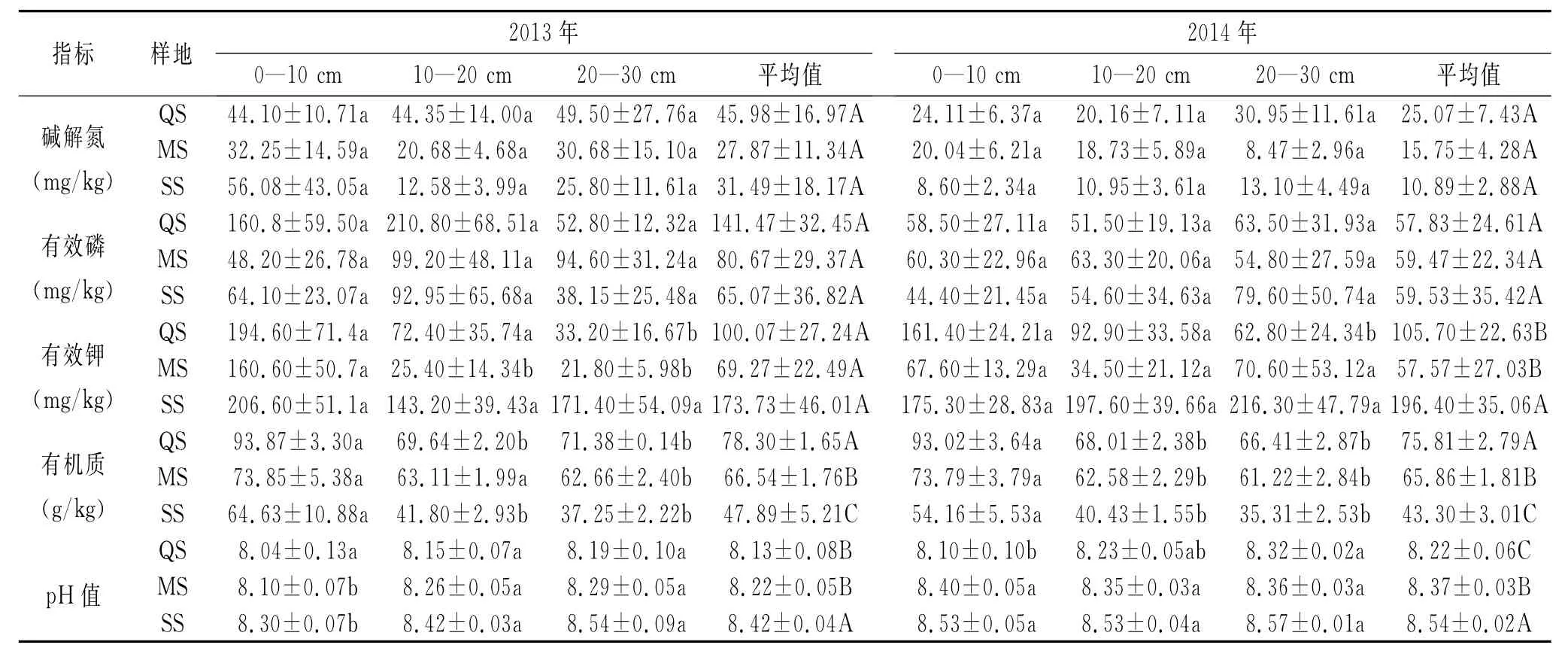
表2 土壤理化性质对不同样地地上生物量的影响

表3 土壤理化性质与不同样地地上生物量相关性分析
4 讨论与结论
(1)从图1中可以看出,两个连续生长季的地上生物量季节变化明显。最大值一般出现在8月或9月,最小值均出现在5月,这是由于祁连山北坡草地受低温的影响,一般在5月初各草地植被才陆续返青,地上生物量才开始积累,随着气温的回升和降水的增多,草本植被陆续生长,生物量也逐步增大[15],又由于降水在7月、8月比较多并且具有滞后性,再加上牧区放牧的影响,可能是导致地上生物量在8月或9月份下降的主要原因。从图中可以看到,6月、7月植被地上生物量的月增长量较大,导致地上生物量增加[16]。从图1中可知样地QS的增长趋势符合单峰模型,8月的增长速度有所减缓,9月气温降低,部分植物枯萎,导致地上生物量开始减小,而样地MS,SS第一个生长季的8月地上生物量均突然减小,其主要原因很可能是环境因素和放牧活动干扰引起的。9月份的生物量没有减少反而增加的原因主要是适宜冷湿环境生长的莎草科植物的继续生长[16]。不同样地在生长季的前期和末期连续两个生长季的波动均不是很大。
从时间尺度上看,2014年祁连山草甸草原的地上总生物量大于2013年的,说明草原植被生长状况较好。在空间尺度上,从东到西,草甸草原地上总生物量依次增多,其主要原因是东部受人为干扰强烈,草地出现不同程度的退化,植被结构渐趋简单,生态功能开始衰退[17],进而导致地上生物量减少。2013年,东部草甸草原样地的地上生物量最大值出现于8月,而西部、中部两个草甸草原样地的地上生物量最大值均推迟了一个月,这是因为中西部地区纬度较高,气温较低,植物萌发较晚,地上生物量积累过程较为缓慢[16]的缘故。
(2)土壤含水量的季节变化受到降水、气温、植被蒸腾、土壤蒸发及土壤结构、地形等多种因素的影响[18]。因此,土壤水分的变化规律十分复杂,是多种环境因子共同作用的结果。对祁连山北坡三个不同样地连续两个生长季土壤含水量的季节变化加以对比,可知变化是相对明显的。2014年,样地QS的土壤含水量呈递增趋势,样地 MS,SS的变化趋势基本一致。2013年,三个样地的土壤含水量变化趋势基本一致。两年土壤含水量在生长季的前期和末期变化均不明显,但2013年三个样地的中期波动较大,而2014年波动不是很大,这可能与草地在不同时期生长的降水量、降水强度以及草地生长状况等因子有关[15]。在生长季前期和后期,降水量较小,受气温和水文等的影响,草类生长比较缓慢,使得草地蒸发较小,土壤含水量波动较小;在生长季中期,由于降水较多,草类生长较快,草地蒸发较大,导致土壤含水量波动较大[15]。另外,空间上连续两个生长季的土壤含水量从东到西(QS→MS→SS)均呈递减趋势,原因很可能是东部地区靠近海洋,降水较多且植被生长状况较好,对降水有一定的拦截和减少地表径流的作用有关。
土壤含水量在土壤剖面方向上的分布依旧受降水、蒸发以及植被等环境因子的影响[18]。对不同样地不同土层深度土壤含水量进行分析,2013年三个样地在不同深度及样地间的差异均不明显,而2014年的差异较明显,说明土壤含水量在2014年的影响较大,其原因很可能是由降水量、降水强度、草地的蒸散量等因素引起的。两年的土壤含水量均主要集中于表层,表层以下相对稳定,且随土层深度的增加土壤含水量呈减小的趋势。2013年样地QS,SS的土壤含水量分别大于2014年的,很可能是因为在一定时期降水较多,草地蒸散量较少,导致水分供给较多,消耗少,使得土壤含水量高于2014年的。
2013年三个样地的地温在不同深度的差异较显著,在样地间的差异并不明显。2014年三个样地的地温在不同深度及样地间的差异均不明显。两年的平均地温均是样地SS>QS>MS,且2013年的地温在不同样地相应深度的温度普遍大于2014年的,其原因很可能是2013年整体的水热状况组合好于2014年的,另外样地SS处于西部,植被相对稀疏,地表接受太阳辐射较多,导致地温较高。
(3)土壤有机质是土壤重要的养分指标,同时还是形成土壤结构的重要因素,直接影响着土壤的肥力、持水能力、土壤抗侵蚀能力和土壤温度等,是表明土壤特性的重要指标[15]。本文通过对三个不同样地两年有机质的差异比较,三个样地在不同深度及样地间的差异是非常明显的。三个样地在土壤剖面方向上从上到下有机质含量基本呈递减趋势,说明土壤有机质主要富集于土壤表层且主要来源于植物的地上凋落物和地下根系,这也是表层有机质含量与其他层有机质含量差异显著的原因[11]。不同样地连续两个生长季的有机质总量差异不是很大,但2013年的总量略大于2014年的,由于有机质的总量取决于生物量的生产与分解的平衡状态,以及土壤储存有机质的能力[19],说明2013年归还于土壤的有机质较多而分解的较少。空间上从东到西均呈递减趋势,说明东部地区土壤肥沃,越往西土地越贫瘠。碱解氮、有效磷在连续两个生长季不同样地及不同深度之间的差异不是很明显,而有效钾的差异是显著的。三个样地在2013年的碱解氮、有效磷的总含量均大于2014年的,且从东到西其量均依次减小,原因很可能是2013年的植被较稀疏,碱解氮、有效磷消耗较少,因此,残留于土壤中的量相对较多。两年的有效钾均是样地SS>QS>MS,原因很可能是样地SS与QS受到人类活动(放牧)干扰较大,这是因为放牧可使土壤有效钾显著增加[6]。p H值的差异是明显的,在土壤剖面方向上从上到下均呈递增趋势,空间上从东到西均呈递增趋势,原因很可能是由于西部地区植被覆盖低,有许多次生的裸地面积大,地表蒸发量大,导致p H值增大,还有可能是由于鼠类活动及放牧干扰等因素,加速了土壤养分的矿化,这与陈涛、盛海彦等人的研究结果一致[6,19]。
(4)通过相关性对比可知,土壤的理化性质对不同样地不同生长季地上生物量的影响不同且土壤理化性质之间的相关性是明显的。土壤含水量与地上生物量在本文中呈正相关、不相关、负相关三种结果,但均不显著,原因可能是含水量较低小于植被生长所需,含水量合适满足植被生长以及含水量较高限制植被生长造成的[20]。地上生物量与地温呈显著正相关、相关性不明显两种结果,由于在一定条件下地温越高,有机质的矿化速率加快,越有利于植物的生长[21]。地上生物量与土壤有机质、碱解氮、有效钾的相关性均不明显,而与有效磷呈极显著正相关、不相关两种结果,前者与高超、石红霄等人的研究结果不一致[22-23],后者与石红霄等人的研究结果一致[23]。地上生物量与p H值呈显著正相关、不相关两种结果,说明影响地上生物量的因子较多,这可能与研究地点的气候、植物群落、草地健康状况及土壤p H值的变化幅度等有关[12]。
土壤理化性质之间的相关性是相当明显的,有机质与碱解氮呈显著负相关,与有效磷呈极显著正相关,与有效钾呈极显著负相关,与地温呈显著负相关,与土壤含水量相关性不明显。碱解氮、有效磷、有效钾之间的相关性不大,有效磷、有效钾分别与地温呈显著负相关。碱解氮与p H值呈极显著正相关,而有效磷、有效钾与p H值的相关性不明显。可以看到土壤含水量对地上生物量及土壤的理化性质影响均不显著。土壤的物理性质对土壤的化学性质有影响,但略有不同。相关研究表明,土壤水分过多会影响有机质的分解[24],一般情况下没有温度的影响大,本文的研究结果与高超等人的研究结果一致[11]。温度的变化是影响土壤有机质组成的主要外部条件[25]。王淑平等人的研究表明温度对有机碳的影响较复杂,适宜的温度有利于土壤有机碳的积累,否则对有机碳的积累具有负效应[26]。也有人认为水分对土壤有机质的影响大于温度的[27]。一定条件下土壤温度的升高有利于碱解氮、有效磷、有效钾的生成。土壤温度升高,加快了有机质的矿化速率,以及磷矿化、钾矿化的速率,使得有机质及腐殖质含量逐渐减少,碱解氮、有效磷、有效钾的含量增多[6,11,19]。
[1]Scurlock J M O,Hall D O.The global carbon sink:a grassland perspective[J].Global Change Biology,1998,4(2):229-233.
[2]索南措.青藏高原东缘高寒草甸植被特征与土壤性质随海拔变化的响应研究[D].兰州:西北师范大学,2013.
[3]Li Xiaogang,Zhang Meilan,Li Zhuoting,et al.Dynamic s of soil properties and organic carbon pool in topsoil of zokor-made mounds at an alpine site of the Qinghai-Tibetan Plateau[J].Biol.Fertil.Soils,2009,45(8):865-872.
[4]Li Xiaogang,Li Fengmin,Bhupinderpal-Singh,et al.Soil management changes organic carbon pools in alpine pastureland soils[J].Soil and Tillage Research,2007,93(1):186-196.
[5]张俊华,李国栋,南忠仁,等.黑河中游不同土地利用类型下土壤碳储量及其空间变化[J].地理科学,2011,31(8):982-988.
[6]盛海彦.放牧干扰对高寒金露梅灌丛草甸群落和土壤因子影响的研究[D].陕西杨凌:西北农林科技大学,2008.
[7]孙旭生.库布齐沙漠不同沙地类型植物群落与土壤关系的研究[D].呼和浩特:内蒙古师范大学,2010.
[8]李恩宇,何贵永.青藏高原高寒草甸不同季节土壤理化性质及酶活性对施肥处理的响应[J].广西植物,2014,34(4):467-472.
[9]王合领.祁连山东缘高寒草甸区土地利用对土壤有机碳的影响研究:以甘肃连城国家级保护区为例[D].兰州:兰州大学,2014.
[10]Li X G,Li F M,Zed R,et al.Soil physical properties and their relations to organic carbon pools as affected by land use in an alpine pastureland[J].Geoderma,2007,139(1):98-105.
[11]高超.东祁连山不同退化程度高寒草甸草原土壤有机质特性及其对草地生产力的影响[D].兰州:甘肃农业大学,2007.
[12]杨成德.东祁连山高寒草甸土壤微生物量及其与土壤物理因子相关性特征[J].草业学报,2007,16(4):62-68.
[13]张禹舜,贾文雄,赵一飞,等.基于CASA模型研究祁连山地区植被净初级生产力的时空变化[J].西北植物学报,2014,34(10):2085-2091.
[14]侯兆疆,赵成章,李钰,等.不同坡向高寒退化草地狼毒株高和枝条数的权衡关系[J].植物生态学报,2014,38(3):281-288.
[15]常博.祁连山排露沟流域草地植被特征及其对水分条件的响应[D].兰州:甘肃农业大学,2014.
[16]贾文雄,刘亚荣,张禹舜,等.祁连山草甸草原物种多样性和生物量与气候要素的关系[J].干旱区研究,2015,32(6):1167-1172.
[17]马金宝,张培栋,李新荣.旱泉沟流域天然鹤丛退化成因及保育对策[J].草业科学,2007,24(4):23-26.
[18]张千千.六盘山叠叠沟小流域土壤水分动态变化与植被生长的研究[D].河北保定:河北农业大学,2009.
[19]陈涛.呼伦贝尔退化羊草草甸物种多样性及土壤理化性质的研究[D].兰州:甘肃农业大学,2008.
[20]杨成德,龙瑞军,陈秀蓉,等.东祁连山高寒草甸土壤微生物量及其与土壤物理因子相关性特征[J].草业学报,2007,16(4):62-68.
[21]李绍良,康师安.内蒙古草原土壤退化进程及其评价指标的研究[J].土壤通报,1997,28(6):241-243.
[22]高超.东祁连山不同退化程度高寒草甸草原土壤有机质特性及其对草地生产力的影响[D].兰州:甘肃农业大学,2007.
[23]石红霄,侯向阳,师尚礼,等.高山嵩草草甸初级生产力、多样性与土壤因子的关系[J].草业学报,2015,24(10):40-47.
[24]Pal D,Broadbent F E.Influence of moisture on rice straw decomposition in soils[J].Soil Science Society of America Journal,1975,39(1):59-63.
[25]徐跃,姚天全.气候因素对土壤有机质组成和性质的影响[J].山地研究,1994,12(3):163-168.
[26]王淑平,周广胜,吕育财,等.中国东北样带(NECT)土壤碳、氮、磷的梯度分布及其与气候因子的关系[J].植物生态学报,2002,26(5):513-517.
[27]王长庭,龙瑞军,王启基,等.高寒草甸不同海拔梯度土壤有机质氮磷的分布和生产力变化及其与环境因子的关系[J].草业学报,2005,14(4):15-20.
Study on the Relations Between the Above Ground Biomass and the Soil Physicochemical Properties in the North Slope of Qilian Mountains
WANG Jie,JIA Wenxiong,ZHAO Zhen,CHEN Jinghua,DING Dan
(College of Geography and Environment Science,Northwest Normal University,Lanzhou730070,China)
In order to reveal the relations between the above ground biomass of meadow steppe and the soil physicochemical properties in the north slope of Qilian Mountain.We chose three typical samples in the eastern,central and western parts in the north slope of Qilian Mountains,collected the sampling data in two consecutive growing seasons,analyzed and examined the above ground biomass and soil physiochemical properties.The results show that total above ground biomass of the each sample is greater in 2014 than in 2013,and the mass increases from the east to the west.The soil moisture,soil temperature,soil organic matter,available potassium,p H value are significantly different in different growing seasons and the different layers among the sample sites.However,soil alkali-hydrolysable nitrogen and available phosphorus are relatively stable,and the differences are not signigicant.Correlation analysis shows that total above ground biomass is significantly positively correlated with soil temperature,p H value,and is significantly positively correlated with the available phosphorus,but is not significantly correlated with soil moisture.The results reveal that the major influence factors on total above ground biomass are soil temperature,available phosphorus and p H value.The soil nutrients mainly depend on the physicochemical properties of the soil,which determines the growth of vegetation and affect the biomass of plants.
Qilian Mountain;meadow steppe;above ground biomass;soil physicochemical properties
S812;S153.6
A
1005-3409(2017)01-0036-07
2016-05-16
2016-06-06
国家自然科学基金地区科学基金项目(41161017)
王洁(1990—),女,甘肃秦安人,硕士,主要从事全球气候变化与生态环境演变研究。E-mail:m13893374364-1@163.com
贾文雄(1974—),男,博士,副教授,主要从事全球变化与生态水文研究。E-mail:wxjiaxy@163.com
