花的形态与花发育的ABCDE模型
2017-12-19孟雨婷黄晓晨侯元同邱念伟
孟雨婷, 黄晓晨, 侯元同, 邱念伟
(曲阜师范大学 生命科学学院, 曲阜 273165)
花的形态与花发育的ABCDE模型
孟雨婷, 黄晓晨, 侯元同, 邱念伟
(曲阜师范大学 生命科学学院, 曲阜 273165)
系统介绍了花发育的ABCDE模型的内容,解释了自然界中各类花形态形成的分子生物学机制,绘制了各种花发育的模式图,推理了各组基因的功能和突变体类型,有助于系统了解花发育的奥秘。
花结构;花发育;ABCDE模型;四聚体模型;异位表达
植物的花千姿百态,大部分的两性花具有花萼、花冠(花瓣的总称)、雄蕊群、雌蕊群(由心皮联合而成)、胚珠等五轮完整的结构(如桃花和梨花)。但有的花只有花萼和花瓣(如雪球荚蒾),有的花则没有花瓣(如桑树和麻栎),还有的花只有雌蕊或雄蕊(如杜仲和垂柳)。1991年美国科学家Coen和Meyerowitz首次提出花器官发育的“ABC模型”[1],为解释各类花发育现象提供了框架。随着研究的深入和更多的花发育基因被发现,出现了很多该模型无法解释的现象,20多年来科学家不断补充和修正“ABC模型”,形成了更为完善的“ABCDE模型”[2-5],能够解释大部分双子叶植物的花发育现象。单子叶植物中的同源基因表达结果证明,ABCDE模型在单子叶禾本科植物中同样适用,其中外稃相当于花萼,浆片相当于花瓣;不过在非禾本科单子叶植物中,ABCDE模型又有少许改动[6]。有文献仅从相关基因的角度介绍了花发育模型,内容并不系统,不利于读者全面了解ABCDE模型[5-7]。本文根据国内外研究进展,将ABCDE模型的内容进行了系统地汇总和整理,重新绘制了各类相关模型和突变表型图片,为相关教学和科研提供参考。
1 典型两性花的结构和花发育相关基因的分类
科学家发现植物花的发育有类似动物的同源异型现象,即同一分生组织的系列产物中,一类成员可以转变成形态或性质不同的另一类成员。目前克隆出了多个花发育相关基因,这些基因决定着花器官特征而不改变花器官的发端,所以也称为“同源异型基因”。根据这些基因在花器官形成中的作用分成A、B、C、D和E共5类。被子植物的典型两性花由外到内可以分为五轮结构:第1轮为花萼,第2轮为花冠(花瓣),第3轮为雄蕊群,第4轮为雌蕊群(心皮),第5轮为胚珠。五轮花结构的形成是5类基因协同作用的结果(表1),这5类基因编码的不是蛋白酶,而是决定花发育的转录因子,它们组合形成的不同蛋白四聚体,结合在DNA上,影响下游各类花器官特征基因的表达,从而决定每轮花器官形成[2-8]。如2A+2E结合形成的四聚体决定花萼的发育,A+2B+E决定花瓣,2B+C+E决定雄蕊,2C+2E决定心皮,C+D+2E决定胚珠(图1)。在上述基因的严格控制下,花的分生组织发育成各轮花器官,任何一组基因发生突变,花的形态都会发生异常。

表1 拟南芥中ABCDE同源基因的分类[2-8]
2 ABCDE模型的内容要点
A组基因控制第1、2轮花器官的发育;B组基因控制第2、3轮花器官的发育(在少数植物中控制第1~3轮的发育);C组基因控制第3、4、5轮花器官的发育(有的植物中只控制第3、4轮的发育);D组基因控制第5轮花器官的发育;E组基因则参与了全部5轮花器官的发育,其中E1-4之一参与第1轮花器官的形成,E3对第2、3、4、5轮花器官的发育均起决定作用[3,9]。也就是说,花萼由A组和E组基因控制,花瓣是由A组、B组、E组基因共同控制,雄蕊是由B组、C组、E组基因共同控制,心皮由C组和E组基因共同控制,胚珠则由C组、D组、E组基因共同控制(图2)。控制的方式就是各组转录因子形成不同的蛋白四聚体(图1)。
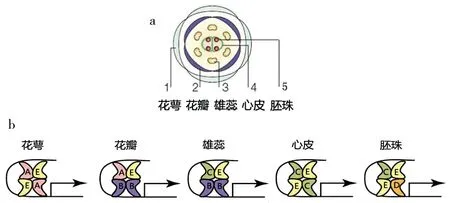
图1 花的五轮结构(a)和花发育转录因子的四聚体模型(b)
其中A组基因和C组基因相互抑制,若A组基因发生突变或丧失功能,C组基因的功能即扩大到整个花的分生组织,或者说C组转录因子会在花的第1、2轮发育中发挥作用。相反,若C组基因发生突变而丧失功能,A组基因的功能也会扩大到整个花的分生组织(图2)[2,4]。

图2 花器官决定的ABCDE模型(根据文献[2-10]修正)
3 各组基因单突变表型
以拟南芥、金鱼草等模式植物为材料,科学家们系统分析了各类同源异型基因的突变表型。
若A组基因发生突变,C组基因就会代替A组基因的功能,决定花萼形成的四聚体2A+2E就会变成决定心皮的四聚体2C+2E,导致花萼变成心皮(图1~3);同理A+2B+E四聚体变成C+2B+E,导致花瓣变雄蕊。花的五轮结构变成雌蕊、雄蕊、雄蕊、雌蕊、胚珠,没有花萼和花瓣(图3、4)。
若B组基因发生突变,花将变成花萼、花萼、雌蕊、雌蕊、胚珠。没有花瓣和雄蕊(图3、4)。
若C组基因发生突变,花将变成花萼、花瓣、花瓣、花萼、花萼。奇怪的是有些植物的花的第五轮——心皮和胚珠部位突变成了一朵只有花瓣的小花,产生“花中生花”现象(图3、4),这一现象在某些重瓣的樱花中也可以观察到,但其分子机制还没有确切解释,有可能是B类基因在心皮中异位表达所致。
若D组的3个基因STK、SHP1和SHP2分别或同时发生突变,都会使胚珠和种子发育受阻,花将变成花萼、花瓣、雄蕊、雌蕊,部分胚珠变成叶状或心皮状结构(图3、4)。
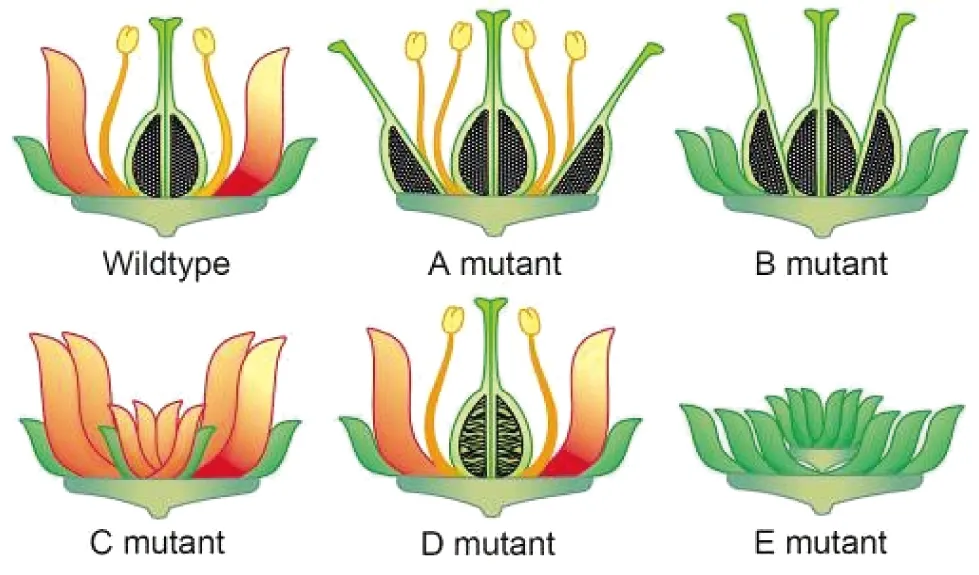
图3 5类基因单突变体的表型(本图E突变表型是sep1 sep2 sep3三突变体)[2]
Wildtype:野生型;mutant:突变体
若E组基因全部发生突变(sep1、sep2、sep3、sep4四重突变),花的五轮结构均不能形成,都变成了叶状结构(图4),与缺少ABC三基因突变体类似,说明E组基因参与了全部5轮花结构的发育。若仅SEP4发生突变,则无明显的突变表型,花结构类似野生型。sep1、sep2和sep4三重突变对花结构的影响也不大。而如果E1-3同时发生突变(sep1、sep2和sep3三重突变),花的五轮结构则全部变为萼片,心皮和胚珠消失,与BC双突变体表型类似,但BC基因的表达水平并无变化,其与BC双突变体的区别是心皮位置变成了类似一朵花的3轮萼片结构(图3 E突变中间部分),说明SEP1-3基因参与了心皮和胚珠的发育。在sep1、sep2纯合突变而sep3杂合突变的植物中,花瓣正常,但雄蕊减少。但在sep1、sep2和sep4纯合突变而sep3杂合突变的花中,花萼和花瓣异常,雄蕊畸形;剥开其心皮既能看到发育的胚珠,还能看到其内仍包含着心皮和雄蕊[9]。

图4 花发育的ABCDE突变模式与突变体花程式
上述结果说明E组的4个基因对各轮花器官的影响不是等比例的,其中SEP3基因对第2~5轮花结构的影响最大;SEP4在五轮花结构中都能表达,在花萼、花瓣、雄蕊、雌蕊的发育中都具有重要作用,但不是必需的。若SEP4与E组基因其他成员或其他组基因同时发生突变会加重突变表型[9-10]。E组基因的4个成员中表达其中任一个成员都能形成花萼;但若全部突变,花萼就会变成叶片状。由于决定第2~5轮花结构形成的E组基因是SEP3,所以在sep1、sep2和sep3三重突变中,参与第2~5轮花结构发育的蛋白四聚体均不能形成,C组基因不能发挥正常功能,只有参与花萼发育的2A+2E四聚体可以正常形成。因此2A+2E四聚体的活性会扩展至第2~5轮花器官[3,8-9],sep1、sep2和sep3三重突变的五轮结构因此全部变成了花萼。有部分文献认为E组基因未参与花萼的形成,主要是由于对E组基因成员的功能未完全搞清楚或者E组基因在不同的植物中功能不同所致[2,4]。
ABCDE模型也证明花是变态的枝条,花器官则是变态的叶片。例如E1-4基因全部缺失,花会变态形成叶状结构;反之,A-B-C-D-E联合作用也可以使营养器官转化为花结构。异位表达A-B-E类基因,能够使叶原基发育成花瓣;异位共表达B-C-E类基因,能够使叶原基转化为雄蕊;异位共表达A-B-C-D-E类全部基因,则可使叶原基转化成完整花器官[3,7]。异位表达时,缺少E组基因不能使叶原基发育成花器官,也证明E组基因是五轮花器官的发育所必需的。以上现象证明ABCDE基因联合作用决定了花器官的形成。很多植物的“老茎生花”现象就是异位表达了花发育基因,如紫荆、可可、阳桃;“叶上生花”现象也是如此,如青荚叶。因此,了解ABCDE模型对植物形态研究是很有帮助的。
4 ABCDE模型仍需不断完善
与ABC模型相比,ABCDE模型能够更好地解释各种异常的花结构,但仍然存在很多问题。如:不同植物中ABCDE模型不完全一致,有些植物中B组基因也参与了花萼的发育[11]。单子叶非禾本科植物中(如郁金香)B组基因不仅在第2和第3轮花器官中表达,也在第1轮花器官中表达,使第1轮和第2轮花器官均发育成了完全相同的花瓣结构[12-14]。还有些植物中C组基因未参与胚珠的形成[4]。ABCDE模型给我们提供了一个理论框架,在具体的每种植物中都可能需要适当修正,在解释不同植物花发育现象时应灵活应用ABCDE模型。
同时,更多的花发育相关基因也不断被鉴定出来,花发育的机制比ABCDE模型更加复杂,如新发现的CRC和SPT基因具有类似C组基因的功能,也能促进心皮的发育;CAL基因则能替代A组和E组基因,维持花瓣的发育[15-16]。另外,酵母三杂交实验证明各组基因成员形成蛋白四聚体类型比图1中的四聚体类型多得多,很多四聚体类型在花发育中的作用还不清楚;而且每种四聚体也并非只在某轮花分生组织中起作用,其活性可以扩展到其他轮的分生组织[16]。因此,ABCDE模型还不能解释自然界中所有的花发育类型,其内容仍需进一步丰富和不断修正。
[1]COEN E S,MEYEREWIM E M.The war of the whorls:genetic interactions controlling flower development[J]. Nature, 1991, 353(6339): 31-37.
[2]FERRARIO S, IMMINK R G,ANGENENT G C. Conservation and diversity in flower land [J]. Current Opinion in Plant Biology, 2004, 7(1): 84-91.
[3]KRIZEK B A, FLETCHER J C. Molecular mechanisms of flower development: an armchair guide [J]. Nature Reviews Genetics, 2005, 6(9): 688-698.
[4]THEISSEN G. Development of floral organ identity: stories from the MADS house [J]. Current Opinion in Plant Biology, 2001, 4(1): 75-85.
[5]李贵生, 孟 征, 孔宏智, 等. ABC模型与花进化研究[J]. 科学通报, 2003, 48(23): 2415-2421.
[6]王力先, 王永飞. 被子植物花发育的ABC模型研究进展[J]. 安徽农学通报, 2007, 13(8): 49-51.
[7]丛 楠, 程治军, 万建民. 控制花器官发育的ABCDE模型[J]. 中国农学通报, 2007, 23(7): 124-128.
[8]CAUSIERA B, SCHWARZ-SOMMERB Z, DAVIESA B. Floral organ identity: 20 years of ABCs [J]. Seminars in Cell & Developmental Biology, 2010, 21(1):73-79.
[9]DITTA G, PINYOPICH A, ROBLES P, et al. The SEP4 gene ofArabidopsisthalianafunctions in floral organ and meristem identity [J]. Current Biology(CB), 2004, 14(21): 1935-1940.
[10]尹伟平, 刘 闯, 潘美昕, 等. 植物生殖发育分子机理简述[J]. 生物学杂志, 2016, 33(4): 78-82.
[11]KIM S, KOH J, YOO M J, et al. Expession of floral MADS-box genes in basal angiosperms: implications for the evolution of floral regulators [J]. The Plant Journal, 2005, 43(5): 724-744.
[12]VAN TUNEN A J, EIKELBOOM W, ANGENENT G C. Floral organogenesis inTulipa[J]. Flowering Newsletter, 1993(16): 33-35, 37-38.
[13]KANNO A, SAEKI H, KAMEYA T, et al. Heterotopic expression of class B floral homeotic genes supports a modified ABC model for tulip(Tulipgesneriana) [J]. Plant Molecular Biology, 2003, 52(4): 831-841.
[14]李洪有, 王 婵, 李丽林, 等. 单子叶植物花器官发育的分子机制及修正的ABC模型[J]. 中国细胞生物学学报, 2013, 35(4): 526-535.
[15]刘建武, 孙成华, 刘 宁. 花器官决定的ABC模型和四因子模型[J]. 植物学通报, 2004, 21(3): 346-351.
[16]FAVARO R, PINYOPICH A, BATTAGLIA R, et al. MADS-box protein complexes control carpel and ovule development in Arabidopsis [J]. Plant Cell, 2003, 15(11): 2603-2611.
ThefloralmorphologyandtheABCDEmodeloffloralorgandevelopment
MENG Yu-ting, HUANG Xiao-chen, HOU Yuan-tong, QIU Nian-wei
(College of Life Science, Qufu Normal University, Qufu 273165, China)
In this paper, the ABCDE model of floral organ development was systematically introduced, and the molecular biology mechanism of different kinds of flowers in nature was explained. We plotted various patterns of flower development and deduced the function of each group genes and the phenotype of different types of mutants. It is helpful for understanding the secrets of the floral morphology.
flower structure; flower development; ABCDE model; quartet model; ectopic expression
2016-12-29;
2017-01-11
山东省自然科学基金(ZR2015DL009);研究生教育创新计划项目(SDYY16087)
孟雨婷,硕士研究生,研究方向为植物发育生物学,E-mail:172013502@qq.com
邱念伟,教授,从事植物逆境生物学研究,E-mail:nianweiqiu@163.com
10.3969/j.issn.2095-1736.2017.06.105
Q945.45
B
2095-1736(2017)06-0105-03
