中华绒螯蟹神经系统及其胚后发育的组织学观察
2017-12-18宋德磊李应东曹延超张洪艳郭恩棉
宋德磊,李应东,曹延超,张洪艳,郭恩棉
( 1.青岛农业大学 海洋科学与工程学院,山东 青岛 266109;2.沈阳农业大学 畜牧兽医学院,辽宁 沈阳 116008; 3.潍坊中晨集团,山东 潍坊 262500 )
中华绒螯蟹神经系统及其胚后发育的组织学观察
宋德磊1,李应东2,曹延超3,张洪艳1,郭恩棉1
( 1.青岛农业大学 海洋科学与工程学院,山东 青岛 266109;2.沈阳农业大学 畜牧兽医学院,辽宁 沈阳 116008; 3.潍坊中晨集团,山东 潍坊 262500 )
为完善中华绒螯蟹神经系统组织学相关资料,文章采用石蜡切片和苏木精—伊红染色技术对中华绒螯蟹神经系统及其胚后发育组织学特点进行了研究。结果表明,中华绒鳌蟹中枢神经系统在Ⅰ期溞状幼虫已见雏形,但前、中、后脑未愈合,前脑较小;Ⅱ期溞状幼虫前脑体积增大;Ⅲ期溞状幼虫中、后脑向上迁移与前脑愈合,到Ⅴ期溞状幼虫,前脑、中脑、后脑完全愈合并迁移到两眼之间;大眼幼虫和稚蟹脑具备了六边形形态。食道下神经节与胸神经节的两条神经索在Ⅰ期溞状幼虫呈分离状态,Ⅱ期溞状幼虫时开始愈合,至Ⅳ期溞状幼虫完全结合。腹神经链在Ⅰ期溞状幼虫发育不完全,Ⅲ期溞状幼虫形成两条神经索,Ⅳ期溞状幼虫两条神经索联系紧密。Ⅴ期溞状幼虫食道下神经节与胸神经节高度愈合,腹神经链发育完全, 并在大眼幼虫期开始与胸神经节愈合。胸腹神经团的3个组成部分在稚蟹期达到完全愈合。
中华绒螯蟹;中枢神经系统;胚后发育;胸腹神经团
器官组织胚后发育学对动物的生长发育起着关键作用,长期以来一直受到科学研究者的广泛重视[1-2]。其中中枢神经系统具有传递刺激、整合信息、控制运动的功能,在动物各项生命活动中发挥着十分重要的作用[3],更加吸引了广大学者的研究兴趣。中华绒螯蟹(Eriocheirsinensis)俗称河蟹或毛蟹,隶属方蟹科,绒螯蟹属。其营养丰富,味道鲜美,是一种重要的经济蟹类,在我国分布广泛,凡是通海的江河、湖泊都有出产[4]。自20世纪50年代开始,已经对中华绒螯蟹中枢神经系统的整体形态、发生发育、X器—窦腺复合体和神经分泌细胞等方面做过研究[5-11],但未见有对其中枢神经系统胚后发育组织学的相关报道。本文对中华绒螯蟹神经系统及其胚后发育情况进行了研究,以便完善中华绒螯蟹神经系统组织学相关资料,并为进一步研究其他甲壳动物神经系统的形态结构和生理功能提供参考。
1 材料与方法
试验材料取自辽宁盘锦,取样后用波恩氏液固定24 h,然后经梯度含量酒精脱水,石蜡包埋,横、纵、水平方向连续切片,苏木精—伊红染色,尼康显微镜观察并拍照。
2 结 果
2.1 中华绒螯蟹神经系统组织学
中华绒螯蟹的神经系统高度集中愈合,主要由脑神经节和胸腹神经团两大部分组成。脑神经节略呈六边形,从脑神经节的前端、两侧和后侧共发出5对主要神经,包括触角神经、视神经、皮肤神经和围食道神经。胸腹神经团呈水滴状,胸腹神经团两侧自前向后呈放射状依次发出11对附肢神经和一支腹部神经[12]。
脑神经节和胸腹神经团均由神经细胞、神经胶质细胞和疏松结缔组织组成。切片观察,明显分成内外两部分,外部染色较深,嗜碱性,称皮质。皮质内神经元胞体较多,细胞间充满了疏松结缔组织与神经胶质细胞。神经元胞体多呈圆形,大小不一,排列较为紧密,核位于细胞中央,胞质内含有较多深染的细小颗粒状尼氏体。脑神经节内部染色较浅,嗜酸性,称髓质。髓质内细胞较少,神经纤维较多。细胞主要有嗜碱性细胞、嗜酸性细胞和嫌色细胞三种,神经纤维以无髓神经纤维为主(图1a,b)。
围食道神经节的细胞多呈卵圆形或梭形,排列稀疏,树突发达但未见其轴突,各神经细胞的突起相互靠拢聚合,排列成多束细长的类纤维束结构(图1c)。

图1 中华绒鳌蟹神经系统细胞观察
a.脑神经节细胞;b. 胸腹神经团细胞;c. 围食道神经节细胞;LCT:疏松结缔组织;Ba:嗜碱性细胞;CN:细胞核;Eo:嗜酸性细胞;GC:胶质细胞;UNF:无髓神经纤维;NB:尼氏体;CC:嫌色细胞.
2.2 中华绒鳌蟹神经系统胚后发育组织学
中华绒螯蟹胚后发育阶段包括溞状幼虫Ⅰ~Ⅴ期和大眼幼虫期。期间,中华绒螯蟹神经系统发育呈现明显的阶段型差异,且与成蟹神经系统存在较大不同。
2.2.1 脑神经节胚后发育组织学
Ⅰ期溞状幼虫各部分脑均已形成,但发育极不完善,前脑较小,其髓质内的纤维分布疏松且具有较多横向的神经束,中脑与后脑位于食道的两侧,通过神经连接组成了围食道神经的一部分。前、中、后脑未愈合(图2a)。
Ⅱ期溞状幼虫前脑发育明显,体积增大,髓质变得较为密实;中、后脑的发育情况与Ⅰ期幼虫差别不大(图2b)。
Ⅲ期溞状幼虫脑进一步增大,中、后脑开始向上迁移,并与前脑愈合,其原部位留下的神经纤维束逐渐形成围食道神经(图2c)。
Ⅳ期溞状幼虫前脑各部分发育基本完善,中、后脑进一步上移至接近成蟹脑的位置,嗅叶下端与食道上端处于同一水平线上(图2d)。
Ⅴ期溞状幼虫中、后脑已迁移至成蟹脑的位置,前、中、后脑完全愈合,初步形成六边形形态(图2e)。
大眼幼虫和稚蟹时期脑进一步发育和完善,具备了六边形形态,皮质增厚,髓质神经纤维联系更为密实(图2f)。
2.2.2 胸腹神经团
Ⅰ期溞状幼虫胸腹神经团三部分未完全愈合,整体分上下两部分。上部分为食道下神经节与胸神经节愈合的部分,由左右两条神经索组成,两条神经索靠横向的神经纤维联系。下部分为游离的腹神经链,腹神经链发育不完全,皮质层细胞染色较深,髓质层极少,几乎不可见(图3a)。
Ⅱ期溞状幼虫食道下神经节与胸神经节进一步愈合,左右两条神经索也逐渐愈合。腹神经链皮质层细胞增加,呈柱状向外发散;两条纤维束状的髓质神经索依稀可见(图3b)。
Ⅲ期溞状幼虫食道下神经节与胸神经节的愈合程度进一步增加,食道下神经节与胸神经节愈合部分的两条神经索左右联系比较紧密。腹神经链左右对称的两条髓质神经索明显可见,呈鱼骨状,但两神经索之间的联系不紧密(图3c)。
Ⅳ期溞状幼虫除食道下神经节与胸神经节进一步愈合外,食道下神经节与胸神经节愈合部分的两条神经索已完全联系起来。腹神经链的两条神经索之间形成较为紧密的联系,髓质内部的无髓神经纤维和胶质细胞分布较为疏松(图3d)。
a.Ⅰ期溞状幼虫脑神经节;b. Ⅱ期溞状幼虫脑神经节;c. Ⅲ期溞状幼虫脑神经节;d. Ⅳ期溞状幼虫脑神经节;e. Ⅴ期溞状幼虫脑神经节;f. 大眼幼虫脑神经节;OL:嗅叶;Pb:前脑;Es:食道;OG;视神经节;Tb:后脑; Db:中脑.
Ⅴ期溞状幼虫食道下神经节与胸神经节高度愈合。腹神经链的发育程度基本完善,其皮质外侧与五条神经联系,髓质位于腹神经链的中央,呈鱼骨状,内部充满了无髓神经纤维和胶质细胞(图3e)。
大眼幼虫腹神经链开始向上与食道下神经节和胸神经节形成愈合(图3f)。
稚蟹腹神经链、食道下神经节和胸神经节三者完全愈合,胸腹神经团正式形成(图3g)。
3 讨 论
中华绒螯蟹中枢神经系统的形态结构特点与机体的形态结构有着密切关系,其溞状幼虫与大眼幼虫中枢神经系统均呈梯形,稚体期变为六边形,可能和溞状幼虫与大眼幼虫腹部均未退化,在体形上相似有关。中华绒螯蟹Ⅰ期溞状幼虫视神经节基本发育完全,且与各附肢相对应的运动神经也均在溞状幼虫期发育完善,这与同为甲壳动物的中国明对虾[7,12](Fenneropenaeuschinensis)相似;但其胸神经节和食道下神经节的出现和愈合却要早于中国明对虾[12-13],这表明节肢动物神经系统的发育不仅与机体的形态结构有关,而且与其生理活动也存在着密不可分的联系。
中华绒螯蟹神经系统的大部分结构在Ⅰ期溞状幼虫已经形成,但尚未完全愈合。Ⅰ期溞状幼虫腹神经链还处于发育早期,髓质层极少,几乎不可见,直至Ⅲ期溞状幼虫腹神经链的左右两条神经索才明显可见。这与蛛形蛤蟆蟹(Hyasaraneus)Ⅰ期溞状幼虫腹神经链的左右两条神经索虽未联合但已形成且清晰可见[14]不同,更与三疣梭子蟹(Portunustrituberculatus)在胚胎发育至卵内溞状幼体阶段,其腹神经链的左右两条神经索已经形成[15]存在着较大差异。中华绒螯蟹的胸腹神经团中,食道下神经节与胸神经节细胞相对较多,且发出多对神经;而腹神经节细胞相对较少,仅向后发出一条腹神经。这与其他蟹类[12,16]相似,可能和蟹类头胸部发达而腹部退化的特殊体型密切相关。
中华绒螯蟹中枢神经系统胸腹神经团整体略呈水滴状且中央具有一个胸动脉孔,这与三疣梭子蟹[12]的胸腹神经团结构高度相似,而与锯缘青蟹(Scyllaserrata)[16]成体的左右对称圆盘状并拥有左右对称的两个胸动脉孔的胸腹神经团存在较大的差异。锯缘青蟹胸腹神经团的结构很可能与其硕大的圆盘状体型相关,据陈宽智[17]对蟹类循环系统的研究证明,蟹类胸动脉分支为胸下动脉和腹下动脉分别进入胸部附肢和腹部器官这一点来看,锯缘青蟹发达的胸动脉很可能与其发达的螯足相关,但这种差异是否存在其他生物学上的联系还需进一步的研究和探讨。
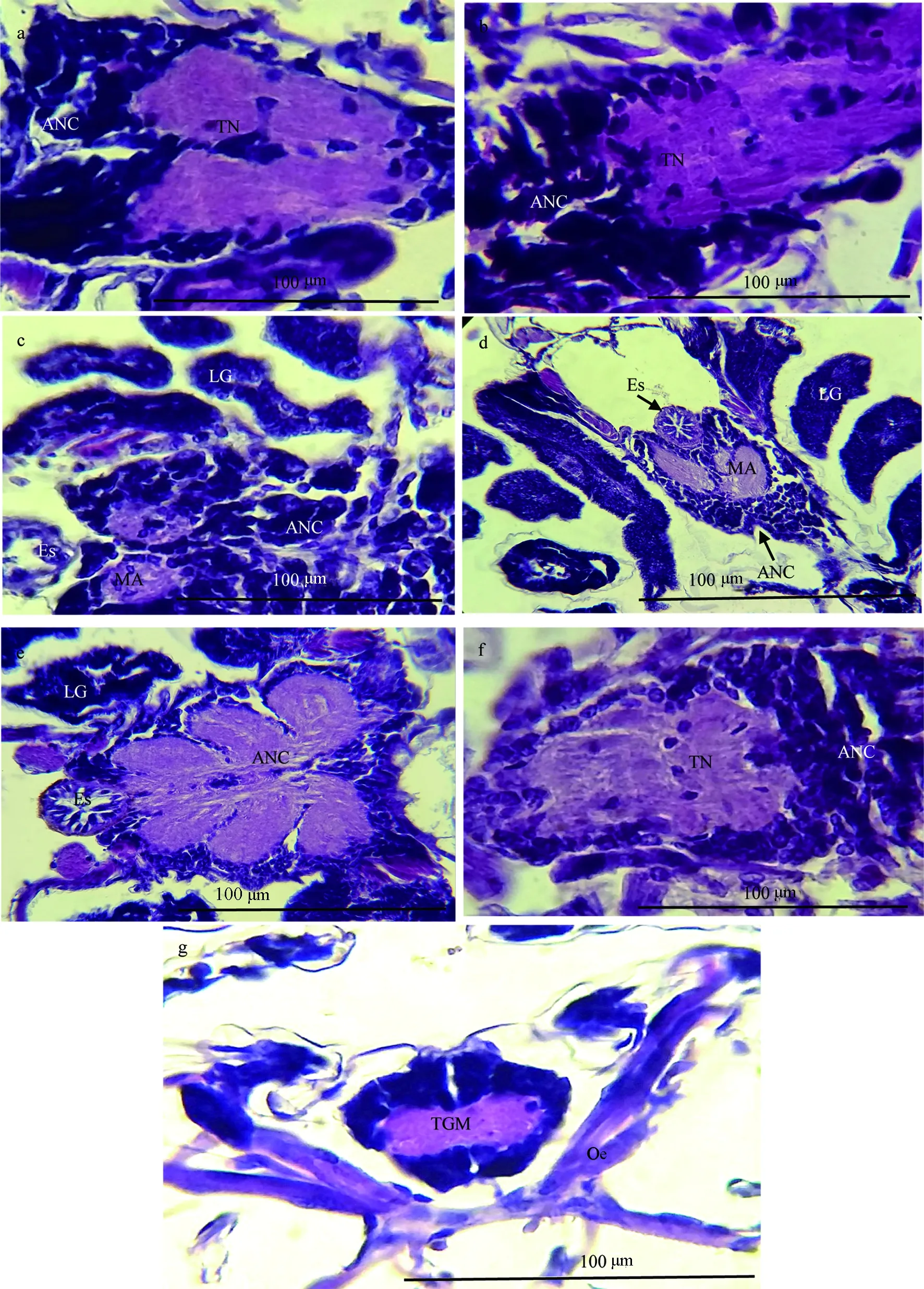
图3 溞状幼虫、大眼幼虫和稚体的胸腹神经团组织学
a. Ⅰ期溞状幼虫胸腹神经团; b. Ⅱ期溞状幼虫胸腹神经团; c. Ⅲ期溞状幼虫胸腹神经团; d. Ⅳ期溞状幼虫胸腹神经团; e. Ⅴ期溞状幼虫胸腹神经团; f. 大眼幼虫胸腹神经团; g. 稚体胸腹神经团;ANC:腹部神经链;TN:胸神经节;Es:食道;MA:腹神经链髓质;LG:步足神经;TGM:胸腹神经团;Oe:围食道神经.
[1] 白雪峰, 李海东, 孙虎山. 分叉小猛水蚤消化和神经系统的组织学研究[J]. 水产科学, 2014, 33(9):583-586.
[2] 孙德兴, 赵兴文, 步海平, 等. 泥鳅消化器官胚后发育组织学[J]. 水产科学, 2015, 34(11):696-701.
[3] 李霞. 水产动物组织胚胎学[M]. 北京:中国农业出版社, 2005:291-292.
[4] 赵文. 水生生物学[M]. 北京:中国农业出版社, 2005:146-147.
[5] 梁箫, 沈春燕, 刘志强, 等. 中华绒螯蟹脑和胸腹神经团的组织学观察[J]. 上海海洋大学学报,2014,10 (3):174-175.
[6] 王文, 朱宁宁, 李正荣, 等. 类立克次体侵染中华绒螯蟹神经组织的光镜和电镜观察[J]. 动物学研究, 2001, 22(6):467-471.
[7] 赵云龙, 李红, 王群, 等. 中华绒螯蟹神经细胞和胶质细胞的光镜及电镜观察[J]. 动物学研究,1999, 20(6):411-414.
[8] 韩青动, 袁春营, 崔青曼, 等. 中华绒螯蟹脑神经分泌细胞的研究[J]. 水利渔业, 2006, 26(5):8-10.
[9] 吴江立, 穆淑梅, 郭明伸, 等. 中华绒螯蟹眼柄视神经节发育特征的初步研究[J]. 河北渔业, 2011, 39(1):13-19.
[10] 袁春营, 崔青曼, 韩青动. 中华绒螯蟹胸神经团神经分泌细胞的显微和超显微结构观察[J]. 动物学杂志, 2006, 41(3):75-79.
[11] 赵景霞. 中华绒螯蟹眼柄端髓X 器官神经分泌细胞钙激活钾通道研究[J]. 水产学报, 2007, 31(4):417-422.
[12] 陈宽智. 东方对虾(Penaeusorientalis)和三疣梭子蟹(Portunustrituberculatus)中枢神经系统解剖及十足目动物腹神经的形态比较[J]. 山东海洋学院学报, 1980, 10(3):91-99.
[13] 张建. 中国明对虾(Fenneropenaeuschinensis)中枢神经系统结构发生和分化的细胞学观察[D].青岛:中国海洋大学, 2004.
[14] Harzsch S, Dawirs R R. Development of the CNS in spider crab larvae [J]. Zoea Newsletter, 1994, 1(2):8-10.
[15] 薛俊增. 三疣梭子蟹胚胎期中枢神经系统的发生和发育[J]. 动物学研究, 2005, 26(2):184-189.
[16] 叶海辉, 李少菁, 金朱兴, 等. 锯缘青蟹胸神经团的组织学研究[J]. 厦门大学学报:自然科学版, 2002, 41(4):502-503.
[17] 陈宽智. 十足目动物(甲壳纲)的循环系统[J]. 动物学杂志, 1982(1):1-3.
HistologicalObservationofNervousSystemandPostembryonicDevelopmentofChineseMittenCrabEriocheirsinensis
SONG Delei1, LI Yingdong2, CAO Yanchao3, ZHANG Hongyan1, GUO Enmian1
( 1. College of Marine Science and Engineering, Qingdao Agricultural University, Qingdao 266109, China; 2.College of Animal Husbandry and Veterinary Medicine, Shenyang Agricultural University, Shenyang 116008, China; 3. Zhongchen Industry Association of Weifang, Weifang 262500, China )
Histological features of nervous system and postembryonic development were studied in Chinese mitten crabEriocheirsinensisby paraffin section and HE staining. The results showed that in spite of separation of the forebrain, midbrain and hindbrain, various parts of central nervous system had formed in phase I of the zoea larvae with a smaller forebrain. The brain volume increased in phase Ⅱ of the zoea. The midbrain and hindbrain migrated upward to be combined with forebrain in phase Ⅲ of the zoea. In phase Ⅴ of the zoea larvae, three parts of the brain healed completely and located between the eyes. A hexagon-shaped brain formed in the megalopa larvae and the juvenile crab. Two nerve cords of lower esophageal ganglia and the thoracic ganglia separated in phase I of the zoea, which began to combine in phase Ⅱ of the zoea and fully integrated in phase Ⅲ of the zoea. Ventral nerve chain developed incompletely in phase I of the zoea and formed two nerve cords in phase Ⅲ of the zoea. Two nerve cords contacted closely in phase Ⅳ of the zoea. The lower esophageal ganglia and the thoracic ganglia fully developed in phase Ⅴ of the zoea. In the megalopa larvae, ventral nerve chain began to heal with thoracic ganglia. Three components of abdominal nerve group were fully consolidated in the juvenile crab.
Eriocheirsinensis;central nerve system;postembryonic development;abdominal ganglion
10.16378/j.cnki.1003-1111.2017.02.011
2016-02-01;
2016-05-10.
2015年国家级大学生创新训练项目(9317).
宋德磊(1996—),男,本科生;研究方向:水产养殖研究. E-mail:1344050503@qq.com. 通讯作者:郭恩棉(1972—),女,副教授,硕士;研究方向:水产养殖教学与研究. E-mail:emguo2013@126.com.
Q954.6
A
1003-1111(2017)02-0183-05
