301份小麦种质醇溶蛋白遗传多样性及其与品质性状的相关性分析
2017-12-07姜小苓张自阳李小军于红彩李秀玲茹振钢
姜小苓 张自阳 李小军 李 淦 于红彩 李秀玲 茹振钢
(河南科技学院小麦中心;河南省现代生物育种协同创新中心; 河南省高校作物分子育种重点开放实验室,新乡 453003)
301份小麦种质醇溶蛋白遗传多样性及其与品质性状的相关性分析
姜小苓 张自阳 李小军 李 淦 于红彩 李秀玲 茹振钢
(河南科技学院小麦中心;河南省现代生物育种协同创新中心; 河南省高校作物分子育种重点开放实验室,新乡 453003)
为探讨小麦醇溶蛋白的遗传多样性及其亚基对品质性状的影响,利用酸性聚丙烯酰胺凝胶电泳(A-PAGE)技术对不同来源的301份小麦品种(系)的醇溶蛋白组成进行分析。结果表明,301份材料共分离出93种不同迁移率的谱带,其中迁移率为58.6、69.4、72.1、16.5和19.2的谱带出现频率最高(均高于80.0%),其余谱带的多态性相对较高。品种间遗传相似系数(GS)的变异范围为0.538~1.000,平均为0.759,中原008和中育1401的亲缘关系最近(GS值为1.0)。在GS=0.740水平上,将所有材料划分为11个类群。相关分析表明,42条谱带与106项次品质性状呈显著或极显著相关,其中谱带ω 25.5、γ 43.7和γ 48.2可显著增加面团稳定时间、面筋指数;谱带γ 52.2和α 80.7可显著增加粗蛋白及干、湿面筋含量;谱带β 59.2可显著提高小麦粉色泽L*值,但与b*值负相关;谱带ω 34.8、ω 36.6和ω 39.3对面团稳定时间及面筋指数具有显著的负向效应。这些重要的醇溶蛋白谱带可作为小麦品质育种的选择标记。
小麦 醇溶蛋白 遗传多样性
醇溶蛋白和谷蛋白是小麦的主要储藏蛋白,约占籽粒蛋白的80%,是面筋的主要成分,其数量和比例决定了面筋的质量[1]。其中,醇溶蛋白约占小麦籽粒蛋白的40%,与面团的黏性密切相关,对烘烤品质有重要作用[2]。醇溶蛋白由小麦第1、第6部分同源群染色体短臂上的 Gli-1和 Gli-2位点编码,其中Gli-1位点包括 Gli-A1、Gli-B1和Gli-D1;Gli-2位点包括Gli-A2、Gli-B2和Gli-D2。这些位点上等位基因的高度变异及位点间不同等位基因组合使小麦醇溶蛋白表现出高度多态性[3]。迄今,在普通小麦的 Gli-1和 Gli-2位点上鉴定出130个等位变异[4]。大量研究表明,小麦醇溶蛋白在不同品种间存在显著差异,其电泳条带的数量及组合方式完全受基因调控,基本不受环境影响,常被用于小麦品种鉴定、纯度检测、亲缘关系分析及遗传多样性研究[5]。
国内外学者对小麦谷蛋白亚基的组成及其与品质关系做了大量研究[6-9],发掘出诸如5+10、14+15等优质亚基组合,但有关小麦醇溶蛋白多态性及其与品质关系的报道较少。例如,Branlard等[10]以70份小麦品种为材料分析了醇溶蛋白的品质效应,发现醇溶蛋白谱带α72.5、α74、α76、α84对面筋的弹性、膨胀性、黏性以及延展性具有正向效应。张平平等[11]分析了33份春小麦的醇溶蛋白组成及其对品质性状的影响,结果表明,ω、γ1和γ型醇溶蛋白与沉降值呈显著正相关,γ2和γ型醇溶蛋白与面团形成时间呈显著正相关,而ω和γ型醇溶蛋白与面团延伸性呈显著负相关。阎旭东等[12]报道醇溶蛋白谱带2.3、62.7、39.6(5)、11.4和23等对沉降值有正向效应,γ 44.5和γ 45.0对面条加工品质有正向效应,而γ 41表现负向效应。
通过A-PAGE技术对来源于国内外不同生态区的小麦主推品种、地方品种或高代品系共301份材料进行醇溶蛋白遗传多样性分析,并探讨了其蛋白亚基与小麦品质的关系,以期为小麦品质育种及分子标记辅助选择提供参考。
1 材料与方法
1.1 试验材料
参试材料为301份不同来源的小麦主推品种、地方品种或高代品系,其中国外41份,国内260份。国内品种(系)包括黄淮冬麦区169份、长江中下游冬麦区33份、北方冬麦区23份、西南冬麦区25份及来自其他麦区10份,所有材料均由河南科技学院小麦中心提供。以Marquis、Neepawa和中国春作为对照品种。
1.2 试验方法
1.2.1 醇溶蛋白组成分析
醇溶蛋白提取和分离参照尹燕枰等[13]的方法。
1.2.2 品质指标测定方法
利用LRMM 8040-3-D实验磨粉机(江苏无锡锡粮机械制造公司)制取小麦粉,出粉率65%左右;利用SBDY-1数显白度仪(上海悦丰仪器公司)检测小麦粉白度;利用UDK159凯氏定氮仪(意大利VELP公司)测定粗蛋白含量,方法参照GB/T 55ll-2008/ISO 20483:2006;利用WZZ-2S/2SS旋光仪(上海易测仪器设备有限公司)测定小麦粉的粗淀粉含量,方法参照GB/T 20378-2006/ISO 10520:1997;利用CR-400色彩色差计(日本美能达公司)测定小麦粉色泽;利用810101粉质仪(德国Brabender公司)测定面团粉质特性,方法参照GB/T 14614—2006;利用2200面筋仪(瑞典波通公司)测定面筋含量及面筋指数,方法参照SB/T10249-95。
1.2.3 统计分析方法
以小麦品种Marquis的谱带为对照,参照Bushuk[14]的方法,利用Excel编辑公式计算谱带的相对迁移率。材料间遗传相似系数参照Nei的[15]方法计算,公式为:GS=2Nij/(Ni+Nj),其中Ni为i材料出现的谱带数,Nj为j材料出现的谱带数,Nij为二者共有的谱带数。根据相对迁移率及品质数据用SPSS 20.0进行相关性分析;采用NTsys-pc 2.10软件,根据UPGMA(不加权成对算数平均法)方法进行聚类分析。
2 结果与分析
2.1 参试材料的醇溶蛋白组成及遗传多样性分析
301份小麦材料的醇溶蛋白表现出丰富的遗传变异,共分离出93条迁移率不同的醇溶蛋白谱带(编号为1~93),谱带频率变异范围为0.33%~98.01%(表1)。其中编号为68、82和85的谱带出现频率最高,其次是编号6和9的谱带,说明这些醇溶蛋白谱带多态性低。另外,6号和9号谱带为双联共显带(图1),有245个材料(占81.4%)出现该谱带。所有谱带中,有34条迁移率不同的谱带频率小于10%,尤其是编号67和80的谱带仅分别在品系CP02-3-5-5和品种绵阳39中出现,可作为该品种(系)的特征带;分离出的93条迁移率不同的谱带没有1条同时在301份参试材料中出现。

表1 不同醇溶蛋白谱带在参试材料的出现频率
由表2可知,301份材料共分离出6 337条醇溶蛋白谱带,单个材料谱带数的变异范围为13~30条,平均为21.05条。多数材料的谱带数为17~25条,占84.39%;21个材料的谱带数少于17条,占6.98%,小麦品系农大8P291谱带数最少,仅有13条;26个材料的谱带数多于25条,占8.64%,来自墨西哥的品系墨176谱带数最多(30条)。表明供试材料的醇溶蛋白编码基因存在着广泛的遗传变异,遗传多样性丰富。
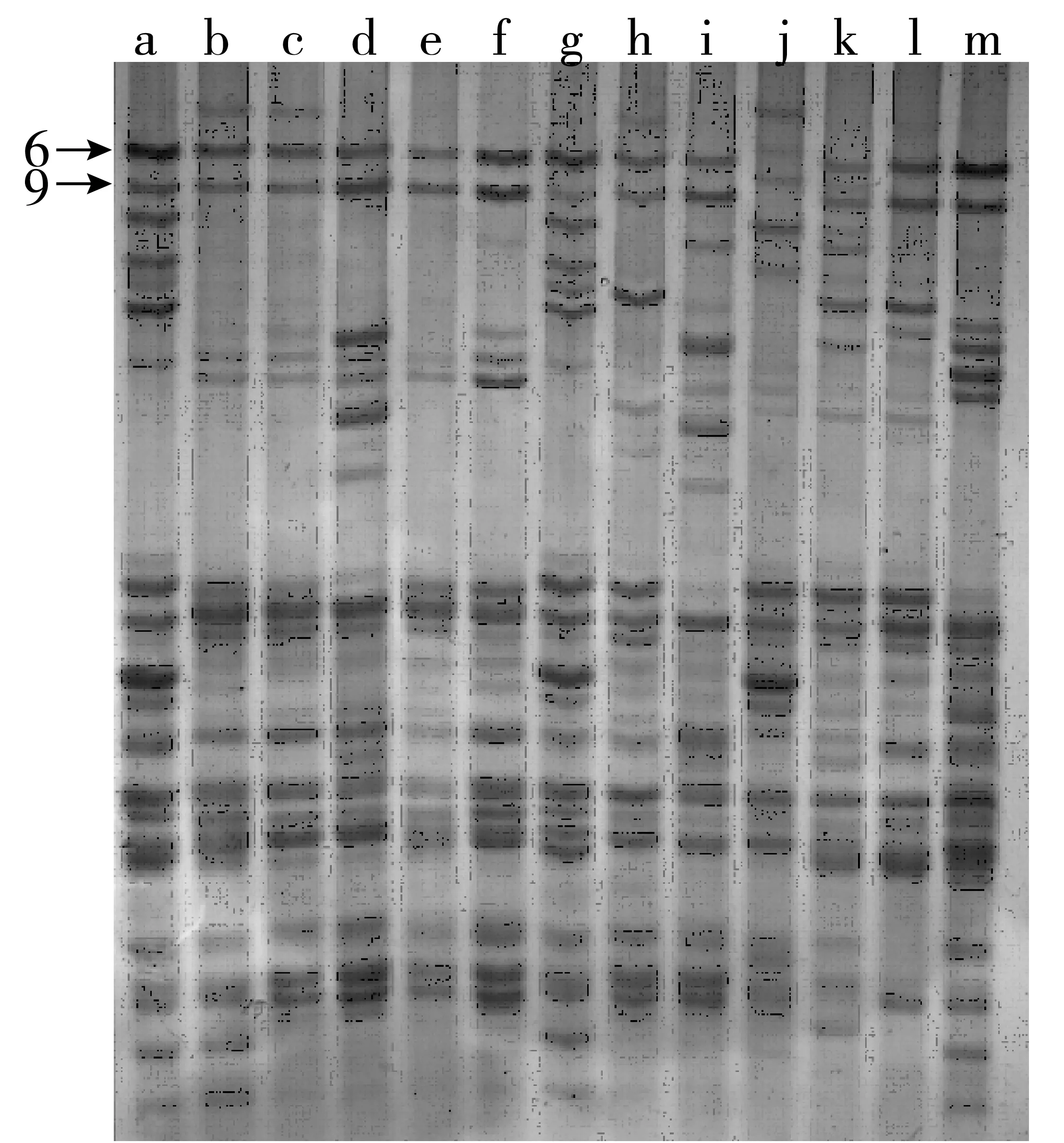
注:a.Marquis;b.百农003171;c.漯2558;d.济创28号;e.驻麦6号A;f.MD8714;g.Neepawa;h.KPL-7;i.泛麦5号;j.KPL-5;k.郑州8960;l.6S139;m.中国春。图1 部分参试材料的醇溶蛋白电泳图谱
表2 参试材料醇溶蛋白谱带数统计分析结果
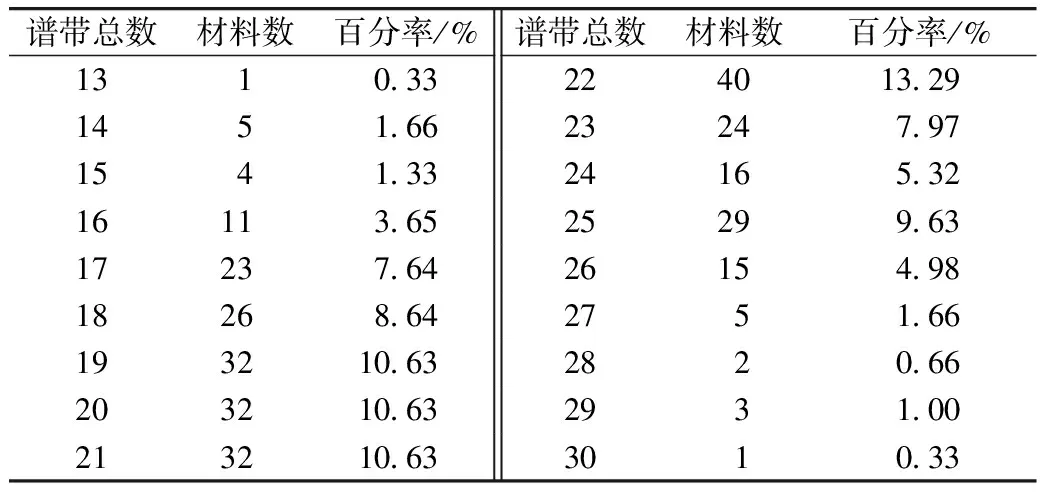
谱带总数材料数百分率/%谱带总数材料数百分率/%131033224013291451662324797154133241653216113652529963172376426154981826864275166193210632820662032106329310021321063301033
2.2 遗传相似性分析
从301份参试材料中选取251份代表性材料进行遗传相似性分析,31 375个醇溶蛋白遗传相似系数(GS)变异范围为0.538~1.000,平均为0.759。GS值次数分布分析表明,GS值主要集中在0.72~0.82范围内,占76.92%;GS值小于0.64的仅有222个,0.71%,而GS值大于0.86的有621个,占1.98%(图2)。引自国外的品系墨176与来自黄淮冬麦区的品系6S139间的遗传相似系数最小(0.538),表明二者的亲缘关系较远;而来自黄淮冬麦区的小麦品种中原008和中育1401之间的遗传相似系数最高(1.000),说明这两个品种的亲缘关系非常近。
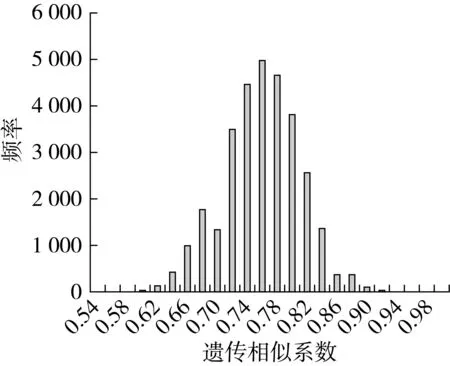
图2 品种(系)间遗传相似系数的次数分布
2.3 聚类分析
对251份材料醇溶蛋白间的GS值进行聚类分析(图略),在GS=0.740水平上,251份材料可划分为11个类群。第一类包含15个材料;第二类包含201个材料,它们又可分为5个亚类;第三类包含21个材料;第四、第五、第十类群各包含1个材料;第六类包含3个材料;第七、第八、第九、第十一类各包含两个材料。整体来看,聚类分析结果与品种的系谱关系基本一致,例如来自河南的周麦系列、百农系列、泛麦系列等品种均聚在第二大类。但也出现个别亲缘关系较远的品种聚类较近的情况,例如来自美国的品系“美国-2”与国内优质面包小麦济南17被聚在一类。
2.4 参试材料品质表现
由表3可知,a*值、b*值、形成时间、稳定时间、粉质质量指数及面筋指数的变异系数均较高(大于30%),说明这些品质参数变异丰富,有利于通过育种手段对其进行改良,而白度、粗蛋白、淀粉、L*值及吸水率的变异系数均小于10%。湿面筋、面筋指数和面团稳定时间等评价面团筋力的主要品质性状在品种(系)间的变异幅度均较大,分别为21.6%~42.1%、3.2%~98.5%和1.0~41.2 min;白度变异范围达到54.7~83.3,表明这些材料的筋力类型及小麦粉白度变异广泛。
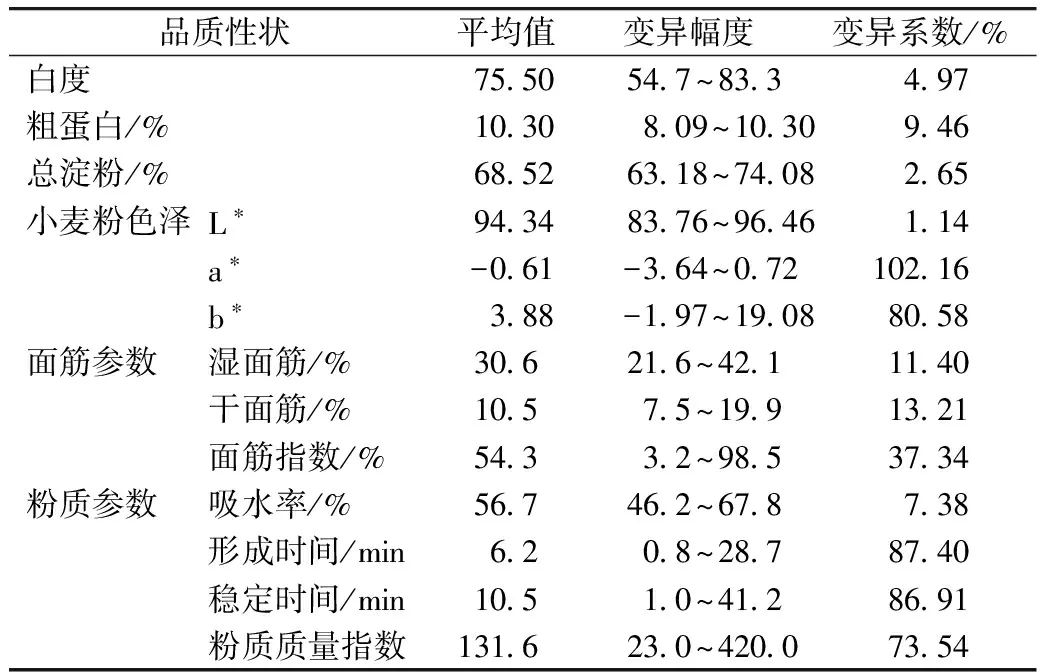
表3 参试材料品质表现
2.5 醇溶蛋白亚基与品质性状的相关性分析
2.5.1 醇溶蛋白亚基与小麦粉理化品质的相关性分析
共有23条谱带与36项次小麦粉理化品质指标呈显著或极显著相关(表4)。其中,迁移率为19.2、56.2、75.4的谱带可增加小麦粉白度,迁移率为29.0、30.6、80.7的谱带可提高粗蛋白含量,迁移率为86.6的谱带可提高淀粉含量,迁移率为59.2和86.6的谱带可提高小麦粉亮度(L*值),这些谱带对小麦粉理化品质均具有正向效应。然而迁移率为35.7、58.6、74.1、71.5、37.6、52.2的谱带对小麦粉理化品质具有负向效应。
2.5.2 醇溶蛋白亚基与面团粉质特性及面筋参数的相关性分析
共有32条谱带与70项次面团粉质及面筋参数呈显著或极显著相关(表5)。迁移率为12.2、23.9、25.5、31.1、42.3、43.7、45.2、46.6、48.2、52.2、52.8、63.0、64.8、80.7等的谱带表现正向效应。其中,迁移率为12.2、45.2、80.7的谱带可增加湿面筋含量,迁移率为23.9、48.2、52.8和63.0的谱带可延长面团稳定时间,迁移率为31.1的谱带可增加吸水率,迁移率为25.5、42.3、43.7的谱带可延长面团形成时间和稳定时间、提高面筋指数;迁移率为46.6、64.8和52.2的谱带可提高干、湿面筋含量。相反,其余的醇溶蛋白谱带对面团粉质和面筋参数表现负向效应。
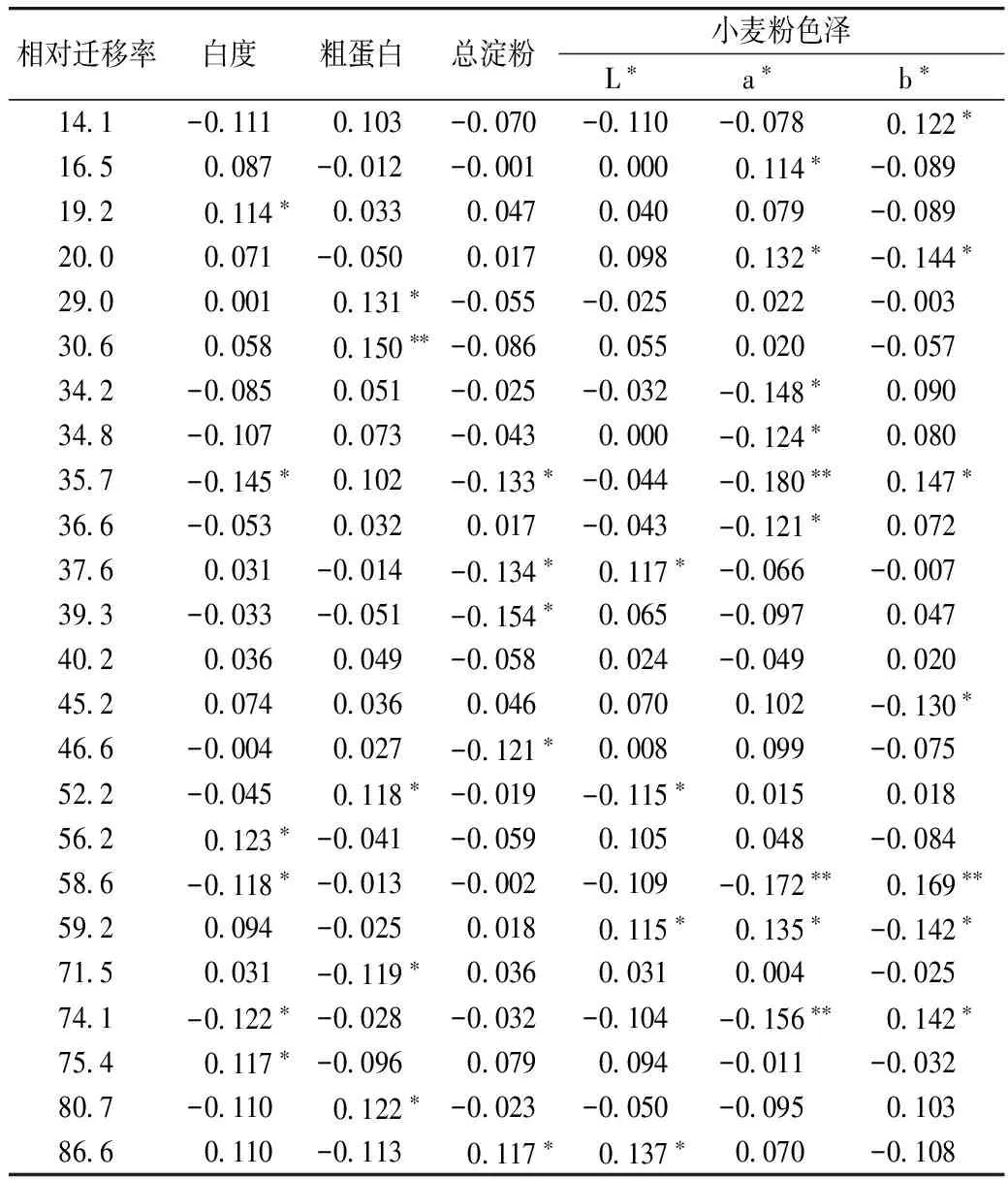
表4 部分醇溶蛋白谱带与小麦粉理化品质的简单相关系数
注:*和**分别表示0.05和0.01的显著水平,余同。
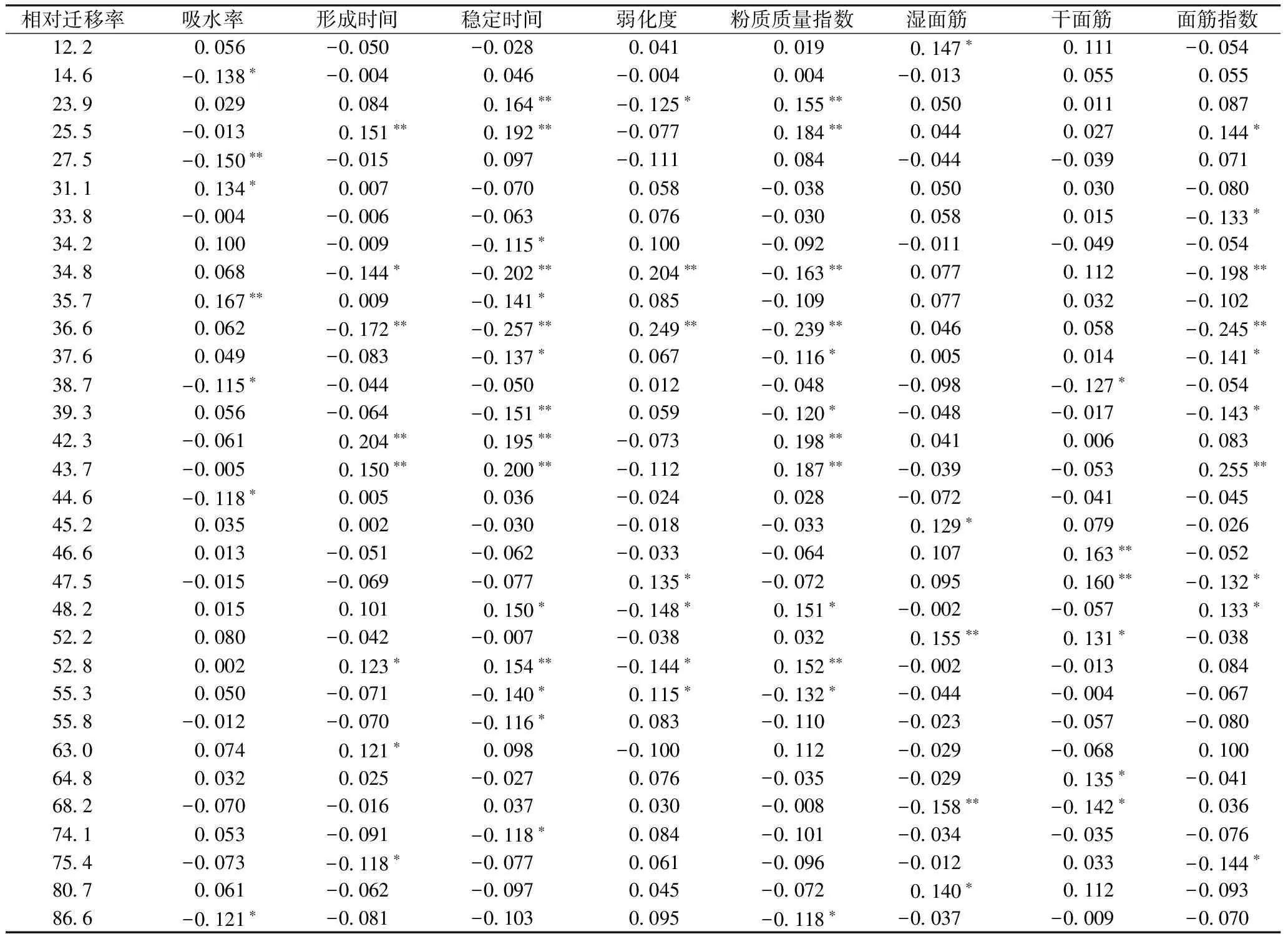
表5 部分醇溶蛋白谱带与面团粉质特性及面筋指标的简单相关系数
2.5.3 重要谱带的品质效应分析
研究共发现42条醇溶蛋白谱带与小麦品质显著相关,从中选取9条同时与多个品质性状密切相关的特征带,并进一步分析其品质效应(表6)。整体来看,具有特征谱带的材料与没有该谱带的材料在相关品质性状上存在显著差异。例如,特征带ω 25.5对面团稳定时间表现正向效应,携带该特征带的材料有67个,占材料总数的22.3%,面团稳定时间平均为13.8 min,显著(P<0.05)高于没有该特征带的材料(平均为9.6 min);特征带ω 36.6与面筋指数呈负相关,有72个材料携带该特征带(占总数的23.9%),面筋指数均值为45.5%,显著(P<0.05)低于没有该特征带的材料(均值为57.1%)。这些重要的谱带可为小麦品质育种提供遗传标记。
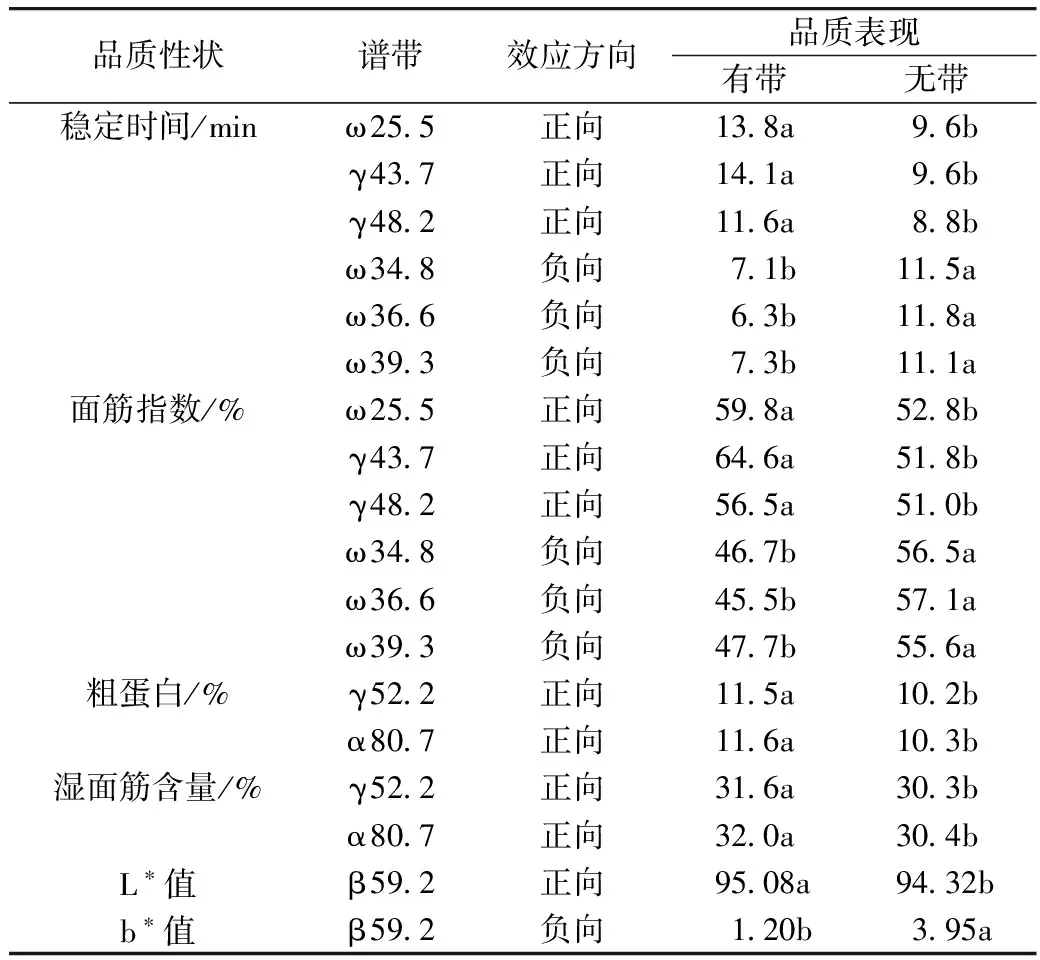
表6 重要醇溶蛋白谱带的品质效应
注:同行不同小写字母表示在5%水平存在显著性差异。
3 讨论
3.1 醇溶蛋白的遗传多样性
小麦优异种质资源发掘是改良小麦品质、提高产量的重要途径[16]。本研究对301份小麦品种(系)的醇溶蛋白组成进行分析,除中原008和中育1401的醇溶蛋白带型完全一致外,其他材料的带型各不相同。另外,参试材料GS值的变异范围为0.538~1.000,平均为0.759,与前人研究结果基本一致[17-18];中原008和中育1401的遗传相似性最大(GS值为1.0),其余材料间的GS值均小于0.978。以上分析表明,参试材料在醇溶蛋白水平存在丰富的遗传多样性,可作为小麦遗传改良的重要基因资源。
聚类分析结果表明,系谱关系相近的材料被聚在一类;但也出现个别亲缘关系较远的品种聚类较近的情况,例如来自美国的品系“美国-2”与国内优质面包小麦济南17被聚在一类;也存在亲缘关系较近却聚类相对较远的情况,例如鲁麦14与其衍生品种济麦20聚类较远。王正阳等[19]也曾报道,可能是在品种选育过程中,醇溶蛋白编码基因发生了重组,以及育种家不同的育种目标和选择倾向,导致醇溶蛋白编码基因位点发生了遗传变异[20]。
3.2 醇溶蛋白亚基与小麦品质的相关性
国内外学者相继报道了一些与小麦品质密切相关的醇溶蛋白谱带[21-24],比较发现有些结果与本研究基本一致。例如,本研究发现迁移率为34.8和36.6的谱带对稳定时间和面筋指数表现负效应,王曙光等[24]报道该谱带与沉降值、蛋白质和湿面筋含量呈负相关;迁移率为42.3的谱带与稳定时间和粉质质量指数呈正相关,阎旭东等[12]报道该谱带与沉降值正相关;迁移率为47.5的谱带可显著降低面筋指数,郭超等[17]、Branlard等[25]证实该谱带与面团延伸面积以及面筋弹性、膨胀性、黏性等品质性状呈负相关。本研究中其他醇溶蛋白谱带与前人研究尚缺乏较好的一致性,可能是不同学者所用的电泳方法特别是凝胶浓度不同,导致不同研究报道中,与品质相关的醇溶蛋白谱带很难一一对应[24],另一方面,醇溶蛋白各位点等位基因的分布具有明显的地域性,不同国家和地区的品种在醇溶蛋白存在的巨大差异也是结果不一致的重要原因[26]。此外,醇溶蛋白的多态性只是代表了小麦第1、第6部分同源群染色体短臂的遗传变异,并不能反映小麦整个基因组的遗传信息。因此,还需进一步结合DNA分子标记进行深入的分析研究。
4 结论
301份参试材料醇溶蛋白谱带丰富,品种间差异明显,所有材料中只有来自河南的中原008和中育1401的带型完全一致,其他材料的带型各不相同;聚类结果与品种的系谱关系基本一致;检测到一批与小麦品质密切相关的醇溶蛋白谱带,可作为小麦品质育种的选择标记。
[1]李敏,高翔,陈其皎,等.普通小麦中α-醇溶蛋白基因(GQ891685)的克隆、表达及品质效应鉴定[J].中国农业科学,2010,43(23):4765-4774
Li M,Gao X,Chen Q J,et al.Cloning,prokaryotic expression and in vitro functional analysis of α-gliadin gene from common wheat[J].Scientia Agricultura Sinica,2010,43(23):4765-4774
[2]Cox T S,Lookhart G L,Walker D E,et al.Genetic relationships among hard red winter wheat cultivars as evaluated by pedigree analysis and gliadin polyacrylamide gel eletrophretic patterns[J].Crop Science Society of America,1985,25:1058-1063
[3]Metakosky E V,Branlard G.Genetic diversity of French common wheat germplasm based on gliadin alleles[J].Theroretical and Applied Genetics,1998,96:209-218
[4]Metakovsky E V.Gliadin allele identification in common wheat.Ⅱ Catalogue of gliadin alleles in common wheat[J].Journal of Genetics and Breeding,1991,45:325-344
[5]张学勇,杨欣明,董玉琛.醇溶蛋白电泳在小麦种质资源遗传分析中的应用[J].中国农业科学,1995,28(4):25-32
Zhang X Y,Yang X M,Dong Y C.Genetic analysis of wheat germplasm by acid polyacrylamide gel electrophoresis of gliadins[J].Scientia Agricultura Sinica,1995,28(4):25-32
[7]Li Y L,Huang C Y,Sui X X,et al.Genetic variation of wheat glutenin subunits between landraces and varieties and their contributions to wheat quality improvement in China[J].Euphytica,2009,169:159-168
[8]Terasawa Y,Takata K,Hirano H,et al.Genetic variation of high-molecular-weight glutenin subunit composition in Asian wheat[J].Genetic Resources and Crop Evolution,2011,58:283-289
[9]张玲丽,李秀全,杨欣明,等.小麦优良种质资源高分子量麦谷蛋白亚基组成分析[J].中国农业科学,2006,39(12):2406-2414
Zhang L L,Li X Q,Yang X M,et al.High-molecular-weight glutenin subunit composition of Chinese wheat germplasm[J].Scientia Agricultura Sinica,2006,39(12):2406-2414
[10]Branlard G,Dardevet M,Saccomano R,et al.Genetic diversity of wheat storage proteins and bread wheat quality[J].Euphytica,2001,119:59-67
[11]张平平,陈东升,张勇,等.春播小麦醇溶蛋白组成及其对品质性状的影响[J].作物学报,2006,32(12):1796-1801
Zhang P P,Chen D S,Zhang Y,et al.Gliadin composition and their effects on quality properties in spring wheat[J].Acta Agronomica Sinica,2006,32(12):1796-1801
[12]阎旭东,卢少源,李宗智.普通小麦醇溶蛋白组份的分布及其与HMW-麦谷蛋白亚基对品质的组合效应[J].作物学报,1997,23:70-75
Yan X D,Lu S Y,Li Z Z.The distribution of gliadin composition and its interaction with HMW-glutenin subunits on breadmaking quality of common wheat[J].Acta Agronomica Sinica,1997,23:70-75
[13]尹燕枰,董学会.种子学试验技术[M].北京:中国农业出版社,2008:181-184
Yin Y P,Dong X H.The Experiment Technology of Seed Science[M].Beijing:China agriculture press,2008:181-184
[14]Bushuk W,Zillman R R.Wheat cultivar identification by gliadin electrophoregrams.I.Apparatus,method and nomenclature[J].Canadin Journal of Plant Science,1978,58:505-515
[15]Nei M.Analysis of gene diversity in subdivided populations[J].Proc Natl Acad Sci USA,1973,70(12):3321-3323
[16]胡琳,许为钢,张磊,等.小麦种质资源鉴定、优异基因发掘及创新利用研究概述[J].河南农业科学,2009(9):22-25
Hu L,Xu W G,Zhang L,et al.Identification,exploration,innovation and utilization of elite germplasm and gene resources in wheat[J].Journal of Henan Agricultural Sciences,2009(9):22-25
[17]郭超,刘红,陈新宏,等.部分美国小麦种质资源醇溶蛋白遗传多样性分析及其亚基对品质性状的影响[J].植物遗传资源学报,2014,15(6):1173-1181
Guo C,Liu H,Chen X H,et al.Genetic diversity of gliadin in some American wheat germplasms and their effect on quality properties[J].Journal of Plant Genetic Resources,2014,15(6):1173-1181
[18]陈晓杰,吉万全,王亚娟.新疆冬春麦区小麦地方品种贮藏蛋白遗传多样性研究[J].植物遗传资源学报,2009,10(4):522-528
Chen X J,Ji W Q,Wang Y J.Genetic diversity analysis on the storage proteins of wheat landraces from Xinjiang wheat production region[J].Journal of Plant Genetic Resources,2009,10(4):522-528
[19]王正阳,倪永静,牛吉山,等.99 份国内小麦新品种(系)醇溶蛋白的遗传多样性分析[J].麦类作物学报,2010,30(2):233-239
Wang Z Y,Ni Y J,Niu J S,et al.Analysis of the genetic diversity at gliadin loci in 99 new domestic wheat cultivars or lines[J].Journal of Triticeae Crops,2010,30(2):233-239
[20]刘华,王宇生,张辉,等.小麦种质资源醇溶蛋白指纹图谱数据库的初步建立及应用[J].作物学报,1999,25(6):674-682
Liu H,Wang Y S,Zhang H,et al.Preliminary construction and application of gliadin fingerprints database of Chinese wheat germplasm[J].Acta Agronomica Sinica,1999,25(6):674-682
[21]Metakovsky E V.Gliadin allele identification in common wheat.ⅡCatalogue of gliadin alleles in common wheat[J].Journal of Genetics and Breeding,1991,45:325-344
[22]Wrigley C W,Robinson P J,Williams W T.Associations between individual gliadin proteins and quality,agronomic and rnorphological attributes of wheat cultivars[J].Australian Journal of Agricultural Research,1982,33:409-418
[23]聂莉,芦静,黄天荣,等.部分新疆小麦材料的醇溶蛋白组成及其对品质性状的影响[J].麦类作物学报,2010,30(4):749-754
Nie L,Lu J,Huang T R,et al.Gliadin composition in Xinjiang wheat and their effect on quality properties[J].Journal of Triticeae Crops,2010,30(4):749-754
[24]王曙光,杨海峰,孙黛珍,等.小麦醇溶蛋白亚基与品质性状的相关性分析[J].中国粮油学报,2013,28(5):31-35
Wang S G,Yang H F,Sun D Z,et al.Analysis of correlation between gliadin subunits and quality characters in wheat[J].Journal of the Chinese Cereals and Oils Association,2013,28(5):31-35
[25]Branlard G,Dardevet M.Diversity of grain proteins and bread wheat quality:ⅠCorrelation between gliadin bands and flour qualily characteristics[J].Journal of Cereal Science,1985,13:329-343
[26]高艾英,吴长艾,朱树生,等.山东省普通小麦醇溶蛋白Gli-1和Gli-2位点等位基因的遗传变异[J].作物学报,2005,31(11):1460-1465
Gao A Y,Wu C A,Zhu S S,et al.Genetic variation at Gli-1 and Gli-2 loci in wheat cultivars from Shandong province[J].Acta Agronomica Sinica,2005,31(11):1460-1465.
Genetic Diversity of Gliadin in 301 Wheat Germplasms and the Relationship with Quality Properties
Jiang Xiaoling Zhang Ziyang Li Xiaojun Li Gan Yu Hongcai Li Xiuling Ru Zhengang
(Center of Wheat Breeding,Henan Institute of Science and Technology; Collaborative Innovation Center of Modern Biological Breeding,Henan Province;Key Discipline Open Laboratory on Crop Molecular Breeding of Henan Institute,Xinxiang 453003)
In order to study the genetic diversity of gliadin and their effects on quality properties in wheat,301 wheat varieties(series)derived from different regions were used to analyze the allelic variation of gliadin by using A-PAGE.Results showed that total of 93 gliadin bands with different mobility were detected in tested materials.Moreover,five bands with the mobility of 58.6,69.4,72.1,16.5 and 19.2 had higher present frequency,greater than 80%,and the others showed higher polymorphism.The genetic similarity(GS)ranged from 0.538 to 1.000,with an average of 0.759.Furthermore,the GS value between Zhongyuan 008 and Zhongyu 1 401 was 1.0,which showed that their genetic relationship was very close.Tested materials could be divided into 11 clusters at the level of GS=0.740.The results of correlation analysis showed that significant or extremely significant correlation was found between 42 bands and 106 kinds of wheat quality parameters.For example,the bands of ω 25.5,γ 43.7 and γ 48.2 could significantly increase dough stability time and gluten index;the bands of γ 52.2 and α 80.7 could significantly increase crude protein,dry and wet gluten contents;the band of β 59.2 could significantly enhance L*value of flour color but reduce b*value.However,the bands of ω 34.8,ω 36.6 and ω 39.3 had significantly negative correlation with dough stability time and gluten index.These important gliadin bands could provide genetic makers for wheat quality breeding.
wheat,gliadin protein,genetic diversity
S512.1
A
1003-0174(2017)11-0014-07
河南省科技攻关(152102110085),河南省小麦产业技术体系岗位专家项目(S2015-01-G01)
2016-10-14
姜小苓,女,1982年出生,讲师,小麦品质改良及育种
茹振钢,男,1958年出生,教授,小麦育种
