水稻粒形遗传与长粒型优质粳稻育种进展
2017-12-02黄海祥钱前
黄海祥 钱前
(1浙江省嘉兴市农业科学研究院,浙江 嘉兴314016;2中国水稻研究所 水稻生物学国家重点实验室,杭州310006;*通讯联系人,E-mail:qianqian188@hotmail.com)
水稻粒形遗传与长粒型优质粳稻育种进展
黄海祥1钱前2,*
(1浙江省嘉兴市农业科学研究院,浙江 嘉兴314016;2中国水稻研究所 水稻生物学国家重点实验室,杭州310006;*通讯联系人,E-mail:qianqian188@hotmail.com)
水稻粒形遗传机制和粒形育种一直是水稻品质和产量研究的重要领域。在粳稻品种选育中,对长粒形遗传种质的选择,已经成为近年来一种新的育种趋势。我们在总结粒长及其相关性状的遗传研究进展和已知粒长相关基因功能的基础上,系统地介绍了南方稻区长粒粳稻的育种历程及其选育策略。同时,对以品种嘉禾218为代表的南方长粒晚粳稻系列品种农艺性状和品质特性进行了比较,在阐明长粒粳稻优质、高产的品种特点的同时,提出长粒晚粳育种中选择粒形、兼顾株型、保产提质的选育思路。
粳稻;粒长;基因;育种
我国的水稻品种属于亚洲栽培稻,它有两个亚种,即我们通常说的粳稻和籼稻。粳稻和籼稻最为显著的外观区别在于其谷粒的长度,籼稻谷粒一般较长,而粳稻谷粒则相反。近年来,由于受到国内一些著名品牌和国际市场泰国香米的影响,粳稻品种在选育过程中有长粒化的趋势[1]。通常以整精米的长宽比来衡量粒型的长短,依据国际水稻研究所的标准,长宽比大于3.0者即为细长型,长宽比小于2.0即为粗短型,长宽比在2.0~3.0则为中间型。我国优质籼米的新标准是长宽比大于 2.8。粳稻长宽比一般为1.5~2.0;长宽比大于2.5,整精米粒长≥6.5 mm者是长粒粳稻。通常,随着粒长的增加,糙米率、整精米率等加工碾磨品质会随之降低。
水稻的粒形性状与产量和品质密切相关,主要包括粒长、粒宽、粒厚、长宽比和长厚比等方面[2-3]。粒长不仅决定稻米的外观和碾磨品质,而且对水稻的千粒重也有影响[4,5],因此阐明水稻粒形的遗传与发育机理,并应用于育种实践,有助于水稻单产的提高和稻米品质的改良。水稻籽粒形成经历两个主要的生物学过程,即早期通过细胞分裂增加细胞数目,后期通过细胞膨大增加细胞的体积,胚乳细胞的数目和体积最终决定了籽粒的大小。许多学者对水稻粒型遗传进行了大量研究,现分述如下。
1 粒形遗传
1.1粒长的遗传
对粒长的遗传众说纷纭,至今仍有不同的研究表明粒长受单基因、双基因、多基因、主基因和微效基因共同控制。但是近年来,研究人员普遍认为水稻粒长以多基因控制为主[6]。Mckeniz等[7]的研究也认为粒长受多对基因共同调控。
粒长的遗传往往具有加性效应,并且在一些研究中也检测到了显性效应,但是粒长的上位效应和细胞质影响却鲜有报道,可能存在核质互作效应[8]。其实早在 1942年,赵连芳就对粒长进行了遗传分析,通过平均粒长4.13 mm的水稻品种4957与平均粒长8.81 mm的品种4269进行杂交,他发现F1的平均粒长为5.33 mm,属于中间型,且在F2粒长出现了分离(4.7~9.7 mm)。石春海等[9]利用5个细长粒品种和8个短粒品种作为材料,以短粒品种作为母本、细长粒品种作为父本,进行8×5不完全双列杂交,采用数量性状加性-显性的遗传模型对早籼谷粒性状进行了分析,发现粒长是以加性效应为主,其中近 20个组合的粒长加性效应值比率为50.6%~98.4%,认为杂交后代中选用谷粒长的单株进行育种的方法是有效的。林鸿宣等[10]利用CB1128作共同父本,分别与特三矮 2号和外引2号两个母本形成的群体(群体Ⅰ、群体Ⅱ),构建了RFLP图谱,并应用方差分析和区间作图法,在两个群体中均定位到5个控制粒长的QTL。群体Ⅰ的5个QTL中,单个QTL的贡献率比较低,总体对粒长的贡献率为49.2%;群体Ⅱ中的5个QTL对粒长的总贡献率高达62.2%,可以看出其加性效应明显大于群体Ⅰ。粒长的遗传还兼有不完全显性作用,并且一般认为显性作用在不同的品种中表现不同:长粒、中粒、短粒、极短粒显性作用依次减弱,但是也存在长粒隐性和短粒显性的现象[11]。
1.2 粒宽的遗传
研究表明,粒宽的遗传是由多基因的加性和显性效应共同控制,并且显性方向随着不同的组合而不同[12]。但也有部分研究显示某些品种的粒宽由主效基因或单基因控制[10]。此外,还有研究发现粒宽受母性影响[13]。Mckenize等[7]的研究表明粒宽由3~7个基因所控制。石春海等[14]的研究表明,在17个不同的配组中,以加性效应为主的组合占总遗传效应值达到显著或极显著水平的94.12%,表明粒宽的遗传是以加性效应为主的;但在中156与浙珍2号的杂交配组中,加性效应不显著,而是显性效应占主导地位。
1.3 谷粒长宽比的遗传
谷粒的长宽比是粒长和粒宽的复合特征,由多基因的加性或显性效应所控制[13,15]。谷粒长宽比的遗传中,加性效应和非加性效应表现均为显著,但主要为加性效应。石春海等[9]在早籼稻谷粒性状的遗传研究中发现,粒长和粒宽对谷粒长宽比均有影响,在遗传效应达显著或极显著水平的 19个杂交组合中,浙农921/湘早籼3号等18个组合以加性效应为主,其加性效应值比率达到了51.6%~99.8%;该研究还表明显性效应对加性效应有影响,显性效应负值显著,会降低杂种后代中加性效应在粒长性状上的表现。符福鸿等[16]对三系杂交稻的谷粒性状进行了遗传分析,发现F1的谷粒长宽比主要受不育系(母本)的影响,父本的影响很小。
1.4 已克隆的粒长相关基因
目前,与水稻籽粒粒长相关的基因或者 QTL被克隆的已有不少,包括SMOS1,IPA1,GS3,TGW,PGL1,PGL2,AGP,qSW5,GL7,GLW7,GS2等。水稻中SMOS1基因编码一个含AP2结构域的转录因子,通过增加细胞数目同时降低细胞大小来调控籽粒的大小[17]。SPL(SOUAMOSA PROMOTER BINDING PROTEIN-LIKE)是水稻中一类重要的转录因子,在调控粒型和株型方面发挥着重要功能。IPA1基因编码OsSPL14,该基因具有一因多效性,获得功能的突变体不但在株型上有较大改良,在粒型和千粒重等性状上也有所改观[18]。GS3是控制粒长的主效QTL,位于第3染色体的着丝粒区域[19]。该基因编码一个跨膜蛋白,由4个结构域组成:一个植物特有的器官大小调节因子(OSR)、一个跨膜蛋白、一个富含半胱氨酸的肿瘤坏死因子受体/神经生长因子受体(TNFR/NGFR)和C端的C型血管性血友病因子(VWFC)。在 OSR结构域中缺失的突变体中,籽粒会变长,可见其具有负调控粒长的功能,C端的 TNFR/NGFR结构域的功能缺失引起粒长变短,可见其具有正调节粒长的功能。该基因功能的丧失将导致籽粒变长,而它的过表达植株会出现籽粒变小、株高矮化和生长发育受限等一系列生理变化。GS3具有一因多效的作用,它不仅是一个粒形基因,同时也控制了植物生长发育进程中的一系列生理生化过程[20]。TGW6编码IAA葡萄糖水解酶,通过影响IAA的供应调控籽粒细胞数量及籽粒长度[21]。PGL1,PGL2作为粒长的正调控因子,过表达PGL1,PGL2均可使水稻粒长变长。而PGL1的负调控因子APG缺失后也会出现类似粒长变长的表型[22,23]。GL7基因编码一个与拟南芥LONGIFOLIA同源的蛋白,该蛋白调控着纵向细胞的伸长,该基因的拷贝数或等位基因越多导致GL7基因表达量上升,进而引起粒长增加[24]。GLW7编码一个高等植物特有的SPL转录因子OsSPL13,它主要通过增加细胞的大小来调控粒长[25-27]。主效QTL GS2编码水稻生长调控因子 OsGRF4,该基因表达量上升,会促进细胞分裂和生长,从而增加穗长和籽粒大小[28]。
目前已经克隆的显著影响粒宽的基因有 6个,即GW2,GW5,GS5,GW8,GS6和GW7。GW2编码E3泛素连接酶,通过蛋白酶体的降解作用负调控水稻籽粒的宽度和粒重[29]。GW5编码一个小的核定位蛋白,与糖原合成激酶GSK2直接互作,抑制其激酶活性,通过参与BR信号调节粒宽和粒重[30,31]。GW8/OsSPL16属于SBP型转录因子,高表达GW8可以促进细胞分裂进而增加粒宽和水稻产量。进一步研究发现GW8与GW7(编码TONNEAU1招募基序蛋白)启动子直接互作并抑制GW7的表达,阻止籽粒纵向伸长,增加籽粒的宽度[32,33]。GS6编码一个GRAS家族蛋白。GS6在ATG后348 bp的翻译提前终止可以显著增加水稻的粒宽和粒重,而且提前终止的等位基因在粳稻中被强烈选择,90个不同的水稻品种中,95%的粳稻GS6翻译提前终止,产生宽粒表型[34]。qSW5与GW2的功能类似,通过增加水稻籽粒的外颖壳的细胞数目使籽粒变宽[35]。qSW5在水稻的第5染色体上,编码一个新的富含精氨酸结构域的蛋白,直接与多聚泛素互相作用,定位在细胞核中。在第5染色体上还有另一个控制水稻粒宽的基因GS5,与qSW5相邻,作为一个粒宽正调控因子,水稻粒宽随着GS5表达量的升高而变得更宽,GS5编码丝氨酸羧肽酶,其对籽粒的灌浆和粒重也有一定的影响[36]。
2 粳稻长粒型育种改良与应用
一般来讲,粳稻的蒸煮食味品质要优于籼稻,而在粒型上,籼稻比粳稻要长,外观品质更佳,市场竞争力强,但谷粒过长,其糙米率、精米率和整精米率又会下降,而且谷粒长与谷粒宽、谷粒厚负相关[37]。因此,在品种培育与改良过程中,如何协调粳稻粒型与品质间的矛盾,即在粳稻消费区保证粳稻蒸煮食味品质特征的同时,增长其粒型性状,减少垩白率进而提高其外观品质,同时提升加工碾磨品质,确实是水稻育种上一个重要课题。
近年来,嘉兴市农业科学研究院、中国水稻研究所等单位合作选育的嘉禾系列品种较好地解决了这个问题。嘉禾系列的品种属于粳型,蒸煮食味品质优异,同时其外观品质基本也达到了优质籼稻的标准。嘉禾系列品种中最为突出的是嘉禾218和嘉禾 212,其遗传背景来源于热带粳稻,经过与温带粳稻不断回交所得。其粒长约7.0 mm,长宽比高达3.0,整精米率接近60%。以籼稻一级稻米6.5 mm和2.8的粒长和长宽比为标准,嘉禾218的这两个性状显著优于籼稻一级稻米。对嘉禾212和嘉禾218的长粒形基因定位结果显示,在其GS3基因上发生了与目前中国大面积生产的长粒籼型一样的突变,突变位点一致,且都是C-A突变(图1)
在品质方面,嘉禾218的米质也达到部颁粳稻一级,米粒晶莹透明,商品性出众,蒸煮米饭食味优良,松软适口。值得一提的是,嘉禾218解决了粒长与整精米率(加工碾磨品质)的矛盾,在保证粒长的同时,整精米率也没有明显下降。而且在透明度、垩白率、垩白度等外观品质方面,胶稠度、直链淀粉含量、蛋白质含量等蒸煮品质方面,都达到了部颁粳稻一级或二级(表1)。因此,被誉为粳稻中的“泰国香米”,产品屡获浙江省农业博览会金奖、中国优质稻米博览会金奖。尤其是 2011年,首次参赛的清溪牌清溪香米,以总分第三,荣获第十届中国优质稻米博览会金奖,进入国内高端大米前列,提升了浙江产地大米在高端市场的竞争力。在浙江省、上海市、安徽省等“长三角”地带,推广面积逐年增大,也产生了较好的社会经济效益。
嘉禾系列品种不但在品质上极为出色,在产量上也有不俗的表现,嘉禾218在2013-2017年度列入浙江省晚稻主导品种。在用嘉禾218同时作为母本的两个杂交组合中,遗传背景一致的情况下,同时选育长粒(嘉禾291)和圆粒(嘉禾288)的品系,对两个不同的品系各方面的参数进行检测后发现,长粒的后代株系,不仅在长宽比等多个品质参数上取得优势(表1),而且在产量上也表现出色(表2)。
3 总结与展望
未来的粳稻育种需要着重改良粒型,充分利用已克隆的粒长相关基因,展开分子辅助育种。通过检测,嘉禾系列的品种中均带有gs3、dep1等影响粒型的基因[38],所以在亲本选育中,要充分考虑选用特定粒型的不育系配组,同时兼顾株型,以保证其丰产性。从嘉禾218的田间长势来看,其叶片偏籼型,分蘖角度、粒型等也偏籼型(图1),嘉禾218品种开创了长粒晚粳育种种质创新的新领域。

图1 长粒粳稻不育系嘉禾212A GS3基因突变位点鉴定Fig. 1. GS3 gene mutational site identification in CMS-Jiahe 212.
种质资源创新、优异基因聚合是今后水稻育种的总体趋势,如嘉丰优2号(浙审稻2017012),其不育系为带有gs3、dep1的长粒晚粳嘉禾212A,恢复系带有理想株型 IPA1-2。2015年试验平均产量为659.4 kg/667 m2,比对照两优培九增产14.7%,达极显著水平;2016年试验平均产量为687.5 kg/667 m2,比对照两优培九增产18.7%,达极显著水平。两年省区试平均产量为673.5 kg/667 m2,比对照两优培九增产16.7%。两年平均全生育期144.2 d,比对照两优培九长7.0 d。该组合有效穗数为11.5万/667 m2,株高126.9 cm,每穗总粒数273.3粒,实粒数215.5粒,结实率78.8%,千粒重25.7 g。经浙江省农业科学院植物保护与微生物研究所 2015-2016年抗性鉴定,平均叶瘟3.2级,穗瘟1.5级,穗瘟损失率1级,综合指数为2.1;白叶枯病5.4级;褐稻虱8级。经中华人民共和国农业部稻米及制品质量监督检测中心 2015-2016年检测,平均整精米率64.1%,长宽比 2.7,垩白粒率 9%,垩白度 0.8%,透明度2级,胶稠度78 mm,直链淀粉含量15.1%,米质各项指标达到中华人民共和国农业部优质粳稻谷二级标准。

表1 部分嘉禾系列品种的品质性状Table 1. Quality characters of some Jiahe series cultivars.
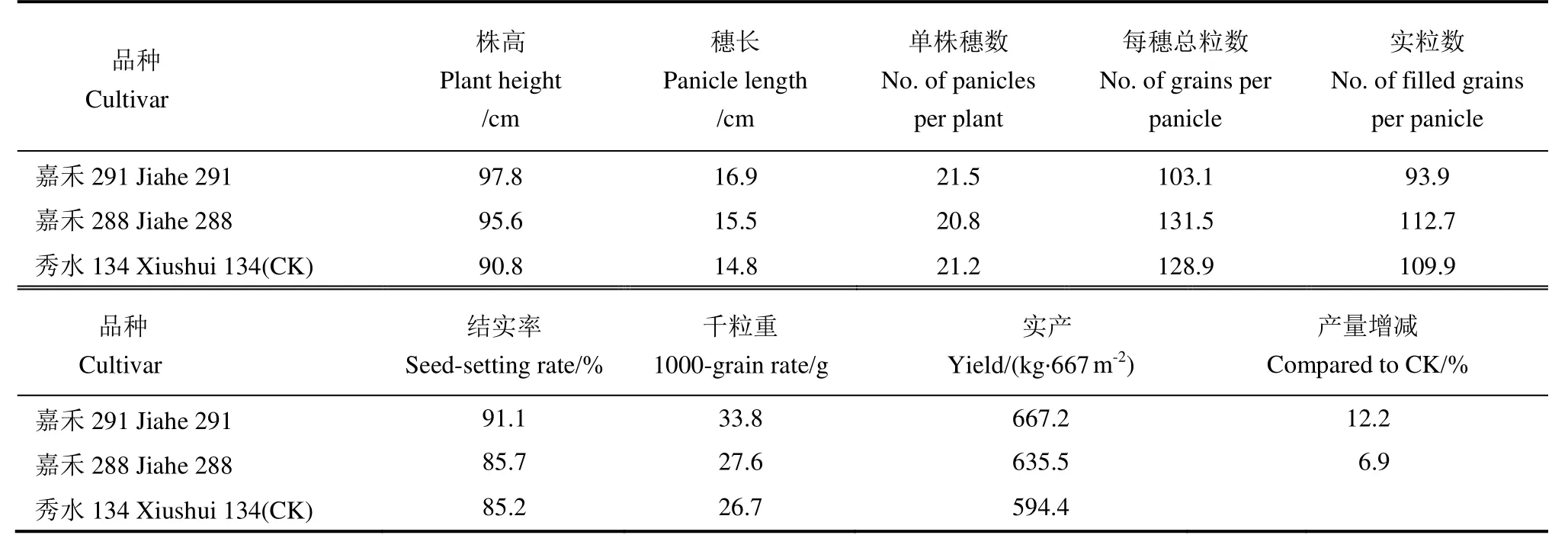
表2 2016年常规晚稻品比试验产量及经济性状Table 2. Quality and economical characters of Jiahe 291 and Jiahe 288 compared with conventional late rice in 2016.
另外,我们利用ZH559/嘉禾218新育成长粒晚粳品种中嘉8号(浙审稻2017008),2014年浙江省单季晚粳稻区域试验平均产量为632.3 kg/667 m2,比对照秀水134增产4.7%;2015年试验平均产量为637.7 kg/667 m2,列区试组产量第1位,比对照秀水134增产7.6%,达极显著水平。两年省区试平均产量为 635.0 kg/667 m2,比对照秀水 134增产6.1%。该组合作单晚种植株高105.0 cm,两年平均全生育期162.1 d,比对照秀水134长0.9 d。有效穗数为23.0万/667 m2,株高106.7 cm,每穗总粒数143.4,每穗实粒数 123.7,结实率 86.3%,千粒重29.7 g。经中华人民共和国农业部稻米及制品质量监督检测中心2014年检测,平均整精米率为64.0%,长宽比2.4,垩白粒率26%,垩白度3.8%,透明度1级,胶稠度72 mm,直链淀粉含量16.0%。中抗稻瘟病、褐稻虱,抗条纹叶枯病和抗稻曲病。
从嘉禾218到中嘉8号,在长粒粳稻育种过程中,我们实践着选择粒形、兼顾株型、保产提质的选育思路,培育出一系列品质与产量协同提高的水稻新品种。最初我们以半矮生型晚粳稻为受体,与热带长粒粳稻杂交,经适当回交,在后代中选择细韧秆稻株,保持其多蘖优势;筛选供体亲本中长穗特性,为保持适宜的着粒密度提供保障;在米质优良的基础上,选择较大千粒重且粒形长的品系,这样有利于产量的提升和品质保证。长粒粳稻品种不仅作为常规晚粳稻,实现了优质与产量的协同提升,转育为不育系后,也同样在杂交水稻组合中,成功培育了能兼顾高产、优质的新品种(如嘉丰优2号)。
在未来的长粒型粳稻育种中,我们不仅可以充分利用GS3、GL7等控制粒长的基因,同时还可以结合分子辅助手段,聚合多个产量优势基因的后代品系,培育更多高产优质的水稻新品种。我们的育种实践也充分证明,改良籽粒的长度后,在保证产量的同时,品质也可以得到提升。在以后的粳稻育种中,需要发掘更多的优异基因资源,改良粳稻穗型、叶角等涉及受光姿态的性状,最终将籼、粳各自良好的性状完美结合。
[1] 胡子诚,贾锦娟. 长粒优质高产水稻新品种宁粳36的选育及其栽培要点. 中国稻米,2007(1): 26.Hu Z C,Jia J J. Breeding and cultivation method of Ningjing 36,a long-grain rice cultivar with good quality.China Rice,2007(1): 26. (in Chinese)
[2] 徐建龙,薛庆中,罗利军,黎志康. 水稻粒重及其相关性状的遗传解析. 中国水稻科学,2002,16(1): 6-10.Xu J L,Xue Q Z,Luo L J,Li Z K. Genetic dissection of grain weight and its related traits in rice (Oryza sativa L.).Chin J Rice Sci,2002,16(1): 6-10. (in Chinese with English abstract)
[3] 高志强,占小登,梁永书,程式华,曹立勇. 水稻粒形性状的遗传及相关基因定位与克隆研究进展. 遗传,2011,33(4): 314-321.Gao Z Q,Zhan X D,Liang Y S,Cheng S H,Cao L Y.Progress on genetics of rice grain shape trait and its related gene mapping and cloning. Hereditas(Beijing),2011,33(4): 314-321. (in Chinese with English abstract)
[4] 罗玉坤,朱智伟,陈能,段彬伍,章林平. 中国主要稻米的粒型及其品质特性. 中国水稻科学,2004,18(2):135-139.Luo Y K,Zhu Z W,Chen N,Duan B W,Zhang L P.Grain types and related quality characteristics of rice in China. Chin J Rice Sci,2004,18(2): 135-139. (in Chinese with English abstract)
[5] 杨梯丰,曾瑞珍,朱海涛,陈岚,张泽民,丁效华,李文涛,张桂权. 水稻粒长基因GS3在聚合育种中的效应.分子植物育种,2010,8(1): 59-66.Yang T F,Zeng R Z,Zhu H T,Chen L,Zhang Z M,Ding X H,Li W T,Zhang G Q. Effect of grain length gene GS3 in pyramiding breeding of rice. Mol Plant Breed,2010,8(1): 59-66. (in Chinese with English abstract)
[6] 姚国新,卢磊. 水稻粒重基因定位克隆研究. 安徽农业科学,2007,35(27): 8468-8478.Yao G X,Lu L. Research on the localization and clone of grain weight genes in rice. J Anhui Agric Sci,2007,35(27): 8468-8478. (in Chinese with English abstract)
[7] Mckenize K S,Rutger J N. Genetic analysis of amylase content,alkali spreading score and grain dimensions in rice. Crop Sci,1983,23(2): 306-313.
[8] 芮重庆,赵安常. 籼稻粒重及粒型性状F1遗传特性双列分析. 中国农业科学,1983,16(5): 14-20.Rui C Q,Zhao C A. Genetic analysis of weight and shape of F1’s grains by diallel crossing method in hsien rice. Sci Agric Sin,1983,16(5): 14-20. (in Chinese with English abstract)
[9] 石春海,申宗坦. 早籼稻谷粒性状遗传效应分析. 浙江农业大学学报,1994,20(4): 405-410.Shi C H,Shen Z T. Analysis of genetic effects of grain traits in indica rice. J Zhejiang Agric Univ,1994,20(4):405-410. (in Chinese with English abstract)
[10] 林鸿宣,闵绍措,熊振民,钱惠荣,庄杰云,陆军,郑康乐. 应用RFLP图谱定位分析籼稻粒形数量性状基因座位. 中国农业科学,1995,28(4): 1-7.Lin H X,Min S K,Xiong Z M,Qian H R,Zhuang J Y,Lu J,Zheng K L. RFLP mapping of QTLs for grain shape traits in indica rice (Oryza sativa L. subsp indica). Sci Agric Sin,1995,28(4): 1-7. (in Chinese with English abstract)
[11] 莫惠栋. 我国稻米品质的改良. 中国农业科学,1993,26(4): 8-14.Mo H D. Quality improvement of rice grain in china. Sci Agric Sin,1993,26(4): 8-14. (in Chinese with English abstract)
[12] 刘晓辉. 谷子千粒重遗传的双列分析. 吉林农业科学,1989(4): 33-35.Liu X H. Diallel genetic analysis of millet 1000 grain weight. J Jilin Agric Sci,1989(4): 33-35. (in Chinese)
[13] 石春海,申宗坦. 籼稻粒形及产量性状的加性相关和显性相关分析. 作物学报,1996,22(1): 36-42.Shi C H,Shen Z T. Additive and dominance correlation analysis of grain shape and yield traits in indica rice. Acta Agron Sin,1996,22(1): 36-42. (in Chinese with English abstract)
[14] 石春海,申宗坦. 早籼粒形的遗传和改良. 中国水稻科学,1995,9(1): 27-32.Shi C H,Shen Z T. Inheritance and improvement of grain shape in indica rice. Chin J Rice Sci,1995,9(1): 27-32.(in Chinese with English abstract)
[15] 周清元,安华,张毅,沈福成. 水稻子粒形态性状遗传研究. 西南农业大学学报,2000,22(2): 102-104.Zhou Y Q,An H,Zhang Y,Shen F C. Study on heredity of morphological character of rice grain. J Southwest Agric Univ,2000,22(2): 102-104. (in Chinese with English abstract)
[16] 符福鸿,王丰. 杂交水稻谷粒性状的遗传分析. 作物学报,1994,20(1): 39-45.Fu F H,Wang F. Genetic analysis on grain characters in hybrid rice. Acta Agron Sin,1994,20(1): 39-45. (in Chinese with English abstract)
[17] Aya K,Hobo T,Sato-Izawa K,Ueguchi-Tanaka M,Kitano H,Matsuoka M. A novel AP2-type transcription factor,SMALL ORGAN SIZE1,controls organ size downstream of an auxin signaling pathway. Plant amp; Cell Physiol,2014,55(5): 897-912.
[18] Jiao Y,Wang Y,Xue D,Wang J,Yan M,Liu G,Dong G,Zeng D,Lu Z,Zhu X,Qian Q,Li J. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat Genet,2010,42(6): 541-544.
[19] Fan C,Xing Y,Mao H,Lu T,Han B,Xu C,Li X,Zhang Q. GS3,a major QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein. Theor Appl Genet,2006,112(6): 1164-1171.
[20] Mao H,Sun S,Yao J,Wang C,Yu S,Xu C,Li X,Zhang Q. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci USA,2010,107(45): 19579-19584.
[21] Ishimaru K,Hirotsu N,Madoka Y,Murakami N,Hara N,Onodera H,Kashiwagi T,Ujiie K,Shimizu B,Onishi A,Miyagawa H,Katoh E. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet,2013,45(6):707-711.
[22] Heang D,Sassa H. An atypical bHLH protein encoded by POSITIVE REGULATOR OF GRAIN LENGTH 2 is involved in controlling grain length and weight of rice through interaction with a typical bHLH protein APG.Breed Sci,2012,62(2): 133-141.
[23] Heang D,Sassa H. Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice.PLoS One,2012,7(2): e31325.
[24] Wang Y,Xiong G,Hu J,Jiang L,Yu H,Xu J,Fang Y,Zeng L,Xu E,Xu J,Ye W,Meng X,Liu R,Chen H,Jing Y,Wang Y,Zhu X,Li J,Qian Q. Copy number variation at the GL7 locus contributes to grain size diversity in rice.Nat Genet,2015,47(8): 944-948.
[25] Si L,Chen J,Huang X,Gong H,Luo J,Hou Q,Zhou T,Lu T,Zhu J,Shang guan Y,Chen E,Gong C,Zhao Q,Jing Y,Zhao Y,Li Y,Cui L,Fan D,Lu Y,Weng Q,Wang Y,Zhan Q,Liu K,Wei X,An K,An G,Han B.OsSPL13 controls grain size in cultivated rice. Nat Genet,2016,48(4): 447-456.
[26] Qi P,Lin Y S,Song X J,Shen J B,Huang W,Shan J X,Zhu M Z,Jiang L,Gao J P,Lin H X. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res,2012,22(12):1666-1680.
[27] Che R,Tong H,Shi B,Liu Y,Fang S,Liu D,Xiao Y,Hu B,Liu L,Wang H,Zhao M,Chu C. Control of grain size and rice yield by GL2-mediated brassinosteroid responses.Nat Plants,2015,21(2): 15195.
[28] Hu J,Wang Y,Fang Y,Zeng L,Xu J,Yu H,Shi Z,Pan J,Zhang D,Kang S,Zhu L,Dong G,Guo L,Zeng D,Zhang G,Xie L,Xiong G,Li J,Qian Q. A rare allele of GS2 enhances grain size and grain yield in rice. Mol Plant,2015,8(10): 1455-1465.
[29] Song X J,Huang W,Shi M,Zhu M Z,Lin H X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet,2007,39(5): 623-630.
[30] Weng J,Gu S,Wan X,Gao H,Guo T,Su N,Lei C,Zhang X,Cheng Z,Guo X,Wang J,Jiang L,Zhai H,Wan J. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight.Cell Res,2008,18(12): 1199-1209.
[31] Liu J,Chen J,Zheng X,Wu F,Lin Q,Heng Y,Tian P,Cheng Z,Yu X,Zhou K,Zhang X,Guo X,Wang J,Wang H,Wan J. GW5 acts in the brassinosteroid signaling pathway to regulate grain width and weight in rice. Nat Plants,2017,3: 17043.
[32] Wang S,Wu K,Yuan Q,Liu X,Liu Z,Lin X,Zeng R,Zhu H,Dong G,Qian Q,Zhang G,Fu X. Control of grain size,shape and quality by OsSPL16 in rice. Nat Genet,2012,44(8): 950-954.
[33] Wang S,Li S,Liu Q,Wu K,Zhang J,Wang S,Wang Y,Chen X,Zhang Y,Gao C,Wang F,Huang H,Fu X. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat Genet,2015,47(8): 949-954.
[34] Sun L,Li X,Fu Y,Zhu Z,Tan L,Liu F,Sun X,Sun X,Sun C. GS6,a member of the GRAS gene family,negatively regulates grain size in rice. J Integr Plant Biol,2013,55(10): 938-949.
[35] Shomura A,Izawa T,Ebana K,Ebitani T,Kanegae H,Konishi S,Yano M. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet,2008,40(8): 1023-1028.
[36] Li Y,Fan C,Xing Y,Jiang Y,Luo L,Sun L,Shao D,Xu C,Li X,Xiao J,He Y,Zhang Q. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet,2011,43(12): 1266-1269.
[37] 徐正进,陈温福,马殿荣,吕英娜,周淑清,刘丽霞.稻谷粒形与稻米主要品质性状的关系. 作物学报,2004,30(9): 894-900.Xu Z J,Chen W F,Ma D R,Lv Y N,Zhou S Q,Liu L X.Correlations between rice grain shapes and main qualitative characteristics. Acta Agron Sin,2004,30(9):894-900. (in Chinese with English abstract)
[38] Huang X,Qian Q,Liu Z,Sun H,He S,Luo D,Xia G,Chu C,Li J,Fu X. Natural variation at the DEP1 locus enhances grain yield in rice. Nat Genet,2009,41(4):494-497.
Progress in Genetic Research of Rice Grain Shape and Breeding Achievements of Long-grain Shape and Good Quality japonica Rice
HUANG Haixiang1,QIAN Qian2,*
(1Jiaxing Academy of Agricultural Sciences,Jiaxing 314016,China; 2State Key Laboratory of Rice Biology,China National Rice Research Institute,Hangzhou 310006,China; *Corresponding author,E-mail: qianqian188@hotmail.com)
The genetic dissection and breeding of rice grain shape has always been an important subject in rice genetics.The long-grain shape has been a new favored quality trait in the breeding of japonica rice in recent years. While summarizing the genetic research progress in grain length-related traits and the corresponding gene function,we also systematically introduced the breeding course and selection strategy of long-grain japonica rice. Meanwhile,the agronomic and quality traits of long grain japonica varieties,represented by Jiahe 218,were compared with those of typical japonica rice varieties. Based on the successful breeding of long-grain japonica rice with high quality and high yield,we put forward the breeding strategy that combines grain shape selection,plant architecture optimization with yield and quality.
japonica; grain length; gene; breeding
S511.032; S511.2+2
A
1001-7216(2017)06-0665-08
10.16819/j.1001-7216.2017.7115
2017-05-20; 修改稿收到日期:2017-06-25。
国家自然科学基金资助项目(91535205,31521064)。
