不同国槐根茎腐烂病镰刀型致病菌的同工酶分析①
2017-11-22王桂清
马 迪 王桂清 张 赛
(聊城大学 农学院,山东 聊城 252059)
不同国槐根茎腐烂病镰刀型致病菌的同工酶分析①
马 迪 王桂清 张 赛
(聊城大学 农学院,山东 聊城 252059)
利用聚丙烯酰胺凝胶电泳技术,对分离于聊城地区国槐根茎腐烂病的24个镰刀菌菌株进行了可溶性蛋白质和同工酶电泳图谱分析及聚类分析,明确了镰刀菌生理分化的物质基础.研究表明,镰刀菌在可溶性蛋白质和EST、SOD、PPO、POD等同工酶存在差异,不同菌株之间某些同工酶谱带数和同一迁移率谱带的亮度和色泽差异非常显著,说明镰刀菌不同种或同种不同菌株间的多态性可在同工酶水平上得到反映.研究还发现,同工酶的变化与镰刀菌的种类以及侵染部位密切相关.
镰刀菌,可溶性蛋白质,同工酶,聚类分析
传统意义上,真菌主要根据菌落及繁殖器官的形态特征进行分类.镰孢菌由于生境复杂,其用于分类的许多形态特征指标容易受培养基营养条件、光质光强、温湿度等环境条件影响,产生变异,从而导致形态分类较困难.同工酶研究即是在种和亚种水平上对真菌进行鉴定的一种有效工具,也是对不同真菌进行种间或种内分析的一种有用手段[1],技术成熟,应用广泛.本研究采用同工酶/可溶性蛋白质凝胶电泳技术,对来源于聊城地区引起国槐根茎腐烂病的不同镰刀菌菌株进行分析,从蛋白质和酶学水平上的多态性分析镰刀菌生理分化的物质基础特征,为研究其生理生化特性、致病性分化等奠定了基础.
1 材料与方法
1.1 供试菌株
供试病原菌为引起国槐根茎腐烂病的镰刀菌,来源于山东省聊城市不同街区、不同发病部位,经单胞分离纯化,共24个菌株.于PDA培养基25±1 ℃恒温条件培养5 d,备用.
1.2 酶蛋白的制备与提取
自菌落边缘,用直径7 mm的打孔器打取菌片,接种于PA培养液中,每瓶15个菌片,于25±1 ℃120 rpm条件下振荡培养6 d,抽滤获取菌丝,蒸馏水冲洗3次,吸干水分备用.按王桂清等方法[2]提取酶蛋白, 1 g菌丝,加入5.0 mL0.l mol·L-1磷酸缓冲液(pH 7.0),冰浴研磨,4 ℃2 000 rpm条件下离心20 min,上清液即为酶蛋白,置于-20 ℃冰箱备用.
1.3 蛋白质及同工酶电泳凝胶染色
参照胡能书[3]方法制胶及非变性PAGE电泳.其中分离胶浓度7.5%,胶内电压90 V;浓缩胶浓度3%,胶内电压80 V.
可溶性蛋白质(Soluble protein)、酯酶(EST)、超氧化物歧化酶(SOD)、过氧化物酶(POD)和多酚氧化酶(PPO)的染色分别参照李成文[4]、赵玉锦[5]、罗广华[6]、褚西宁[7]和袁凤杰[8]的方法.
1.4 电泳图谱的聚类分析
测量凝胶图谱中各谱带的泳动距离,计算Rf 值.将电泳后获得的每一条谱带的有无作为统计参数,有酶带记为“1”,无酶带记为“0”,从而得到电泳的“0、1”矩阵,运用NTSYS2.1软件进行数据分析,生成24个镰刀菌菌株可溶性蛋白和同工酶电泳聚类分析树状图.
Rf 值的计算公式为

2 结果与分析
2.1 病菌可溶性蛋白质和同工酶电泳图谱分析
2.1.1 谱带数量分析.引起国槐根茎腐烂病的24个镰刀菌菌株的可溶性蛋白和同工酶电泳图谱如图1所示.不同的镰刀菌菌株在可溶性蛋白质和同工酶电泳图谱的谱带数量上存在一定的差异:
可溶性蛋白质(Soluble protein)标记比较丰富,不同来源的24个镰刀菌菌株共分离出了32条谱带,其中供试菌株G1的谱带最多,为24条,供试菌株J10-4的谱带最少,为12条,谱带在凝胶的上、中、下各部都有分布.共同谱带有5条,Rf值分别为0.027,0.065,0.087,0.130和0.913,占所有谱带的15.6%,特异性谱带有27条,占所有谱带的84.4%,说明,24个供试菌株在可溶性蛋白质水平上差异较大,生理分化明显.
酯酶(EST)电泳图谱也是标记最丰富的同工酶图谱之一,供试菌株共分离出21条颜色较深且清晰可见的谱带,其中共同谱带3条,Rf值分别为0.096,0.277和0.511,占所有谱带的14.3%,特异性谱带18条,占所有谱带的85.7%.同时实验结果表明不同菌株谱带数量差异较大,供试菌株J10-4和G1的谱带最多,为11条,菌株J7-7、J7-2和J10-5的谱带最少,为5条,谱带在凝胶的上、中、下各部都有分布.
超氧化物歧化酶(SOD)同工酶的聚丙烯酰氨凝胶电泳图谱(图1)显示,供试菌株SOD同工酶电泳产生的条带数远少于可溶性蛋白和EST,不同来源的24个镰刀菌菌株共分离出了10条清晰透明的SOD谱带,其中供试菌株J7-3的谱带最多,为8条,供试菌株J7-2的谱带最少,为2条,谱带主要分布在凝胶的中部.共同谱带2条,Rf值分别为0.333和0.479,占所有谱带的20%;特异性谱带8条,占比80%.多数菌株的谱带数在3-5条之间,有一定的相似性.
病菌多酚氧化酶(PPO)电泳结果(图1)显示,供试菌株共分离出11条PPO谱带,无共同谱带,均为特异性谱带.其中供试菌株J7-3的谱带最多,为6条,菌株G4-3没有谱带,菌株G4-2、G5-3、G5-4、J9-4、J10-3和J10-4都只有1条PPO谱带,主要分布在凝胶的中上部.
过氧化物酶(POD)同工酶谱带数很少,仅4条,且没有共同谱带,均为特异性谱带.其中供试菌株J10-4的谱带最多,为2条(Rf1=0.056,Rf3=0.222);菌株G3在Rf2=0.167处、G6在Rf3=0.222处、G5-2在Rf4=0.264处和G4-1、G4-2、G5-4、J7-3、J9-2、J9-3、J9-4和J10-5在Rf1=0.056处各有1条POD谱带;其余12个菌株没有谱带,主要分布在凝胶上部.
2.1.2 谱带颜色分析.从电泳图谱(图1)可以看出,不同镰刀菌菌株的可溶性蛋白和同工酶谱带不仅在数量上有差异,而且相同谱带的颜色和宽度也存在差异,说明酶带活性不同,酶带越宽、颜色越深,酶带活性越高.如菌株G5-4和J7-1.两者在可溶性蛋白质(Soluble protein)图谱中Rf8=0.065处和在酯酶(EST)图谱中Rf5=0.096处的谱带颜色明显不同,J7-1的明显比G5-4的深,即该处菌株J7-1的酶带活性比G5-4的高;再如SOD酶,24个菌株在Rf9=0.479处的共同谱带颜色和宽度明显不同,菌株J7-7、G5-1和G5-2的谱带明显比其它菌株的深且宽,即这些菌株在该处的酶带活性高.
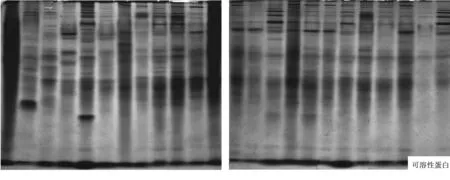
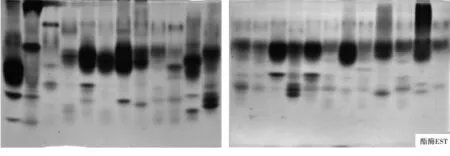
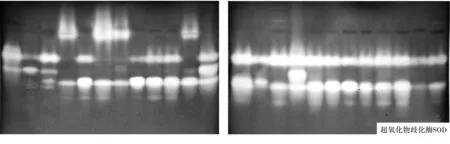
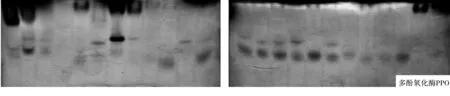
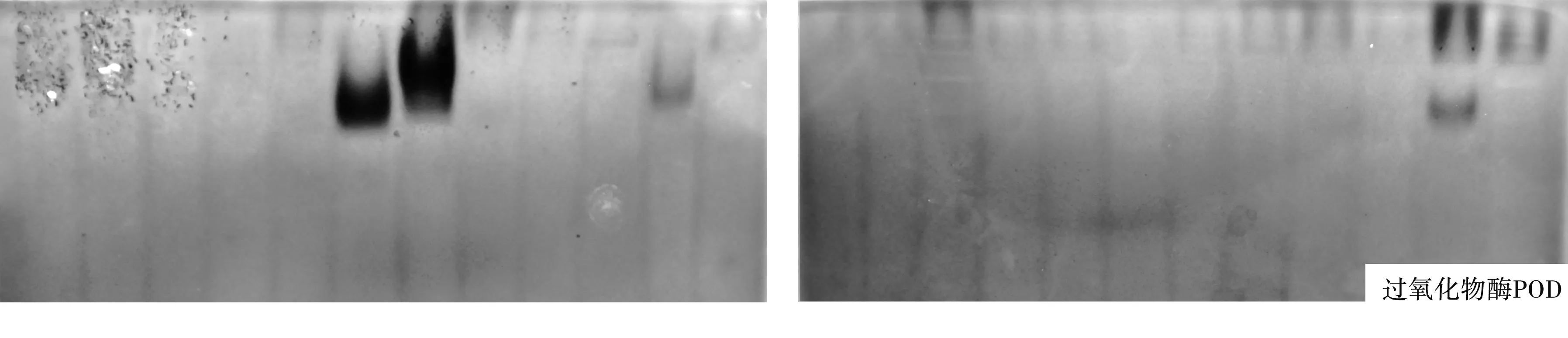
图1 不同镰刀菌可溶性蛋白质和同工酶电泳图谱注:图谱从左到右依次为供试菌株G1、G2、J7-7、G4-3、G5-3、G6、G3、G4-1、G4-2、G5-1、G5-2、G5-4、J7-1、J7-2、J7-3、J7-5、J8、J9-1、J9-2、J9-3、J9-4、J10-3、J10-4、J10-5.
2.2 病菌同工酶水平上的聚类分析
根据可溶性蛋白及EST、SOD、POD和PPO 4种同工酶谱带,运用NTSYS软件得出聚类树状图(图2).从图中可以看出供试菌株的相似系数在0.60-0.93之间,说明24个镰刀菌菌株可溶性蛋白及4种同工酶谱带存在多样性.以相似系数67%为阈值,可将供试菌株分为5个组(如表1),第Ⅰ组只包括菌株G1;第Ⅱ组包括菌株G2和J7-7;第Ⅲ组包括菌株G4-3和G6;第Ⅳ组比较大,包括G5-3、G4-2、G5-1、G5-2、G5-4、G3和G4-1共7个菌株,这7个菌株又可以相似系数72.5%为阈值分为3个亚组,其中菌株G5-3、G4-2和G5-1为一亚组,菌株G5-2和G5-4为一亚组,菌株G3和G4-1为一亚组;第Ⅴ组最大,包括12个菌株,即J7-1、J7-2、J7-5、J8-1、J9-1、J9-2、J10-3、J9-3、J9-4、J7-3、J10-5和J10-4,12个菌株又可根据相似系数83.5%为阈值分为4个亚组,其中菌株7-1和7-2为一个亚组,菌株7-5、8-1、9-1、9-2、10-3、9-3和9-4为一个亚组,菌株7-3和10-5为一个亚组,菌株10-4为一个亚组.
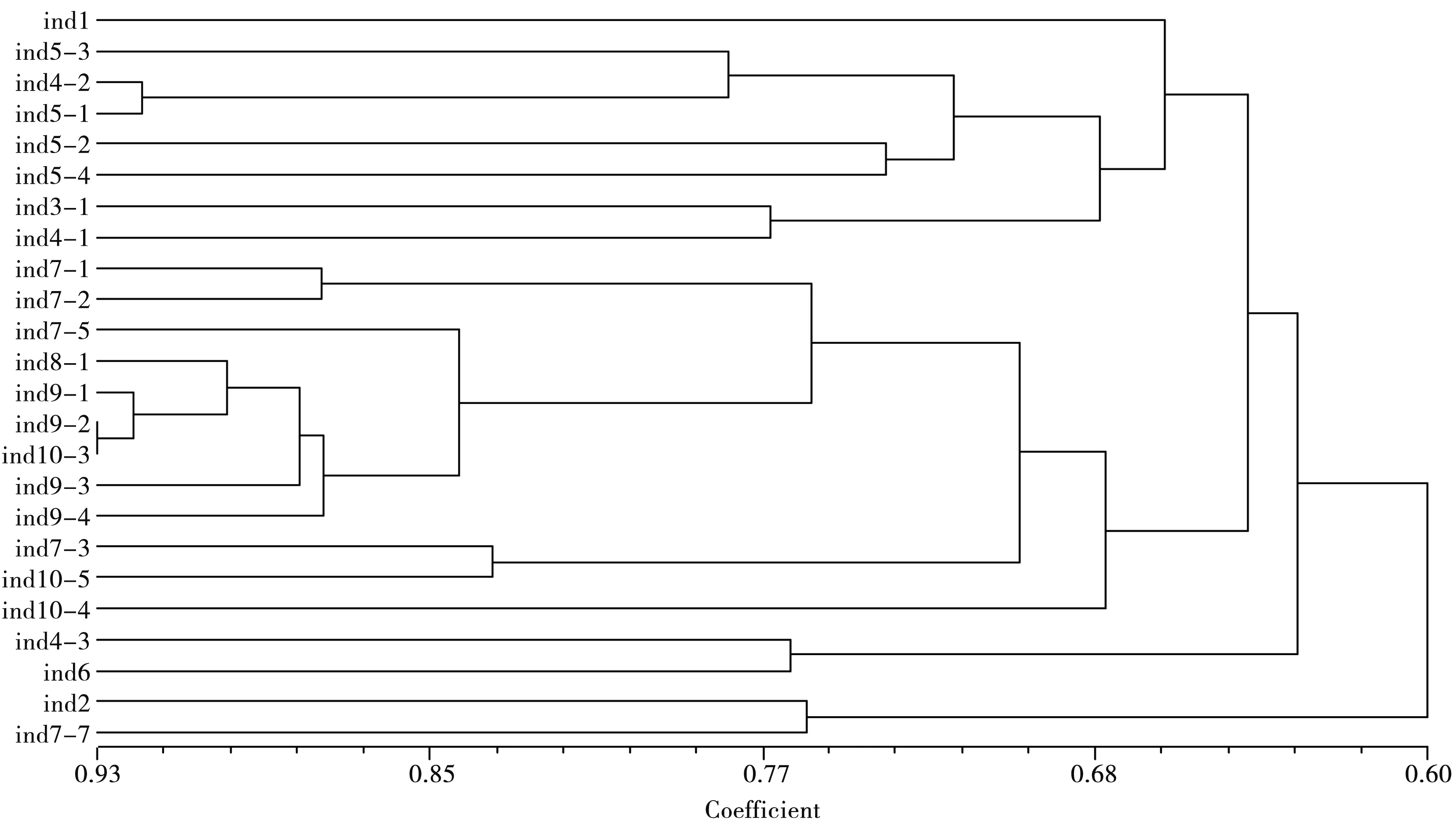
图2 不同镰刀菌菌株可溶性蛋白及4种同工酶水平上的聚类分析
形态学和分子生物学鉴定结果[9]表明,第Ⅳ组和第Ⅴ组的19个菌株均为腐皮镰刀菌(F.solani),而在同工酶分组中却分属两组,第Ⅳ组中的7个菌株均分离于国槐根部(菌株号以G开头),而第Ⅴ组中的12个菌株则分离于国槐茎部(菌株号以J开头).说明可溶性蛋白和同工酶聚类分组既与病原菌种类有关,又与同种病菌不同菌株的分离部位密切相关.
表1不同镰刀菌菌株的分组结果
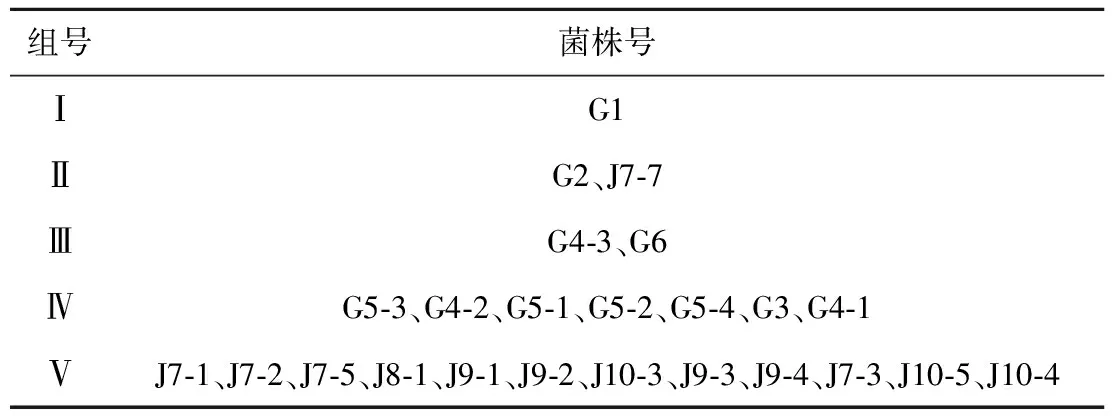
组号菌株号ⅠG1ⅡG2、J7⁃7ⅢG4⁃3、G6ⅣG5⁃3、G4⁃2、G5⁃1、G5⁃2、G5⁃4、G3、G4⁃1ⅤJ7⁃1、J7⁃2、J7⁃5、J8⁃1、J9⁃1、J9⁃2、J10⁃3、J9⁃3、J9⁃4、J7⁃3、J10⁃5、J10⁃4
3 结论与讨论
3.1 国槐根茎腐烂病镰刀型致病菌在同工酶水平上存在多态性
同工酶作为一类蛋白质,普遍存在于生物的同一种属或同一个体的不同组织,甚至同一组织、同一细胞中,同工酶的研究已经成为形态遗传学的重要内容.本试验通过对国槐根茎腐烂病镰刀型致病菌可溶性蛋及EST、SOD、POD和PPO的PAGE电泳,结果表明,供试菌株产生了大量的条带,表现出不同程度的多态性;电泳谱带相似系数为62%以上,说明可溶性蛋白和同工酶图谱具有属、种的特异性;供试菌株不仅在可溶性蛋白质和同工酶的谱带数量上存在差异,而且同一迁移率的谱带在色泽深度和亮度上也存在明显不同,说明供试菌株在酶活性方面也存在分化性或多态性,该研究与Huss等学者[10~12]的研究结果相一致.纹枯病菌、核盘菌、辣椒疫霉菌等在可溶性蛋白质和同工酶水平上也存在多态性[13-15],说明可溶性蛋白质和同工酶已在多种病原菌生理分化中作为辅助鉴别的生化标记.
3.2 酶学聚类分析结果与镰刀菌的种类和侵染部位密切关系
根据可溶性蛋白及EST、SOD、POD和PPO四种同工酶进行电泳,聚类分析结果表明,不同的镰刀菌种类分属于不同的组;19个腐皮镰刀菌菌株分属于两个不同的组,第Ⅳ组中的7个腐皮镰刀菌菌株均分离于国槐根部(菌株号以G开头),而第Ⅴ组中的12个腐皮镰刀菌菌株则分离于国槐茎部(菌株号以J开头),说明酶学聚类分组与同种病菌不同菌株的分离部位有密切的关系.镰刀菌种类和侵染国槐不同部位的腐皮镰刀菌在酶学性质方面存在差异,这种差异可能会导致致病性或致病机理方面的差异.陈伟群[16]研究表明,可溶性蛋白质凝胶电泳技术可以明确地将不同链格孢菌属Alternaria菌株鉴定到种级水平.邹庆道[17]、车建美等[18]研究表明,同一寄主来源的禾谷镰刀菌和尖孢镰刀菌的基因表达均存在一定的差异,存在分离部位分化现象.作为基因表达的产物,酶能够从分子水平上间接地反映生物间的遗传变异.同工酶电泳技术以生化表现型反映基因型,已成为真菌亲缘关系研究的一项重要辅助手段,在菌物分类和鉴定中得到了充分的应用.
[1] Majer D, Mithen R, Lewis B G, et al. The use of AFLP fingerprinting for the detection of genetic variation in fungi[J]. Mycol Res, 1996, 100:1 107-1 111.
[2] 王桂清,陈捷. 玉米灰斑病菌的可溶性蛋白质及同工酶多态性[J].植物保护学报,2005,32(3):291-294.
[3] 胡书能,万贤国. 同工酶技术及其应用[M].长沙:湖南科学技术出版社,1985.
[4] 李成文. 聚丙烯酸按凝胶电泳蛋白带的特异快速染色法[J].生物化学与生物物理进展, 1983,(6):71-73.
[5] 赵玉锦,郭晓君. 蛋白质电泳实验技术[M].北京:科学出版社,1986.
[6] 罗广华. 植物SOD的凝胶电泳及酶活性的显色[J].植物生理学通讯,1983,(6):44-45.
[7] 褚西宁,白玉明. 九株根霉可溶性蛋白质和三种酶的电泳图谱比较研究[J]. 真菌学报,1994,13(2):121-126.
[8] 袁凤杰,林秉承. 毛细管电泳导论[M]. 北京:科学出版社,1998.
[9] 王桂清,曾路,马迪,等.国槐根茎腐烂病病原菌的形态与分子鉴定[J],东北林业大学学报,2017,45(5):106-110.
[10] Huss, M J, Campbell C L, Jennings D B, et al. Isozyme variation among biological species in the Gibberella fujikuroi species complex (FusariumsectionLiseola) [J].American society for microbiology, 1996, 62 (10): 3 750-3 756.
[11] 曹丽华,叶华智. 麦类穗腐镰刀菌的同工酶电泳分析[J]. 西北农业学报,2001, 10(2): 45-48.
[12] 宋丽娜,周涵韬,车建美,等. 不同寄主尖孢镰刀菌同工酶异质性的研究[J]. 厦门大学学报:自然科学版,2004,43(增刊):84-86.
[13] 朱名海,贺晓霞,杨媚,等. 3种作物纹枯病菌可溶性蛋白和酯酶同工酶的比较[J]. 华中农业大学学报,2016,35(1): 55-59.
[14] 李沛利,刘丹,叶华智,等.四川省不同来源核盘菌菌株的可溶性蛋白和酯酶同工酶的电泳图谱分析[J].植物保护, 2015, 41(4): 139-142.
[15] 李萍,叶涛,郑婷,等. 辣椒疫霉可溶性蛋白和酯酶同工酶电泳分析[J].安徽农业大学学报,2015,42(6):915-920.
[16] 陈伟群,张天宇. 十字花科上的链格孢属真菌可溶性蛋白质凝胶电泳分析[J].西北农业学报,1993,2(4):53-58.
[17] 邹庆道,陈 捷,张子君. 玉米穗、茎腐病镰刀菌相互关系的同工酶分析[J].植物保护学报,2003,30(3):279-283.
[18] 车建美, 葛慈斌, 刘波,等. 黄瓜和西瓜尖孢镰刀菌同工酶异质性分析[J].亚热带农业研究,2005,1(1):45-48.
TheIsozymeAnalysisofDifferentFusariumsp.CausingChineseScholartreeRootRotDisease
MA Di WANG Gui-qing ZHANG Sai
(School of Agronomy,Liaocheng University,Liaocheng 252059,China)
24Fusariumstrains of Chinese scholartree root rot pathogen collected from Shandong province Liaocheng area were clustered based on their soluble protein and isozyme profiles obtained through native PAGE in order to base physiologically the pathogen differentiation at protein or isozyme level. A significant diversity ofFusariumwas found in terms of spectrum change of soluble protein and isozyme including EST, SOD, PPO and POD among those strains, and the obvious difference in the numbers of bands and activities of bands with the same Rf value were also detected. These data strongly supported that isozyme polymorphism could be indicators to reveal the pathogen diversity. The results also showed that the changes of isozyme were closely related to the species and the infection site ofFusarium.
Fusariumsp.,soluble protein,isozyme,cluster analysis
2017-05-10
山东省自然科学基金项目(ZR2012CL17),聊城市科技发展计划项目(2014GJH10)资助
王桂清,E-mail:wangguiqing@lcu.edu.cn.
S792.26;Q939.5
A
1672-6634(2017)03-0064-05
