低温胁迫对大花三色堇和角堇光合作用与叶绿素荧光的影响
2017-11-13杜晓华齐阳阳李建飞刘会超
杜晓华,齐阳阳,李建飞,刘会超
(河南科技学院 园艺园林学院,河南新乡 453003)
低温胁迫对大花三色堇和角堇光合作用与叶绿素荧光的影响
杜晓华,齐阳阳,李建飞,刘会超
(河南科技学院 园艺园林学院,河南新乡 453003)
旨在探讨低温对大花三色堇和角堇光合作用的影响,为大花三色堇和角堇的抗寒栽培及育种提供参考,以1个大花三色堇资源HAR和3个角堇资源JB、E01、08H为试材,采用人工模拟低温条件(-5 ℃ 8 h,-5 ℃ 16 h,-5 ℃ 24 h,-10 ℃ 7 h,-10 ℃ 14 h)对其进行低温处理,测定其光合及叶绿素荧光参数变化。结果表明,低温胁迫导致大花三色堇和角堇叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、PSⅡ的最大光化学效率(Fv/Fm)、光化学淬灭系数(qP)和实际光化学效率(ΦPSⅡ)下降,且随低温胁迫程度增加呈下降趋势,而胞间CO2浓度(Ci)和初始荧光(F0)、非光化学淬灭系数(qN)则呈上升趋势,且变化幅度因基因型不同而存在差异。依据低温胁迫下各试材的光合和叶绿素荧光指标变化情况,角堇E01和JB耐寒性较强,其次为角堇08H,大花三色堇HAR耐寒性较差。
三色堇;低温胁迫;光合速率;叶绿素荧光
中国北方地区秋冬季园林绿化花卉种类偏少,品种较为单一,缺乏丰富的花卉烘托元旦等节日气氛。大花三色堇(Viola×wittrockiana)和角堇(Violacornuta)是堇菜科堇菜属1 a或2 a生花卉,该花卉品种繁多、色彩鲜艳、花期长、耐寒,素有“花坛皇后”的美誉[1],目前在中国北方主要用作春季花坛花卉。探索低温胁迫对大花三色堇和角堇的生理影响,对其耐寒种质资源的筛选、品种选育和抗寒栽培等具有重要的指导意义。
光合作用是植物最重要的合成代谢途径之一,也是植物对低温最敏感的生理过程之一[2]。光合和叶绿素荧光技术可无损、快捷测定低温对植物的胁迫效应,目前已在西瓜[3]、番茄[4]、茄子[5]、甜椒[6]、玉米[7]、棉花[8]、切花菊[9]、草莓[10]、杨梅[11]、彩叶草[12]等多种作物的低温逆境生理研究中得到应用。但有关低温胁迫对大花三色堇和角堇的光合和叶绿素荧光特性的影响目前鲜见报道。为此,本研究以4份大花三色堇和角堇种质资源为试材,采用叶绿素荧光技术结合光合作用系统,研究低温对大花三色堇和角堇的光合和叶绿素荧光参数的影响,以期了解其对低温的生理适应机制,为大花三色堇和角堇的引种、耐寒种质资源筛选和品种选育及抗寒性栽培提供参考。
1 材料与方法
1.1 试验材料
试材为4份大花三色堇和角堇材料,其中大花三色堇HAR为荷兰BUZZY种子公司Alpenglow品种的自交选系;角堇JB为美国高美斯公司Penny Blue的自交选系;角堇E01为上海园林科学研究所的紫花白心品种的自交选系;角堇08H为荷兰BUZZY公司Johnny jump up的自交选系。种子由河南科技学院新乡市草花育种重点实验室提供。试验材料于2014年10月播种于200孔穴盘中,2~3片真叶后移栽于10 cm×10 cm营养钵中,15~20 ℃培养。待苗长至7~10片叶时进行低温胁迫处理。
1.2 试验设计
选择长势基本一致、健壮无病虫害的植株,先在5 ℃下进行1周抗寒锻炼,然后进行低温胁迫处理。设5个低温胁迫处理:-5 ℃ 8 h(Ⅰ),-5 ℃ 16 h(Ⅱ),-5 ℃ 24 h(Ⅲ),-10 ℃ 7 h(Ⅳ),-10 ℃14 h(Ⅴ),以三色堇适宜生长温度(20 ℃)为对照(CK)。胁迫处理后,每株选取生长部位相同的叶片,分别测定叶片光合参数和叶绿素荧光参数,重复3 次。
1.3 测定指标及方法
光合和叶绿素荧光参数测定采用雅欣1611G叶绿素荧光仪测定植株叶片的叶绿素荧光参数。暗适应20 min后,测定叶片的初始荧光(F0)、PSⅡ的最大光化学效率(Fv/Fm)、PSⅡ实际光化学量子产量(ΦPSⅡ)、光化学猝灭系数(qP)和非光化学猝灭系数(qN)。光化光强度为280 μmol·m-2·s-1,饱和脉冲光强为2 500 μmol·m-2·s-1,脉冲时间1.0 s。
采用Li-6400便携式光合测定系统同步测定叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等,测定时光照强度为 800 μmol·m-2·s-1±10 μmol·m-2·s-1,CO2摩尔分数为350 μmol·mol-1±10 μmol·mol-1。
1.4 数据处理
采用DPS 3.01 软件分别进行同一材料不同胁迫条件和同一胁迫条件不同材料间的方差分析,利用Duncan’s多重比较法进行差异显著性分析。
2 结果与分析
2.1 低温胁迫对大花三色堇和角堇光合参数的影响
2.1.1 对大花三色堇和角堇净光合速率的影响 净光合速率是植物叶片在一定环境下光合能力的直观反映。从图1可知,低温胁迫导致大花三色堇和角堇叶片净光合速率下降。随着胁迫程度的增加,Pn的下降幅度增大,且各试材Pn的降低幅度不同。Ⅰ处理条件下,与CK相比,大花三色堇HAR的Pn显著下降幅度最大,降低36.4%;其次为角堇08H(Pn下降13.4%),达显著水平;而角堇JB和E01的Pn未出现显著下降。在Ⅱ处理条件下,4份材料的Pn与对照相比均显著下降,其中大花三色堇HAR和角堇08H、JB和E01的Pn分别降低59%、41.3%、30.9%和22.2%。Ⅲ与Ⅳ处理条件下结果相同,HAR、08H、JB、E01的Pn与CK相比分别降低67.8%、64.8%、55%和50%。Ⅴ处理条件下,4份试验材料的Pn比对照降低93%~96%。低温胁迫对4份三色堇试材的Pn的影响程度依次为:HAR>08H>JB>E01。
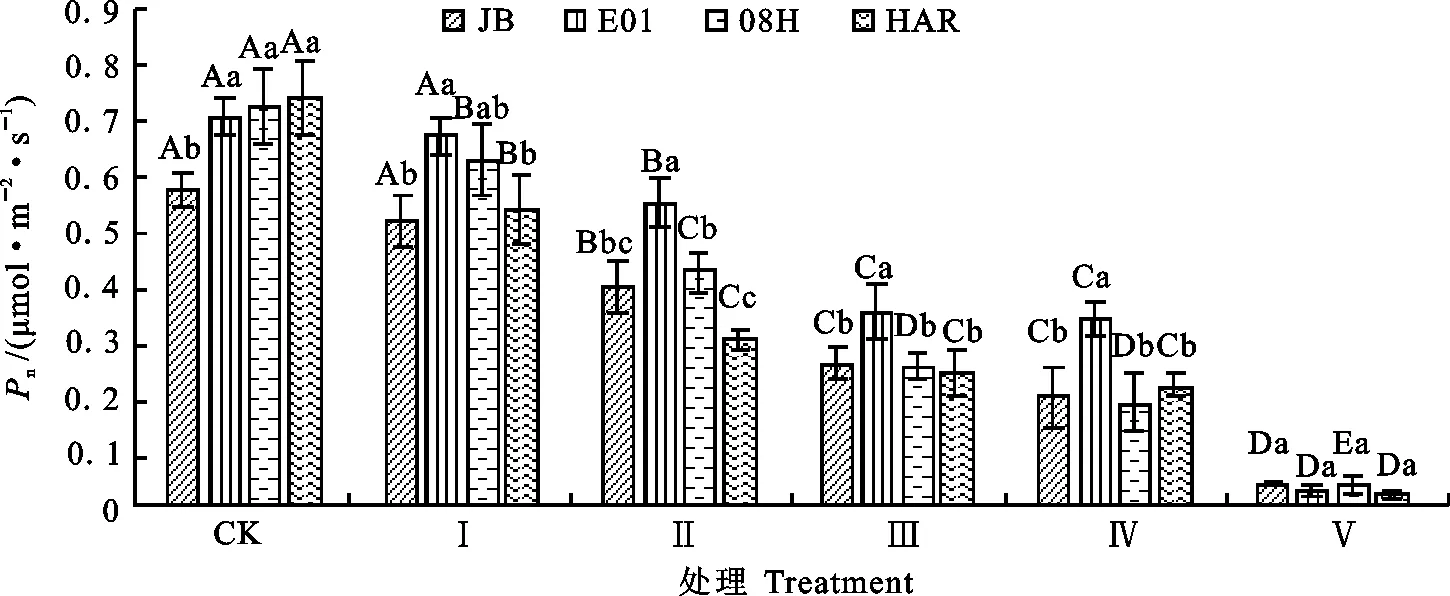
不同大写字母表示同材料在不同低温处理间的差异显著水平(P<0.05);小写字母表示同一低温处理内不同材料间的差异显著水平(P<0.05)。下图同。
Different uppercase letters represent significant difference among different treatments of the same accession(P<0.05);Lowercase letters represent the significant difference among different accessions under same treatment.The same below.
图1低温胁迫下大花三色堇和角堇净光合速率的变化
Fig.1ChangesofnetphotosyntheticrateofViola×wittrockianaandViolacornutaunderlowtemperaturestree
2.1.2 对大花三色堇和角堇Tr和Gs的影响 如图2和图3所示,4份材料的Tr和Gs随着低温胁迫程度的增加呈下降趋势。Ⅰ处理条件下,与对照相比,大花三色堇HAR的Tr和Gs分别下降21%和15.7%,达显著水平;而角堇08H、E01和JB的Tr和Gs均无显著差异。Ⅱ处理条件下,角堇08H的Tr和Gs比对照分别下降28%和25.3%,差异达显著水平;角堇E01和JB的Tr与对照相比分别下降32%、20%,差异达显著水平。Ⅲ处理条件下,4份材料的Tr和Gs显著下降,除E01外,其他3份试材较Ⅱ处理显著下降。Ⅳ处理条件下,对大花三色堇和角堇Tr和Gs的影响与Ⅲ处理相近。Ⅴ处理条件下,与对照相比,角堇JB、E01、08H和大花三色堇HAR的Tr分别下降97%、68%、91%和91%;大花三色堇HAR的Gs降低91%,角堇JB的Gs降低90%,且显著低于Ⅳ处理条件下。说明在低温胁迫下,大花三色堇和角堇叶片Gs下降,同时其Tr也相应下降。

图2 低温胁迫下大花三色堇和角堇Tr的变化Fig.2 Changes of TrViola×wittrockiana and Viola cornuta under low temperature stress
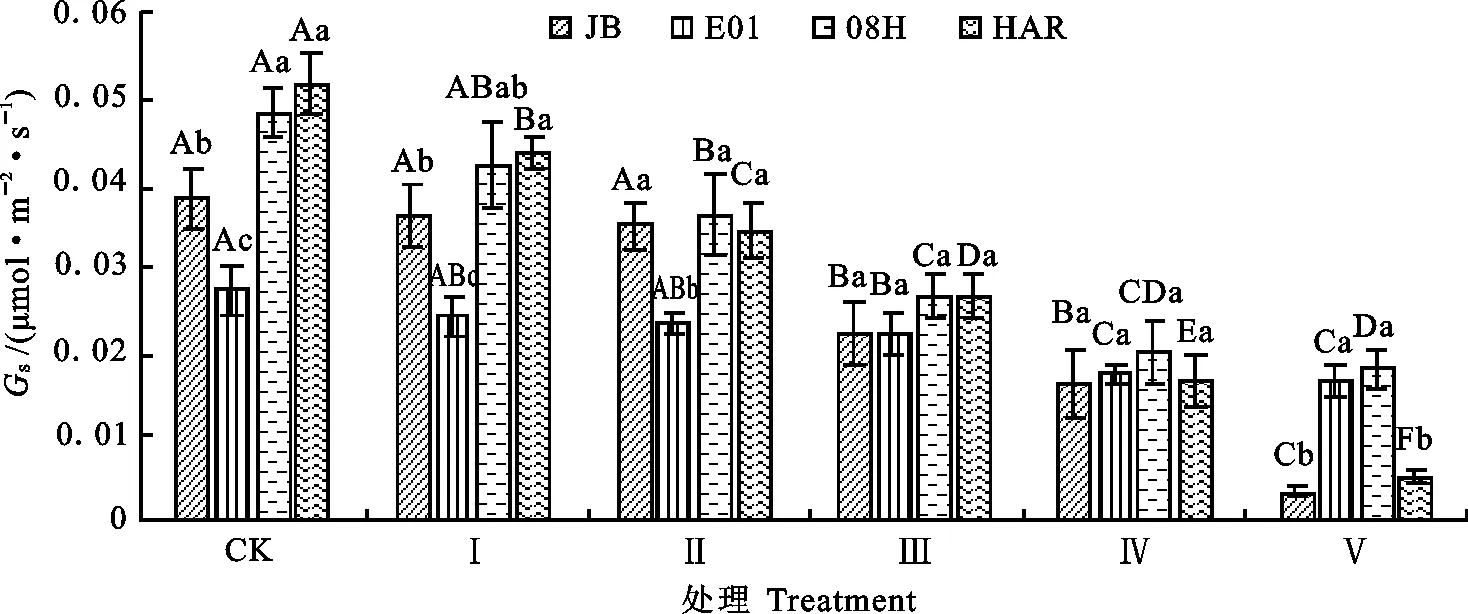
图3 低温胁迫下大花三色堇和角堇Gs的变化Fig.3 Changes of GsViola×wittrockiana and Viola cornuta under low temperature stress
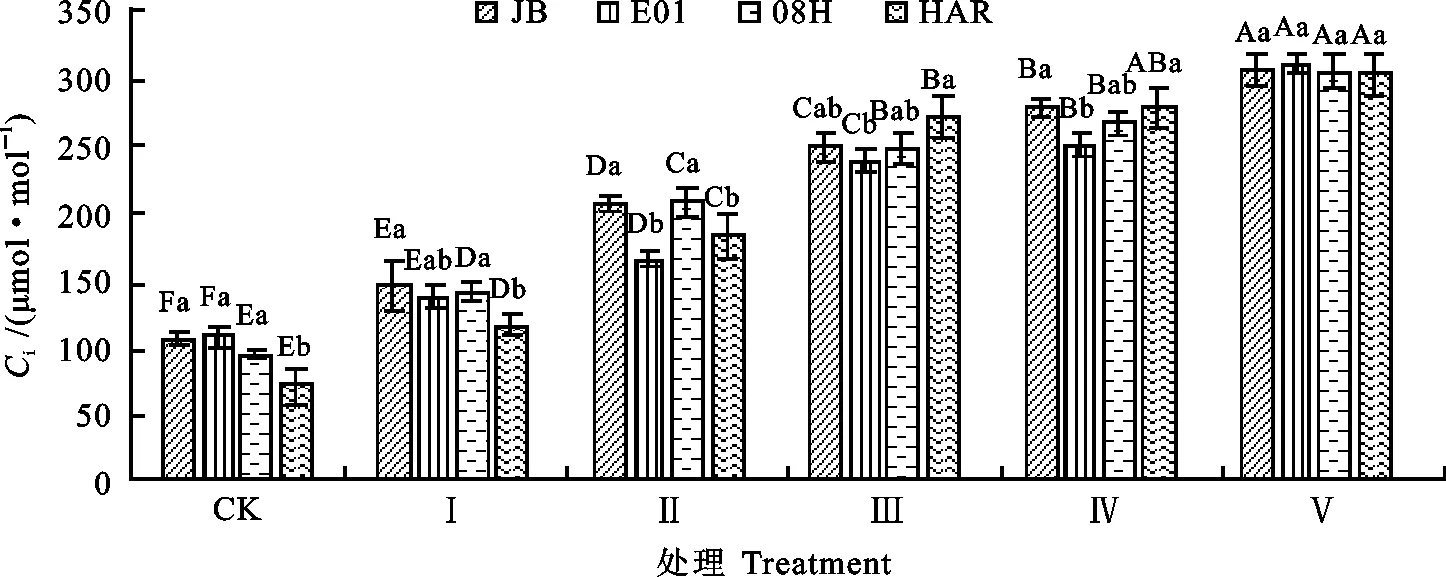
图4 低温胁迫下大花三色堇和角堇Ci的变化Fig.4 Changes of CiViola×wittrockiana and Viola cornuta under low temperature stress
2.1.3 对大花三色堇和角堇Ci的影响 由图4可知,在低温胁迫下,4份材料叶片的Ci呈增加趋势。Ⅰ处理条件下,与对照相比,4份三色堇材料的Ci均无显著差异;Ⅱ处理条件下,与对照相比,大花三色堇HAR的Ci增加21%,达显著水平,而角堇08H、E01、JB的Ci均未达显著水平;Ⅲ处理条件下,角堇08H、E01、JB的Ci分别较对照增加24%、20%、20%,差异达显著水平;Ⅳ处理条件与Ⅲ处理相比,3份角堇的Ci无显著差异。Ⅴ处理条件下,与对照相比,HAR、08H、E01、JB的Ci分别较对照增加335%、227%、192%和191%。
由图3和图4可知,大花三色堇和角堇在低温胁迫初期,Gs下降时Ci无显著下降,说明此时Pn下降既有气孔因素也有非气孔因素;随着低温胁迫程度的增加,Ci不降反升,说明胞间CO2用于光合同化部分减少,光合系统受损,Pn下降为非气孔因素所致。
2.2 低温胁迫对大花三色堇和角堇叶绿素荧光参数的影响
2.2.1 对叶片F0和Fv/Fm的影响F0是PSⅡ反应中心处于完全开放时的荧光产量,逆境引起光合机构破坏可使其升高[13]。Fv/Fm是PSⅡ最大光化学量子产量,其值降低幅度可反映植物叶片在逆境中受到的光抑制程度[14]。由图5和图6可知,Ⅰ和Ⅱ处理条件下,4份试验材料叶片的Fv/Fm和F0值与对照相比均未出现显著下降。但Ⅲ或Ⅳ处理条件下,4份材料叶片的Fv/Fm值与对照相比显著下降,表明此程度的低温胁迫已导致大花三色堇与角堇叶片的PSⅡ光能转化效率下降,出现了明显地光抑制,但此时除大花三色堇HAR的F0出现明显上升外,其余3份角堇材料的F0未出现显著上升,说明后三者的光合机构未受到伤害。Ⅴ处理条件下,4份试验材料的Fv/Fm下降幅度进一步加大,其中角堇JB、E01、08H和大花三色堇HAR的Fv/Fm值分别比对照分别下降28.5%、48.2%、66.6%、64.5%,F0均出现显著上升,表明此时低温胁迫对三色堇叶片的光抑制程度进一步加剧,光合机构遭受破坏。大花三色堇HAR的F0值上升明显,Fv/Fm值下降幅度大于角堇,说明HAR的光合器官抗寒性较差。
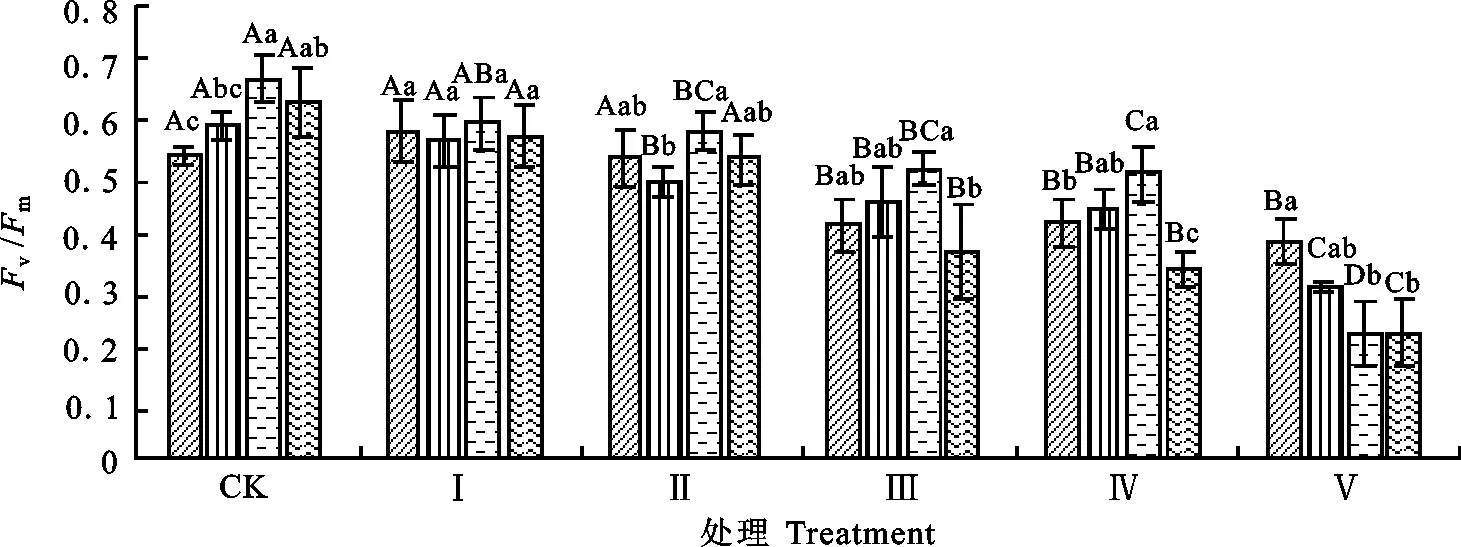
图5 低温胁迫下大花三色堇和角堇叶片Fv/Fm的变化Fig.5 Changes of Fv/FmViola×wittrockiana and Viola cornuta under low temperature stress
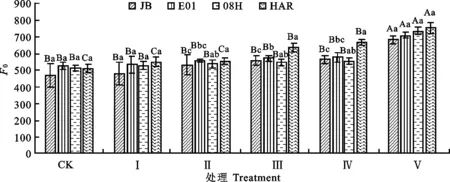
图6 低温胁迫下大花三色堇和角堇F0的变化Fig.6 Changes of FoViola×wittrockiana and Viola cornuta under low temperature stress
2.2.2 对大花三色堇和角堇ΦPSⅡ和qP的影响ΦPSⅡ反映在照光条件下PSⅡ反应中心部分关闭时的实际原初光能捕获效率[13]。qP表示PSⅡ天线色素吸收的光能用于光化学电子传递的比率,一定程度上反映PSⅡ反应中心的开放程度。qP愈大,PSⅡ的电子传递活性愈大[15]。从图7和图8可以看出,随低温胁迫程度加重,大花三色堇和角堇叶片的ΦPSⅡ和qP呈下降趋势,不同试验材料间存在一定差异。Ⅰ处理条件下,与对照相比,4份试材的ΦPSⅡ和qP未出现显著差异。Ⅱ处理条件下,与对照相比,大花三色堇HAR的ΦPSⅡ和qP分别下降18%和38%,差异达显著水平。Ⅲ处理条件时,与对照相比,角堇08H、E01和JB的ΦPSⅡ和qP在显著下降。Ⅴ处理条件下,大花三色堇HAR和角堇08H、E01、JB的ΦPSⅡ显著低于对照和Ⅲ处理条件下,与对照相比,分别降低65%、55%、41%和53%;qP与对照相比,分别降低57%、55%、44%和46%。表明低温胁迫导致大花三色堇和角堇光合电子传递效率显著下降, 且胁迫程度越高,下降幅度越大,其中大花三色堇HAR下降最大,角堇08H次之,E01下降最小。
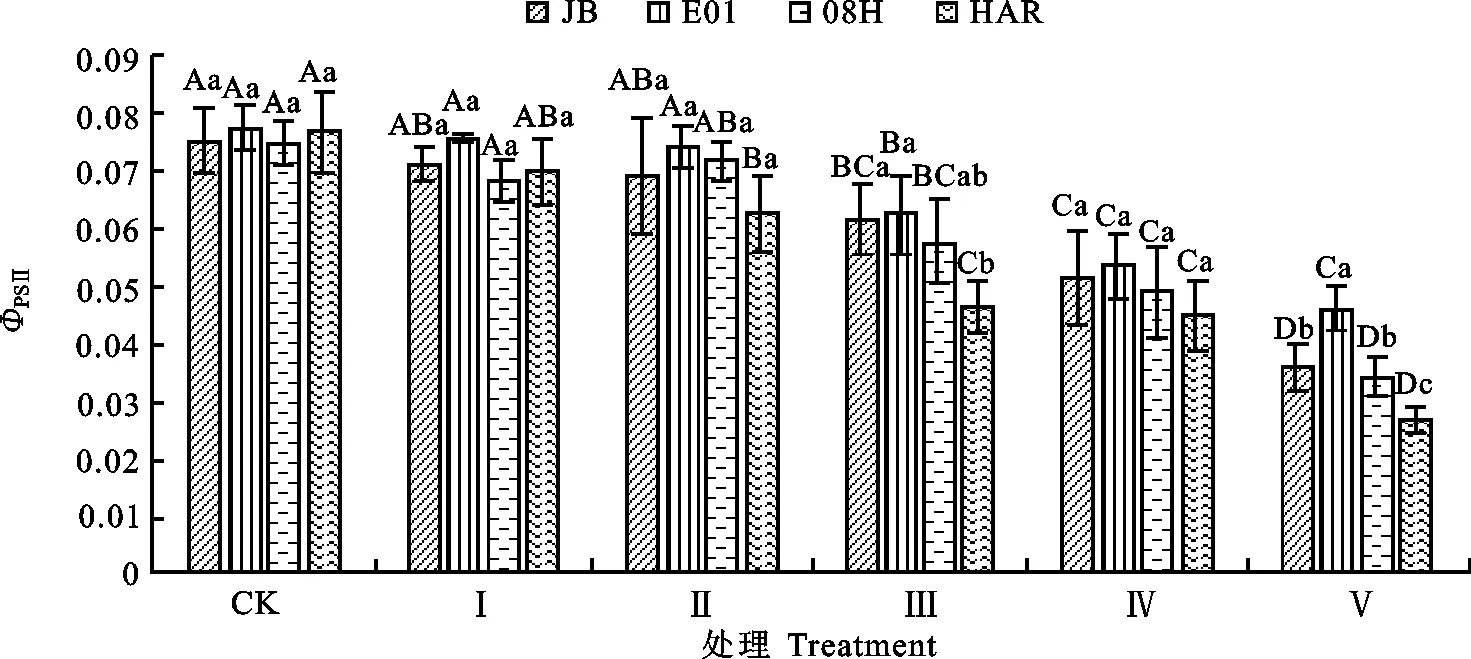
图7 低温胁迫下大花三色堇和角堇ΦPSⅡ的变化Fig.7 Changes of ΦPSⅡ Viola×wittrockiana and Viola cornuta under low temperature stress

图8 低温胁迫下大花三色堇和角堇qP的变化Fig.8 Changes of qPViola×wittrockiana and Viola cornuta under low temperature stress
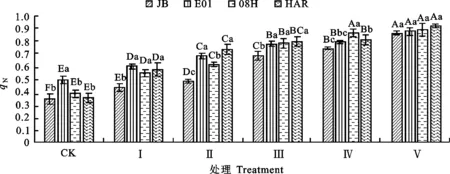
图9 低温胁迫下大花三色堇和角堇qN的变化Fig.9 Changes of qNViola×wittrockiana and Viola cornuta under low temperature stress
2.2.3 对大花三色堇和角堇qN的影响qN是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热能形式耗散掉的部分,是植物的一种自我保护机制[13]。由图9可知,与对照相比,各低温胁迫处理下4份材料叶片的qN均出现上升,说明低温胁迫导致光抑制时,其启动了热耗功能,以避免因吸收过多光能造成伤害。随着低温胁迫程度增加,各试验材料的qN上升幅度也不断增加,表明随着低温胁迫造成的光抑制加重,热耗散也在增加。低温胁迫下,不同材料间qN的增加幅度存在一定差异。Ⅴ处理条件下,大花三色堇HAR和角堇08H、E01、JB的qN分别较对照增加164%、132%、78%和154%。
3 讨 论
关于堇菜属植物的耐寒性研究,罗玉兰等[16]曾研究了上海自然低温对‘紫花地丁’与荷兰引进的三色堇的一些生理指标(电导率、含糖量、花青素含量)的影响,但该研究采用自然低温,无法考察三色堇所能承受的具体低温范围。本研究采用人工模拟低温,以中国北方新乡市冬季气温(1月份平均气温-5 ℃,极端最低气温-10 ℃[17])为参考,并根据冬季气温的日变化情况,设置不同的低温胁迫时间,采用的叶绿素荧光技术结合光合作用系统,开展了堇菜属重要的观赏植物大花三色堇和角堇的耐寒性研究,对抗寒栽培与育种实践具有重要参考价值。
低温对光合作用影响最明显的就是Pn的下降,Farquhar等[18]研究认为,低温导致光合速率下降的因素既有气孔因素也有非气孔因素。如果Pn和Gs同时下降,Ci也相应下降,即气孔因素限制占主导[3];如果Gs下降的同时Ci升高,则是由于非气孔因素阻碍了CO2的利用,造成Ci的积累所致[4,6,9]。本试验中,在-5 ℃胁迫24 h或-10 ℃胁迫7 h,大花三色堇和角堇叶片的Pn和Gs下降的同时,Ci呈上升趋势,说明此时大花三色堇和角堇叶片Pn下降不是由气孔导度下降使CO2供应减少所致,而是由于非气孔因素(如光化学活性限制、RuBP羧化限制和无机磷限制)阻碍CO2利用,造成细胞间隙CO2积累所致[19]。在低温胁迫程度较轻时(-5 ℃胁迫8 h或16 h以下),Ci未出现明显下降或上升,说明此时Pn下降既有气孔因素也有非气孔因素。
对于由非气孔因素所导致的Pn下降,Fv/Fm降低表明植物受到光抑制,初始荧光Fo的升高则表明PSⅡ失活或被破坏[13]。本研究结果表明,-5 ℃胁迫8 h或16 h,大花三色堇和角堇叶片Fv/Fm未出现明显下降,说明没有产生光抑制;而-5 ℃胁迫24 h或-10 ℃胁迫7 h,三色堇叶片出现光抑制,但F0未出现显著上升;而当-10 ℃胁迫14 h,三色堇F0出现显著上升,说明光合机构已受到低温破坏。关于Fv/Fm下降的原因,由于低温对碳同化的影响远比对其他光合过程的影响要大得多[2],因此可能是低温胁迫下Rubisco、Rubisco活化酶等活性下降导致叶片同化CO2的能力下降,引起卡尔文循环中对ATP 和NADPH的需求量下降,反馈抑制了PSⅡ电子传递[6]。本试验中,当低温胁迫程度较重(-5 ℃处理24 h或-10 ℃胁迫7 h以上),大花三色堇和角堇的PSⅡ天线色素吸收的光能用于光化学电子传递的比率(qP)和PSⅡ反应中心电子传递的量子产额(ΦPSⅡ)均明显下降,说明低温对PSⅡ产生了抑制或破坏。ΦPSⅡ下降可能与 QA→QB 的电子传递过程受到抑制有关[2]。此外,低温也降低了类囊体膜流动性,进而影响位于类囊体膜上的电子传递链和光合磷酸化过程的组分如PSⅠ、PSⅡ、细胞色素b6 f 复合体、ATP合酶等[2],造成ΦPSⅡ下降。也有研究表明,冷敏感植物如甜椒等在低温弱光下光合下降的主要原因是PSⅠ发生光抑制[20-21]。对于大花三色堇和角堇光合下降的主要原因是PSⅠ还是PSⅡ受抑制,本试验还缺乏数据进行说明。低温胁迫下三色堇qP值下降,qN上升,反映了PSⅡ天线色素吸收的光能不能完全用于光化学电子传递,而借助叶黄素循环以热形式耗散掉[2,21]。且随着低温胁迫程度加深,qN趋于升高,说明当光抑制增加时,非化学能量耗散加强,说明qN的升高是植物抵御过量光能进行自我保护的一种重要机制[11-12]。从不同低温条件对4份大花三色堇和角堇试材光合和叶绿素荧光参数的影响来看,大花三色堇HAR的耐寒性最差,而角堇E01和JB的抗寒性较强,这与通过细胞膜透性指标得出结论基本一致[22]。
Reference:
[1]张其生,包满珠,卢兴霞,等.大花三色堇育种研究进展[J].植物学报,2010,45(1):128-133.
ZHANG Q SH,BAO M ZH,LU X X,etal.Research advances in breeding of pansy(Viola×wittrockiana)[J].ChineseBulletinofBotany,2010,45(1):128-133(in Chinese with English abstract).
[2]陶宏征,赵昶灵,李唯奇.植物对低温的光合响应[J].中国生物化学与分子生物学报,2012,28(6):501-508.
TAO H ZH,ZHAO CH L,LI W Q.Photosynthetic response to low temperature in plant[J].ChineseJournalofBiochemistryandMolecularBiology,2012,28(6):501-508(in Chinese with English abstract).
[3]侯 伟,孙爱花,杨福孙,等.低温胁迫对西瓜幼苗光合作用与叶绿素荧光特性的影响[J].广东农业科学,2014,41(13):35-39.
HOU W,SUN A H,YANG F S,etal.Effects of low temperature stress on photosynthesis and chlorophyll fluorescence in watermelon seedlings[J].GuangdongAgriculturalSciences,2014,41(13):35-39(in Chinese with English abstract).
[4]王丽娟,李天来,李国强,等.夜间低温对番茄幼苗光合作用的影响[J].园艺学报,2006,33(4):757-761.
WANG L J,LI T L,LI G Q,etal.Effects of low night temperature on photosynthesis of tomato seedlings[J].ActaHorticulturaeSinica,2006,33(4):757-761(in Chinese with English abstract).
[5]郁继华,舒英杰,吕军芬,等.低温弱光对茄子幼苗光合特性的影响[J].西北植物学报,2004,24(5):831-836.
YU J H,SHU Y J,LÜ J F,etal.Influences of low temperature and poor light on photosynthetic characteristics in eggplant seedlings[J].ActaBotanicaBoreali-occidentaliaSinica,2004,24(5):831-836(in Chinese with English abstract).
[6]眭晓蕾,毛胜利,王立浩,等.低温对弱光影响甜椒光合作用的胁迫效应[J].核农学报,2008,22(6):880-886.
SUI X L,MAO Sh L,WANG Li H,etal.Effects of low temperature on photosynthesis of sweet pepper under low light[J].JournalofNuclearAgriculturalSciences,2008,22(6):880-886(in Chinese with English abstract).
[7]陈 梅,唐运来.低温胁迫对玉米幼苗叶片叶绿素荧光参数的影响[J].内蒙古农业大学学报(自然科学版),2012,33(3):20-24.
CHEN M,TANG Y L.Effects of low temperature stress on chlorophyll fluorescence characteristics of com seedlings[J].JournalofInnerMongoliaAgriculturalUniversity(NaturalScienceEdition),2012,33(3):20-24(in Chinese with English abstract).
[8]武 辉,戴海芳,张巨松,等.棉花幼苗叶片光合特性对低温胁迫及恢复处理的响应[J].植物生态学报,2014,38(10):1124-1134.
WU H,DAI H F,ZHANG J S,etal.Responses of photosynthetic characteristics to low temperature stress and recovery treatment in cotton seedling leaves[J].ChineseJournalofPlantEcology,2014,38(10):1124-1134(in Chinese with English abstract).
[9]梁 芳,郑成淑,孙宪芝,等.低温弱光胁迫及恢复对切花菊光合作用和叶绿素荧光参数的影响[J].应用生态学报,2010,21(1):29-35.
LIANG F,ZHENG CH SH,SUN X ZH,etal.Effects of low temperature-and weak light stress and its recovery on the photosynthesis and chlorophyll fluorescence parameters of cut flower chrysanthemum[J].ChineseJournalofAppliedEcology,2010,21(1):29-35(in Chinese with English abstract).
[10]郑 毅.温度胁迫对草莓叶片光合作用的影响[D].合肥:安徽农业大学,2005.
ZHENG Y.Effect of temperature stress on photosynthesis in strawberry leaves[D].Hefei:Anhui Agriculture University,2005(in Chinese with English abstract).
[11]邵 毅,徐 凯.温度胁迫对杨梅叶片光合作用的影响[D].杭州:浙江林学院,林业与生物技术学院,2009.
SHAO Y.Effect of Temperature stress on photosynthesis inMyricarubraleaves[D].Hangzhou:Zhejiang Forestry University,School of forestry and biotechnology,2009.(in Chinese with English abstract).
[12]王 兆.低温胁迫对彩叶草的生理效应及抗寒性研究[D].福州:福建农林大学,2014.
WANG ZH.Physiological responses to chilling stress inSolenostemonscutellarioidesand its inductive research[D].Fuzhou:Fujian Agriculture and Forestry University,2014(in Chinese with English abstract).
[13]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
ZHANG SH R.A discussion on chlorophyll fluorescence kinetics parameters and their significance[J].ChineseBulletinofBotany,1999,16(4):444-448(in Chinese with English abstract).
[14]陈建明,俞晓平,程家安.叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J].浙江农业学报,2006,8(1) :51-55.
CHEN J M,YU X P,CHENG J A.The application of chlorophyll fluorescence kinetics in the study of physiological responses of plants to environmental stresses[J].ActaAgriculturaeZhejiangensis,2006,8(1) :51-55(in Chinese with English abstract).
[15]李 晓,冯 伟,曾晓春.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006,26(10):2186-2196.
LI X,FENG W,ZENG X CH.Advances in chlorophyll fluorescence analysis and its uses[J].ActaBotanicaBoreali-OccidentaliaSinica,2006,26(10):2186-2196(in Chinese with English abstract).
[16]罗玉兰,陆 亮,王泰哲.本地和荷兰三色堇抗寒性的比较(简报)[J].植物生理学报,2001,37(1):27-28.
LUO Y L,LU L,WANG T ZH.Comparison on chilling resistance between local pansy and holland pansy[J].PlantPhysiologyJournal,2001,37(1):27-28(in Chinese with English abstract).
[17]中国天气.2015-01-01/2015-01-31[EB/OL].http://www.weather.com.cn/weather1d/101180301.shtml.
The weather in China.2015-01-01/2015-01-31[EB/OL].http://www.weather.com.cn/weather1d/101180301.shtml(in Chinese)
[18]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].AnnualReviewofPlantPhysiology,1982,33(1):74-79.
[19]ALLEN D J,ORT D R.Impact of chilling temperatures on photosynthesis in warm climate plants[J].TrendsinPlantScience,2001,6(1):36-42.
[20]LI X G,BI Y P,ZHAO S J,etal.The susceptibi lity of cucumber and sweet pepper to chilling under low irradiance is related to energy dissipation and water-water cycle[J].Photosynthetica,2003,41(2):259-265.
[21]MÜLLER P,LI X P,NIYOGI K K.Non-photochemical quenching.A response to excess light energy[J].PlantPhysiology,2001,125(4):1558-1566.
[22]齐阳阳,杜晓华,王梦叶,等.大花三色堇和角堇对低温胁迫的生理响应及其抗寒性研究[J].江苏农业科学,2017,45(15):115-118.
QI Y Y,DU X H,WANG M Y,etal.Physiological response and cold resistant ofViola×wittrockianaandViolacornutato low temperature[J].JiangsuAgricultureScience,2017,45(15):115-118(in Chinese with English abstract).
EffectofLowTemperatureStressonPhotosynthesisandChlorophyllFluorescenceofViola×wittrockianaandViolacornuta
DU Xiaohua,QI Yangyang,LI Jianfei and LIU Huichao
(School of Horticulture and Landscape Architecture,Henan Institute of Science and Technology,Xinxiang Henan 453003,China)
In order to understand effects of low temperature on photosynthesis ofViola×wittrockianaandViolacornuta,and provide reference for cultivation and breeding under environment of low temperature ,an experiment was conducted with aViola×wittrockianagermplasm,named HAR,and threeViolacornutagermplasm,named JB,E01 and 08H.The photosynthetic and chlorophyll fluorescence parameters of 4 pansies were measured after the plants treated under five low temperature conditions,namely,-5 ℃ for 8 h,16 h and 24 h,-10 ℃ for 7 h and 14 h.The results showed that under low temperature stress,the net photosynthetic rate(Pn),stomatal conductance(Gs),transpiration rate(Tr),Fv/Fm,photochemical quenching coefficient(qP) and actual photochemistry(ΦPSⅡ) of pansies decreased with the increace of low temperature stress,while the intercellular CO2concentration(Ci),initial fluorescence(F0) and non-photochemical quenching coefficient(qN) increased.The degree of change was different among four genotypes.According to changes of photosynthetic and chlorophyll fluorescence parameters of the tested materials,we found that E01 and JB had stronger cold tolerance,the next one was 08H,while the cold tolerance of HAR was poor relatively.
Viola×wittrockiana;Low temperature stress;Photosynthetic rate;Chlorophyll fluorescence
2016-11-16
2016-12-12
Science and Technology in Henan Province(No.152102110088).
DU Xiaohua,male,Ph.D,associate professor.Research area:genetics and breeding of ornamental plants.E-mail: duxiaohua0124@sina.com
S681.9
A
1004-1389(2017)10-1499-08
日期:2017-10-18
网络出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20171018.1733.030.html
2016-11-16
2016-12-12
河南省科技攻关(152102110088)。
杜晓华,男,博士,副教授,研究方向为观赏植物遗传育种。E-mail:duxiaohua0124@sina.com
(责任编辑:史亚歌Responsibleeditor:SHIYage)
