甘薯MADS-box转录因子IbMADS11-Like的克隆及胁迫响应分析
2017-11-11谈传婷宋炜涵朱明库董婷婷李宗芸
谈传婷 宋炜涵 朱明库 徐 涛 董婷婷 李宗芸
(江苏师范大学生命科学学院,徐州 221116)
甘薯MADS-box转录因子IbMADS11-Like的克隆及胁迫响应分析
谈传婷 宋炜涵 朱明库 徐 涛 董婷婷*李宗芸
(江苏师范大学生命科学学院,徐州 221116)
MADS-box蛋白在植物生长发育及抗逆等过程中均发挥重要功能。本实验室根据甘薯近缘野生种I.trifida基因组序列,在甘薯栽培种徐薯22(Ipomoeabatatas(L.) Xu22)中克隆到一个STMADS11亚家族MADS-box基因,命名为IbMADS11-Like。实时定量RT-PCR分析表明,IbMADS11-Like基因在甘薯根中大量表达,并且随着块根的形成和膨大表达量逐渐降低,表明该基因可能参与了甘薯块根的发育过程。胁迫处理分析表明,IbMADS11-Like基因的表达受干旱、盐和高温的诱导,而低温则抑制其表达。此外,IbMADS11-Like基因对ABA、IAA、ZT、BR、ACC、JA及GA等激素的处理也有不同程度的响应,暗示IbMADS11-Like基因可能参与了甘薯生长发育及胁迫的调控过程。这些结果为进一步分析IbMADS11-Like基因在甘薯块根发育和胁迫响应中的功能奠定了基础。
MADS-box蛋白;IbMADS11-Like;转录因子;克隆;非生物胁迫;甘薯
MADS-box蛋白是一类广泛存在于真核生物中的转录因子,在N端存在一段高度保守的DNA结合域,称为MADS(MCM1,AG,DEF and SRF)结构域[1]。植物中的MADS-box蛋白大多为MIKC型MADS-box蛋白,这种蛋白除了MADS结构域外还含有I结构域、K结构域和C结构域[2]。
MIKC型MADS-box蛋白在植物中主要参与花器官的建成[3]、开花时间[4]和果实成熟[5]等生殖生长发育过程。研究表明MIKC型MADS-box蛋白在植物的营养生长过程中也发挥了重要作用[6]。SQUA(SQUAMOSA)亚家族MADS-box基因在植物的花器官和营养组织中都大量表达。例如,马铃薯POTM1(potatoMADS-box1)基因在花分生组织和营养器官中大量表达,沉默该基因导致马铃薯块茎产量降低、侧芽生长增强[7]。AGL17亚家族MADS-box基因在营养器官中特异表达。例如,拟南芥ANR1基因在根中大量表达,参与侧根的生长发育,并且调控根对硝酸盐的吸收过程[8]。STMADS11亚家族基因也在促进植物营养生长过程发挥重要作用。例如,马铃薯STMADS11和STMADS16基因都促进了植物的营养生长过程[9~10]。
近年来,有报道表明MADS-box蛋白也参与了植物胁迫和激素响应过程。2008年,Lee[11]和Khong[12]的研究表明,AGL12亚家族MADS-box基因OsMADS26与水稻生物胁迫和非生物胁迫响应密切相关,降低该基因的表达,水稻对干旱和病原菌的抗性增强。Arora等[13]的研究也表明,水稻OsMADS18、OsMADS22、OsMADS26和OsMADS27基因在低温和干旱处理中上调表达至少2倍,而OsMADS2、OsMADS30和OsMADS55基因在干旱和盐胁迫条件下调表达。Lee[14]和Duan[15]的结果显示,STMADS11亚家族基因OsMADS22、OsMADS47和OsMADS55都是油菜素内酯(BR)信号途径的负调控因子。
甘薯(Ipomoeabatatas(L.) Lam.)是世界上重要的粮食、蔬菜、饲料、工业原料和生物能源作物,在我国的粮食安全中发挥着重要作用。目前,甘薯因其特有的根形态特征及其抗旱耐盐的生理特征,已成为块根发育及抗逆研究的重要植物。近年来随着分子生物学技术的发展,已从甘薯中克隆了一些MADS-box基因。2002年,Kim等[16]在甘薯中克隆到两个STMADS11亚家族MADS-box基因IbMADS3和IbMADS4,分析发现这两个基因都在营养器官中大量表达,并且在纤维根和色素根中表达最强。进一步分析发现,IbMADS3和IbMADS4基因在甘薯块根形成层中大量表达,表明这两个基因可能参与了甘薯块根的形成和膨大过程。2005年,Kim等[17]又报道了3个可能与甘薯块根发育相关的MADS-box基因IbMADS79、IbAGL17和IbAGL20,这3个基因分别属于AP1、AGL17和TM3-like亚家族。表达模式分析表明,3个基因均在根中表达。2008年,Ku等[18]克隆到一个AGL17家族基因IbMADS1,该基因在块根发育过程中大量表达。在马铃薯中异源超表达IbMADS1基因发现,转基因马铃薯表现出侧根膨大现象,表明IbMADS1可能是块根膨大过程的一个关键调控因子。Noh等[19]在甘薯中克隆了一个与IbMADS1同源性高达99%的MADS-box基因SRD1。表达模式分析表明,SRD1只在根中表达,并且在纤维根中表达较弱,在初始膨大的块根中表达增强。在甘薯中超表达SRD1基因,组织培养阶段的转基因植株表现为纤维根变短增粗的现象,后生木质部和形成层细胞显著增加,表明该基因通过生长素调控了根的后生木质部和形成层的形成,进而调控了甘薯块根的膨大。
目前,虽然已从甘薯中克隆到了一些MADS-box基因,但只对这些基因在块根的发育过程进行了初步分析,MADS-box基因与甘薯抗逆性相关分析仍未见报道。本实验室根据甘薯近缘野生种I.trifida基因组数据,在甘薯栽培种Ipomoeabatatas(L.) Xu22中克隆到一个新的STMADS11亚家族MADS-box基因,命名为IbMADS11-Like。利用实时定量RT-PCR对IbMADS11-Like基因在甘薯各组织的表达模式进行了研究,并利用非生物胁迫和激素处理研究了该基因与甘薯抗逆性之间的关系,为进一步分析IbMADS11-Like基因的功能奠定了基础。
1 材料与方法
1.1 实验材料、主要试剂及主要仪器
实验材料:供试甘薯徐薯22(Ipomoeabatatas(L.) Xu22)薯块由中国徐州甘薯研究中心提供。薯块栽种于江苏师范大学温室(光照16 h,28℃/黑暗8 h,18℃)。
主要试剂:常规限制性内切酶、r-Taq酶、PrimeSTAR®高保真酶、M-MLV反转录酶、DNA Ligation Kit Ver.2.0、pMD18-T载体、限制性内切酶以及Trizol RNA提取试剂盒均购自大连TaKaRa公司。DNA Marker(DL2000及DL2000 plus)购自北京全式金生物技术有限公司。DNA纯化试剂盒、氨苄青霉素(Amp)、生长素(IAA)、玉米素(ZT)、脱落酸(ABA)、油菜素内脂(BR)、乙烯前体1-氨基环丙烷-1-羧酸(ACC)、赤霉素(GA)购自Omega和Sigma公司。大肠杆菌E.coli DH5α为本实验保存。
主要仪器:MyCycler型PCR仪(美国Bio-Rad);Nanodrop ND-2000型超微量核酸蛋白测定仪(美国Thermo);凝胶电泳仪(美国Bio-Rad);超净工作台(中国安泰公司);GL25M型台式高速冷冻离心机(德国Eppendorf);GelDocXR型凝胶成像仪(美国Bio-Rad);Stepone plus型定量RCR仪(美国ABI)。
1.2 实验方法
1.2.1甘薯各组织总RNA的提取和cDNA第一条链的合成
选取3个月苗龄甘薯,取茎、叶、纤维根、铅笔根(直径1 cm)、发育中的块根(直径3 cm)和成熟块根(直径>10 cm),液氮研磨成粉,利用Tizol RNA提取试剂盒说明书分别提取各组织总RNA。0.8%琼脂糖凝胶电泳检测RNA的完整性。Nanodrop测定各组织RNA浓度,并取1μg RNA,参照反转录试剂盒说明书(M-MuLV,Takara),以Oligo d(T)18(5′TTTTTTTTTTTTTTTTTT3′)为引物合成各组织cDNA。
1.2.2 甘薯IbMADS11-Like基因的筛选及克隆
利用已知甘薯MADS-box基因(IbMADS1、IbSRD1、IbMADS3、IbMADS4、IbMADS10等)的MADS保守区序列,对甘薯近缘野生种I.trifida基因组序列进行比对,筛选到一个新的MADS-box基因序列,命名为IbMADS11-Like。根据I.trifida基因组序列设计引物IbMADS-F及IbMADS-R(表1),以栽培种甘薯徐22纤维根、铅笔根和块根的混合cDNA为模板,用PrimeSTAR HS DNA聚合酶扩增IbMADS11-Like基因全长。PCR体系如下:5×PS Buffer 10 μL、2.5 mmol·L-1dNTPs 4 μL、cDNA 1.5 μL、IbMADS-F/R引物(10 μmol·L-1)各1.5 μL、PrimeSTAR HS DNA聚合酶0.5 μL,加ddH2O至50 μL。PCR反应条件如下:98℃,变性1 min→[98℃,变性10 s→56℃,退火10 s→72℃,延伸1.5 min]×35循环→72℃,延伸10 min→4℃,保存。利用1.0%琼脂糖凝胶电泳对PCR产物进行检测。检测无误后,加A尾连接pMD18-T载体,并转化到大肠杆菌DH5α感受态细胞中。氨苄青霉素(50 mg·L-1)筛选阳性转化子。将酶切鉴定正确的菌株送上海生工测序。

表1 引物序列
1.2.3 IbMADS11-Like基因的信息学分析
利用ExPASy(Expert Protein Analysis Software,http://web.expasy.org/)在线软件对目的蛋白进行等电点、疏水性等基本性质分析。利用NetPhos 2.0(http://www.cbs.dtu.dk/services/NetPhos/)对目的蛋白进行磷酸化位点预测。利用PSORT Prediction对目的基因进行亚细胞定位预测。利用DNAMAN对MADS-box蛋白进行序列多重比对,MEGA5.1软件构建系统发进化树。利用NCBI的BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi)和Conserved Domain Search Service(CD Search)(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)对目的基因进行序列同源分析和保守结构域分析。
1.2.4 IbMADS11-Like基因的表达特性分析
根据IbMADS11-Like的基因序列设计特异定量引物IbMADS11-Like-FQ及IbMADS11-Like-RQ(表1),以β-tubulin基因为内参设计引物tubulin-F及tubulin-R(表1),以3个月苗龄的甘薯徐22茎、叶、纤维根、铅笔根(直径1 cm)、发育中的块根(直径3 cm)和成熟块根(直径>10 cm)cDNA为模板,进行实时定量RT-PCR分析。体系如下:2×SYBR Premix Ex Taq 5.0 μL、IbMADS11-Like-FQ/RQ混合引物(10.0 μmol·L-1) 0.5 μL、cDNA 1.0 μL,加ddH2O至10.0 μL。PCR反应程序如下:95℃,30 s→[95℃,5 s→60℃,30 s→信号收集]×39循环→[65℃+0.5℃/Cycle,2 s→信号收集]×60循环。利用2-△△CT法进行数据处理。
1.2.5 非生物胁迫处理
选取1个月苗龄甘薯,剪取包含2~3片叶片的枝条(约10 cm)于无菌离子水中,置于光照箱中培养(光照16 h,28℃/黑暗8 h,18℃),无菌离子水每3天更换一次。7~8 d后选取株高及长势一致的薯苗进行盐、干旱、高温和低温处理,所有胁迫处理实验均分别于0、1、3、6、12、24、48、72 h取根组织作为样品。盐胁迫处理:将甘薯幼苗转移至含有100 mmol·L-1NaCl的水溶液,根部避光培养。干旱胁迫处理:将甘薯幼苗转移至含有100 mmol·L-1PEG的水溶液,根部避光培养。高温处理:将甘薯幼苗转移至40℃,根部避光。低温处理:将甘薯幼苗转移4℃,根部避光培养。上述材料采取后立即液氮冷冻处理,并放于-80℃备用。各材料合成cDNA后,以IbMADS11-Like-FQ/RQ为引物,以tubulin为内参,进行实时定量RT-PCR分析。
1.2.6 激素处理
选取1个月苗龄甘薯,剪取包含2~3片成熟叶的枝条(约10 cm)于无菌离子水中,置于温室中培养(光照16 h,28℃/黑暗8 h,18℃),无菌离子水每3天更换一次。7~8 d后选取株高及长势一致的薯苗进行激素处理。将甘薯幼苗分别转移至100 mmol·L-1ABA溶液、100 mmol·L-1BR溶液、100 mmol·L-1ACC溶液、100 mmol·L-1IAA溶液、100 mmol·L-1GA溶液、100 mmol·L-1JA溶液中,根部避光培养,分别于0、1、3、6、12、24、48、72 h取根组织。上述材料采取后立即液氮冷冻处理,并放于-80℃备用。各材料合成cDNA后,以IbMADS11-Like-FQ/RQ为引物,以tubulin为内参,进行实时定量RT-PCR分析。
2 结果与分析
2.1 IbMADS11-Like基因的克隆
利用甘薯中已知的MADS-box基因MADS区保守序列对I.trifida基因组数据库进行筛选,获得一个新的STMADS11亚家族MADS-box基因。根据I.trifida基因组数据库序列设计引物IbMADS-F/R,以甘薯纤维根、铅笔根、块根混合cDNA为模板,克隆到一条CDS区长为648 bp的基因,命名为IbMADS11-Like(图1A)。将该序列克隆到pMD18-T载体中,限制性内切酶EcoRⅠ和SphⅠ 37℃保温3 h,切出一条长度约400 bp的条带(图1B)。将该质粒送至上海生工测序,测序结果与I.trifida基因组数据库序列比对显示,两序列并不完全一致,相似度为91.7%(图1C)。

图1 IbMADS11-Like基因的克隆及序列比对 A. IbMADS-Like基因的克隆;B. pMD18-T::IbMADS11-Like质粒酶切验证;C. Ipomoea batatas(L.) Xu22及I.trifida中IbMADS-Like基因的序列比对 M1. DL2000 Marker;1,2. IbMADS11-Like目的片段;M2. DL2000 Plus Marker;3. pMD18-T::IbMADS11-Like质粒EcoRⅠ及SphⅠ的酶切结果;trifida. I.trifida中的IbMADS-Like基因序列;Xu22. Ipomoea batatas(L.) Xu22中的IbMADS11-Like基因序列Fig.1 Electrophoretogram of the cloning of IbMADS11-Like and sequence aligment A. Amplification of target gene IbMADS-Like; B. Digestion of pMD18-T::IbMADS11-Like vector; C. Sequence aligment of IbMADS-Like in Ipomoea batatas(L.) Xu22 and I.trifida; M1. DL2000 Marker; 1,2. Target DNA of IbMADS11-Like; M2. DL2000 Plus Marker; 3. Vector of pMD18-T::IbMADS11-Like digested by EcoRⅠ & SphⅠ; trifida. IbMADS-Like sequence of I.trifida; Xu22. IbMADS11-Like sequence of Ipomoea batatas(L.) Xu22
2.2 IbMADS11-Like基因的生物信息学分析
2.2.1IbMADS11-Like基因及蛋白序列的理化性质分析
DNA START、ExPASy分析发现,该基因编码一个由216个氨基酸残基组成的蛋白质,其中包含32个负电荷氨基酸残基(Asp+Glu)和35个正电荷氨基酸残基(Arg+Lys)。分析发现,该蛋白质分子量为24.3 kD、理论等电点为8.59。通过ExPASy分析发现,IbMADS11-Like蛋白的脂溶指数(aliphatic index)为81.63,总亲水平均值(Grand average of hydropathicity(GRAVY))为-0.581,表明该蛋白质为亲水脂溶性蛋白。ExPASy的Tmpred预测(http://www.ch.embnet.org/software/TMPRED_form.html)显示,IbMADS11-Like蛋白的34~58位氨基酸区域内具有跨膜结构。利用PSORT Prediction进行亚细胞定位预测,发现IbMADS11-Like基因定位于细胞核,这与MADS-box基因的亚细胞定位保持一致。NetPhos 2.0在线磷酸化位点预测显示,IbMADS11-Like蛋白有29个磷酸化位点,其中丝氨酸(Ser)磷酸化位点21个、苏氨酸(Thr)磷酸化位点5个、酪氨酸(Tyr)磷酸化位点3个,表明该蛋白在翻译后存在翻译后修饰。
2.2.2MADS-box蛋白序列同源比对及系统进化树分析
根据已知MADS-box蛋白的结构域序列,以及NCBI Conserved Doman Research(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)分析发现,IbMADS11-Like蛋白序列含有MIKC型MADS-box蛋白保守的MADS结构域和K结构域(图2)。利用DANMAN对IbMADS11-Like及甘薯中已知的MADS-box蛋白序列进行比对,结果显示IbMADS11-Like与其他甘薯MADS-box蛋白在MADS区和K区相似度较高(图2)。利用MEGA5.1及Cluatal X对IbMADS11-Like、甘薯已知MADS-box蛋白、马铃薯MADS-box蛋白及拟南芥MADS-box蛋白进行系统进化树构建,结果表明IbMADS11-Like蛋白与马铃薯STMADS11蛋白同源性较高,属于STMADS11亚家族(图3)。我们推测IbMADS11-Like可能与STMADS11亚家族蛋白相同,在植物的营养器官发育中发挥重要功能。

图2 IbMADS11-Like与甘薯中其他MADS-box蛋白的序列比对 方框.MADS区;虚线.I区;双横线.K区;单横线.C区Fig.2 Homology comparison of IbMADS11-Like and other MADS-box proteins in sweet potato Box. MADS box; Dotted line. I box; Double line. K box; Single line. C box

图3 IbMADS11-Like与其他MADS-box蛋白的系统进化树分析 0.1. 每100个氨基酸中有10个不同Fig.3 Phylogenetic analysis of IbMADS11-Like and other MADS-box proteins 0.1. 10% changes were observed between two sequence
2.3IbMADS11-Like基因的组织特异性表达分析
为研究IbMADS11-Like基因在甘薯各组织的表达特性,本实验提取了3个月苗龄薯苗茎(S)、叶(L)、纤维根(FR)、铅笔根(DR1)、发育中的块根(DR2)、成熟块根(MR)组织总RNA,琼脂糖凝胶电泳结果显示各组织RNA都具有良好的完整性(图4A)。利用Nanodrop检测了各RNA的浓度,取1 μg合成了各组织cDNA。实时定量RT-PCR分析发现,IbMADS11-Like基因在茎和叶中表达量较低,在纤维根和铅笔根中大量表达,并且随着块根的发育表达量逐渐降低(图4)。这个结果表明,IbMADS11-Like基因可能与甘薯的块根发育密切相关。

图4 甘薯各组织RNA及IbMADS11-Like基因的表达特性分析 A.甘薯各组织总RNA;b. IbMADS11-Like在甘薯各组织中的表达特性分析;L.叶;S.茎;FR.纤维根;DR1.发育中的块根(直径2 cm);DR2.发育中的块根(直径3 cm);MR.成熟块根Fig.4 RNAs from different tissues of sweet potato and the expression pattern of IbMADS11-Like A. RNAs from different tissues of sweet potato; b. The expression pattern of IbMADS11-Like; L. Leaf; S. Stem; FR. Fiber root; DR1. Development root(The diameter is 1 cm); DR2. Development root(The diameter is 3 cm); MR. Mature storage root

图5 胁迫处理对IbMADS11-Like基因表达的影响A. NaCl;B.干旱;C.低温;D.高温Fig.5 Impacts of stresses on the expression of IbMADS11-Like A.NaCl; B.Drought; C.High tempreture; D.Low tempreture
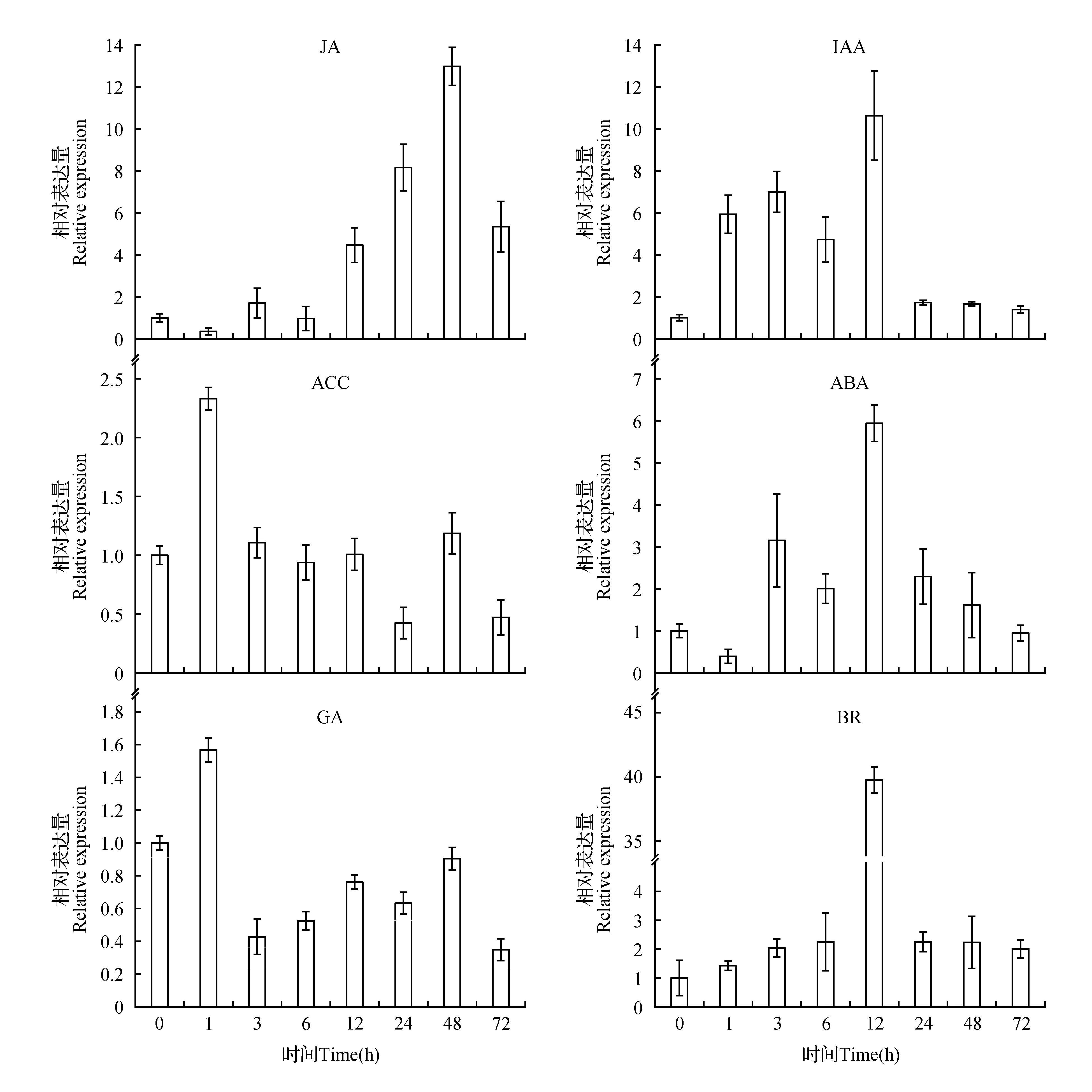
图6 激素处理对IbMADS11-Like基因表达的影响Fig.6 Impacts of hormonal treatments on the expression of IbMADS11-Like
2.4非生物胁迫对IbMADS11-Like基因表达的影响
为分析IbMADS11-Like基因的功能,本实验室对薯苗进行了高盐、模拟干旱、高温和低温处理,并利用实时定量RT-PCR对IbMADS11-Like基因在各胁迫处理材料中的表达情况进行了分析。结果表明,高盐、干旱、高温、低温等非生物胁迫均对IbMADS11-Like基因造成了不同程度的影响。盐胁迫和高温胁迫使IbMADS11-Like基因表达量逐渐上升,在处理72 h后IbMADS11-Like基因的表达量分别上升了5和10倍(图5)。在干旱胁迫条件下,IbMADS11-Like基因表达量逐渐上升,在12 h达到峰值,随后表达量逐渐下降,最高表达量比对照高约25倍(图5)。低温处理条件下,IbMADS11-Like基因的表达被逐渐抑制,在72 h后表达量达到最低(图5)。以上4种胁迫处理结果暗示,IbMADS11-Like基因对非生物胁迫敏感,可能与甘薯的抗逆相关。
2.5激素处理对IbMADS11-Like基因表达的影响
为进一步分析IbMADS11-Like基因的功能,本实验室对薯苗进行了外源激素(ABA、GA、JA、IAA、ACC、BR)处理。实时定量RT-PCR结果表明,在不同的外源激素处理下IbMADS11-Like基因表现出不同的表达量变化。外源JA处理条件下,IbMADS11-Like基因表达量被逐渐诱导,在48 h时到达最高,约被诱导14倍,随后表达量逐渐降低(图6)。与JA处理的结果相似,IAA和ABA处理后IbMADS11-Like基因表达量逐渐上升,到达峰值后迅速下降,但与JA处理不同的是IbMADS11-Like基因的表达量在12 h达到峰值(图6)。ACC处理1 h时IbMADS11-Like的表达量升高,随后迅速下降到原来水平,并基本保持不变(图6)。IbMADS11-Like在GA处理1 h后被大量诱导,而后迅速下降到原来水平的50%左右,并基本保持不变(图6)。而在BR处理时,IbMADS11-Like基因的表达量在12 h后上升约40倍(图6)。这些结果表明,IbMADS11-Like基因对不同的外源激素表现出不同的敏感性。
3 讨论
MADS-box蛋白是一类庞大的转录因子家族,在植物的生长发育过程中发挥着重要功能。目前,在拟南芥、水稻、番茄等植物中,已对MADS-box蛋白进行了大量的系统分析和功能研究[2,20~21]。然而目前,在甘薯中只克隆到8个MADS-box基因,并且只对SRD1和IbMADS1进行了功能研究[18~19]。MADS-box蛋白在甘薯中的研究仍相对缺乏。因此,本实验室根据最近发表的甘薯近缘野生种I.trifida基因组序列,在栽培种甘薯Ipomoeabatatas(L.) Xu22中克隆到了一个新的MADS-box基因(图1)。系统分析表明,该基因属于STMADS11亚家族,与马铃薯STMADS11基因同源性较高(图3),命名为IbMADS11-Like。研究表明,STMADS11亚家族成员大多与植物的营养生长发育密切相关。例如,马铃薯中的STMADS11和STMADS16都被报道参与调控了植物的营养生长过程[9~10]。甘薯中的IbMADS3和IbMADS4也被报道只在根、茎、叶等营养器官中表达[16]。因此,我们推测IbMADS11-Like可能与STMADS11亚家族的其他成员相同,在营养器官生长发育过程发挥重要功能。此外,IbMADS11-Like基因在甘薯中的组织表达特性分析显示,该基因在茎、叶和根中均有表达,尤其在纤维根中表达量最高(图4),这与STMADS11亚家族的基因表达特性保持一致,进一步暗示IbMADS11-Like基因可能参与调控甘薯的营养生长过程。
定量RT-PCR结果显示,IbMADS11-Like基因的表达量随着块根的发育迅速下降,在成熟块根中达到最低,只有纤维根中的0.2%(图4),这与甘薯STMADS11亚家族成员IbMADS3和IbMADS4的表达模式[16]非常相似,表明IbMADS11-Like基因可能参与了甘薯块根的发育过程。另外,研究表明,甘薯块根的发育与植物生长物质有着密切的关系。王庆美等[22]的研究显示,甘薯块根产量与ABA的含量呈显著正相关。Eguchi等[23]表明,外源施加人工合成的细胞分裂素可以有效促进甘薯块根的形成。Noh等[19]的研究结果显示,在甘薯块根形成初期,IAA含量上升,外源施加IAA的类似物萘乙酸时,甘薯块根数量明显增加。此外,油菜素内酯和茉莉酸也被证明可能与甘薯的块根发育相关[24~25]。因此,为进一步分析IbMADS11-Like基因的功能,本实验室对薯苗进行了外源激素处理。图6显示,IbMADS11-Like基因在ABA、IAA、JA、BR等植物生长物质处理下均有不同程度的响应,这一结果进一步暗示IbMADS11-Like基因可能参与了甘薯块根的形成和发育过程。
近年来,有报道表明MADS-box蛋白也参与了植物的非生物胁迫和激素响应过程,例如水稻OsMADS26参与了干旱和病原菌的调控[12];OsMADS18、OsMADS22、OsMADS26和OsMADS27基因在低温和干旱处理条件上调表达;OsMADS2、OsMADS30和OsMADS55基因在干旱和盐胁迫条件下调表达[13]。本实验的表达模式分析结果显示,IbMADS11-Like基因在甘薯纤维根中大量表达,在其他组织表达相对较弱(图4),表明该基因可能与干旱、盐等应答相关。为进一步分析IbMADS11-Like基因在甘薯中的功能,本实验室对薯苗进行了高盐、模拟干旱、高温、低温等非生物胁迫处理以及激素处理。实时定量RT-PCR结果表明,IbMADS11-Like基因在高盐、模拟干旱、高温胁迫条件下均被不同程度的诱导,尤其在干旱条件下,IbMADS11-Like基因的表达量升高了约25倍(图5)。而在低温条件下,IbMADS11-Like基因的表达量被明显抑制(图5)。这些结果表明,IbMADS11-Like基因可能参与了甘薯的非生物胁迫响应。此外,研究表明,干旱、盐等非生物胁迫应答常与植物激素的变化相关。例如,ABA通过调节气孔的开合、渗透压及抗逆性物质的含量,在植物的抗旱、抗寒过程发挥了重要功能[26]。细胞分裂素可通过清除自由基、提高抗氧化性,提高植物对低温、水涝和干旱等逆境的抗性[27]。低温、伤害、干旱等胁迫条件都可以诱导乙烯的产生[28]。生长素和赤霉素也被证明可以促进多胺的产生来缓解植物胁迫[29]。我们的结果显示,IbMADS11-Like基因对ABA、JA、IAA、ACC、GA及BR都有不同程度的响应,这些结果进一步表明,IbMADS11-Like基因可能参与了甘薯的胁迫响应。
综合以上结果,IbMADS11-Like可能是一个甘薯块根发育和非生物胁迫相关的调控蛋白,但这仍需要进一步研究证实。因此,本实验室构建了IbMADS11-Like基因的RNAi及35S超表达载体,拟转化甘薯进一步验证IbMADS11-Like基因的功能,相关实验正在进行中。
1.García-maroto F,Carmona M J,Garrido J A,et al.New roles for MADS-box genes in higher plants[J].Biologia Plantarum,2003,46(3):321-330.
2.Parenicová L,De Folter S,Kieffer M,et al.Molecular and phylogenetic analyses of the complete MADS-box transcription factor family inArabidopsis:new openings to the MADS world[J].The Plant Cell,2003,15(7):1538-1551.
3.Mandel M A,Gustafson-brown C,Savidge B,et al.Molecular characterization of theArabidopsisfloral homeotic geneAPETALA1[J].Nature,1992,360(6401):273-277.
4.Lee J,Lee I.Regulation and function of SOC1,a flowering pathway integrator[J].Journal of Experimental Botany,2010,61(9):2247-2254.
5.Vrebalov J,Ruezinsky D,Padmanabhan V,et al.A MADS-box gene necessary for fruit ripening at the tomatoripening-inhibitor(rin) locus[J].Science,2002,296(5566):343-346.
6.Montiel G,Gantet P,Jay-allemand C,et al.Transcription factor networks.Pathways to the knowledge of root development[J].Plant Physiology,2004,136(3):3478-3485.
7.Rosin F M,Hart J K,Van Onckelen H,et al.Suppression of a vegetative MADS box gene of potato activates axillary meristem development[J].Plant Physiology,2003,131(4):1613-1622.
8.Alvarez-buylla E R,Liljegren S J,Pelaz S,et al.MADS-box gene evolution beyond flowers:expression in pollen,endosperm,guard cells,roots and trichomes[J].The Plant Journal,2000,24(4):457-466.
9.García-maroto F,Ortega N,Lozano R,et al.Characterization of the potato MADS-box geneSTMADS16 and expression analysis in tobacco transgenic plants[J].Plant Molecular Biology,2000,42(3):499-513.
10.Carmona M J,Ortega N,Garcia-maroto F.Isolation and molecular characterization of a new vegetative MADS-box gene fromSolanumtuberosumL.[J].Planta,1998,207(2):181-188.
11.Lee S,Woo Y M,Ryu S I,et al.Further characterization of a rice AGL12 group MADS-box gene,OsMADS26[J].Plant Physiology,2008,147(1):156-168.
12.Khong G N,Pati P K,Richaud F,et al.OsMADS26 negatively regulates resistance to pathogens and drought tolerance in rice[J].Plant Physiology,2015,169(4):2935-2949.
13.Arora R,Agarwal P,Ray S,et al.MADS-box gene family in rice:genome-wide identification,organization and expression profiling during reproductive development and stress[J].BMC Genomics,2007,8:242.
14.Lee S,Choi S C,An G.Rice SVP-group MADS-box proteins,OsMADS22 and OsMADS55,are negative regulators of brassinosteroid responses[J].The Plant Journal,2008,54(1):93-105.
15.Duan K,Li L,Hu P,et al.A brassinolide-suppressed rice MADS-box transcription factor,OsMDP1,has a negative regulatory role in BR signaling[J].The Plant Journal,2006,47(4):519-531.
16.Kim S H,Mizuno K,Fujimura T.Isolation of MADS-box genes from sweet potato(Ipomoeabatatas(L.) Lam.) expressed specifically in vegetative tissues[J].Plant & Cell Physiology,2002,43(3):314-322.
17.Kim S H,Hamada T,Otani M,et al.Isolation and characterization of MADS box genes possibly related to root development in sweetpotato(IpomoeabatatasL.Lam.)[J].Journal of Plant Biology,2005,48(4):387-393.
18.Ku A T,Huang Y S,Wang Y S,et al.IbMADS1(IpomoeabatatasMADS-box 1 gene) is involved in tuberous root initiation in sweet potato(Ipomoeabatatas)[J].Annals of Botany,2008,102(1):57-67.
19.Noh S A,Lee H S,Huh E J,et al.SRD1 is involved in the auxin-mediated initial thickening growth of storage root by enhancing proliferation of metaxylem and cambium cells in sweetpotato(Ipomoeabatatas)[J].Journal of Experimental Botany,2010,61(5):1337-1349.
20.Wu F,Shi X W,Lin X L,et al.The ABCs of flower development:mutational analysis ofAP1/FUL-like genes in rice provides evidence for a homeotic(A)-function in grasses[J].The Plant Journal,2016,89(2):310-324.
21.Ito Y.Regulation of tomato fruit ripening by MADS-box transcription factors[J].Japan Agricultural Research Quarterly:JARQ,2016,50(1):33-38.
22.王庆美,张立明,王振林.甘薯内源激素变化与块根形成膨大的关系[J].中国农业科学,2005,38(12):2414-2420.
Wang Q M,Zhang M L,Wang Z L.Formation and thickening of tuberous roots in relation to the endogenous hormone concentrations in sweetpotato[J].Scientia Agricultura Sinica,2005,38(12):2414-2420.
23.Eguchi T,Yoshida S.Effects of application of sucrose and cytokinin to roots on the formation of tuberous roots in sweetpotato(Ipomoeabatatas(L.) Lam.)[J].Plant Root,2008,2:7-13.
24.Ravi V,Chakrabarti S K,Makeshkumar T,et al.Molecular regulation of storage root formation and development in sweet potato[M].//Janick J.Horticultural Reviews.Hoboken,NJ:John Wiley & Sons,2014,42:157-208.
25.Bae J M,Kwak M S,Noh S A,et al.Overexpression of sweetpotato expansin cDNA(IbEXP1) increases seed yield inArabidopsis[J].Transgenic Research,2014,23(4):657-667.
26.Yoshida T,Mogami J,Yamaguchi-shinozaki K.ABA-dependent and ABA-independent signaling in response to osmotic stress in plants[J].Current Opinion in Plant Biology,2014,21:133-139.
27.Glaser R,Kiecolt-glaser J K,Marucha P T,et al.Stress-related changes in proinflammatory cytokine production in wounds[J].Archives of General Psychiatry,1999,56(5):450-456.
28.Morgan P W,Drew M C.Ethylene and plant responses to stress[J].Physiologia Plantarum,1997,100(3):620-630.
29.刘桂丰,杨传平.盐逆境条件下3个树种的内源激素变化[J].东北林业大学学报,1998,26(1):1-3.
Liu G F,Yang C P.Endogenous hormone change of three tree species under salt stress[J].Journal of Northeast Forestry University,1998,26(1):1-3.
Supported by the National Natural Science Foundation of China(31501352);Natural Science Foundation of Jiangsu Provence(BK20150229);National Training Programs of Innovation and Entrepreneurship for Undergraduate(201510320022,201510320022Z);Jiangsu Normal University doctoral degree teachers research support project(14XLR012)
introduction:TAN Chuan-Ting(1996—),female,undergraduate,Major in plant molecular biology.
date:2016-12-20
CloningandExpressionAnalysisofStressTreatmentsofaMADS-boxTranscriptionFactorinSweetPotato
TAN Chuan-Ting SONG Wei-Han ZHU Ming-Ku XU Tao DONG Ting-Ting*LI Zong-Yun
(School of Life Science, Jiangsu Normal University,Xuzhou 221116)
MADS-box proteins play important functions in the development and stress response of plants. A STMADS11 subfamily gene was cloned fromIpomoeabatatas(L.) Xu22 according toI.trifidagenome sequences, namedIbMADS11-Like. By real-time RT-PCRIbMADS11-Likewas expressed abundantly in roots, and a rapid declining trend was observed as storage root formation and development. TheIbMADS11-Likemight be involved in the storage root development of sweet potato. Stress treatment experiments showed thatIbMADS11-Likewas induced by drought, NaCl and high temperature, while was reduced by low temperature. ABA, IAA, ZT, BR, ACC, JA and GA treatments also impacted the expression ofIbMADS11-Like, indicating thatIbMADS11-Likemight play functions in the development and stress response in sweet potato. These results play a foundation for further study on the physiological function ofIbMADS11-Likein the development and stress response of sweet potato.
MADS-box protein;IbMADS11-Like;transcription factor;cloning;abiotic stress;sweet potato
国家自然科学基金(31501352);江苏省自然科学基金(BK20150229);大学生实践创新训练计划项目(201510320022,201510320022Z);江苏师范大学博士学位教师科研支持项目(14XLR012)
谈传婷(1996—),女,本科生,主要从事植物分子生物学方面的研究工作。
* 通信作者:E-mail:dtt@jsnu.edu.cn
2016-12-20
* Corresponding author:E-mail:dtt@jsnu.edu.cn
Q78;S531
A
10.7525/j.issn.1673-5102.2017.04.015
