峨嵋蔷薇(Rosa omeiensis Rolfe)节瘤结构及其与植物自身关系的研究
2017-11-11普布多吉徐爱国蒋思萍
马 超 普布多吉 陈 彬* 徐爱国 蒋思萍
(1.西藏自治区高原生物研究所,拉萨 850001; 2.中国农业科学院植物保护研究所,北京 100193)
峨嵋蔷薇(RosaomeiensisRolfe)节瘤结构及其与植物自身关系的研究
马 超1,2普布多吉1陈 彬1*徐爱国1蒋思萍1
(1.西藏自治区高原生物研究所,拉萨 850001;2.中国农业科学院植物保护研究所,北京 100193)
研究了西藏自治区拉萨市雄色保护区内不同海拔环境中峨嵋蔷薇节瘤结构、发生的情况、发生规律及其功能的生态学效应。结果显示:(1)峨嵋蔷薇节瘤是由木质部增生致密组织形成不规则的无髓的轮纹结构,节瘤膨大由于分生组织形成植物肿瘤,由大量大小宽10~15 μm×长25 μm×高30 μm的细胞组成。(2)节瘤的发生与植物自身的生长状况显著相关,节瘤发生的密度与海拔梯度的关系不大.而与盖度和植株的高度具有显著的相关性(P=0.0081<0.01)。(3)研究区域尺度范围内,峨嵋蔷薇是区域内离散分布的重要种群,可见的节瘤枝干年龄大于9年,通过节瘤实现灌丛中个体的自疏作用,保持灌丛和整个种群的竞争优势。结论:峨嵋蔷薇的节瘤是与植物自身协同进化过程中形成,节瘤实现灌丛中个体相对快速增长和死亡,实现灌丛内个体在疏减,该文丰富了自疏现象的表现形式。
峨嵋蔷薇;节瘤;重要值;自疏作用
节瘤是植物器官上的膨大部分,形成的原因主要有物理因素和生物因素两种,物理因素主要是创伤引起的,而生物因素主要是由微生物引起的,如豆科植物的根瘤菌引起的瘤状突起。当前对节瘤的研究主要集中在豆科植物的根瘤或茎瘤,其形成原因主要是通过共生的固氮菌[1~2],在我们所查到的文献中根/茎节瘤主要是豆科和蝶形花科合萌属[3~4]而未见对于峨嵋蔷薇茎节瘤研究的报道。峨眉蔷薇节瘤发生在一定的植株高度,非人为因素,属于自然形成的,但又不同于一般病理节瘤发生在局部或一侧,多数节瘤在茎部均匀发生形成膨大部分。
峨眉蔷薇(RosaomeiensisRolfe)广泛分布于中国西部省份,多生于山坡、山脚下或灌丛中,其海拔分布范围为750~4 000 m[5],而据我们的调查实际分布超过海拔4 400 m。藏语音译“色薇美朵”,藏药应用的部位主要是花瓣、根、果实[5],其果实化学成分也已进行了分析[6]。峨嵋蔷薇在西藏地区属于常见种,其形态特征上与绢毛蔷薇(Rosasericea)相近,被认为是绢毛蔷薇的变种[7],通过形态对比,西藏拉萨市雄色保护区内的蔷薇是前者[8]。中国植物志记载其存在许多变型种[5],对中国云南省峨嵋蔷薇的花、叶等表型多样性进行了分析,认为群体间的遗传变异是它的表型性状的主要变异来源[9]。迄今为止,峨眉蔷薇的节瘤特征及其机理还未引起关注。
植物种群在一定的生境条件下,种群数量会增长,当受到种群最大容量的制约,种群内部个体间的竞争会产生自疏作用使种群维持在一定的水平,自疏作用发生在种群中内部个体按照一定的规则(-3/2或-4/3法则)发生[10~12]。本文以峨嵋蔷薇为实验对象探讨其灌丛中分支个体自疏现象和其产生机制。选址西藏自治区拉萨市雄色自然保护区内雄色寺附近,由于民俗和信仰的因素,该区域植被保护相对完好[8],该文研究了保护区内峨嵋蔷薇节瘤的发生规律,来探索该植物自疏作用的机制。
2 材料方法
2.1 试验地点
野外实验于2016年9月在西藏自治区拉萨市境内的雄色自然保护区开展。保护区位于青藏高原的西藏自治区拉萨市曲水县才纳乡雄色峡谷,样地海拔在4 100 m以上,局部坡度变化差异较大。曲水县年平均气温4.5℃;年降水量565.6 mm,降水主要集中在夏季,夏季凉爽湿润,冬季寒冷干燥,雄色寺周边局部小气候适宜,植被保存完好,主要是由高山灌丛组成。整个样地坡向南至南偏西,样地内峨嵋蔷薇主要分布在坡度较大和平缓的碎石地上。样方中,优势灌丛有峨嵋蔷薇(RosaomeiensisRolfe)、鬼箭锦鸡儿(Caraganajupata(Pall.) Poir.)、拉萨小檗(Berberishemsleyana)、小蓝雪花(CeratostigmaminusStapf ex Prain),间或存在匍匐荀子(CotoneasteradpressusBois两种)、高山柳(SalixcupularisRehd.)等。
2.2 样方设计
样地在西藏自治区拉萨市雄色自然保护区内设置,共设置五个海拔梯度,根据植株高度设置样方,样方面积为10 m×10 m,每个梯度设置4个样方作为重复,样方间距30 m左右,整个实验共有样方19个(4×5-1,样线D海拔4 400 m处缺少一个样方,因存在长期人为活动,寺庙等房屋建筑外,同时还有简单的道路和水利相关设施,不同种群密度变化较大)。利用GPS记录地理位置信息,并记录样方其它的地理信息(表1)。
表1调查样地的地理信息
Table1Geographicalinformationofsamplingsitesinvestigated

编号Serialnumber纬度NLatitude经度ELongitude海拔Altitude(m)坡向Slopeaspect人为活动HumanactivitiesA129°26′42″91°1′24″4130南South+B129°26′45″91°1′29″4210南South+C129°26′51″91°1′27″4320南偏西Southwest++D129°27′7″91°1′29″4400南偏西Southwest+++E129°27′29″91°1′26″4440南偏西Southwest+
说明:+的数量表示人活动的多少。
Note:The number of + indicates how much people activities.
2.3 数据采集与分析处理
统计了每个样方内峨嵋蔷薇的种群密度(以灌丛数量计算),测定了其高度和盖度;灌丛中峨嵋蔷薇节瘤发生的数量及节瘤的高度、节瘤下部正常茎粗;统计主要优势灌丛中的密度、高度及盖度。
茎粗采用游标卡尺测量,为两次垂直方向茎粗之和的平均值;灌木重要值=(相对盖度+相对高度)/200,仅计算统计的植物,相对高度=该灌木的高度/已统计灌木的高度之和×100,相对盖度=该灌木的盖度/已统计灌木的盖度之和×100,高度使用米尺测量,盖度根据经验估算得出[13]。线性模拟与相关性分析使用SASS 9.2和Microsoft excel 2007软件进行数据分析和作图。通过sass对数据节瘤发生概率、发生个数(表2)与海拔梯度、种群密度、植株高度与盖度等进行单因素和多因素进行线形分析。

图1 节瘤和正常枝干木质显微结构 A.节瘤横断面;B-C.节瘤致密组织横断面;D.节瘤致密组织纵切;E-F.导管组织横断面;G-K.节瘤中导管组织纵切孔纹结构;L.孔纹结构;M.节瘤下正常茎轮纹;N-O.节瘤的轮纹;P-Q节瘤致密组织横断面;R正常茎纵切Fig.1 Microstructure of nodulation and normal stem A. Cross section of the nodulation; B-C. Cross section of the dense tissue of the nodulation; D. Longitudinal section the dense tissue of the nodulation; E-F. Cross section of vessels; G-K. Perforation plate structures of vessels in nodulation; L. Perforation plate structures of vessels; M. Normal stem’s annual rings; N-O. Ring lines in nodulation; P-Q. Cross section of the dense tissue of the nodulation; R. Longitudinal section of normal stem
Table2Incidenceandnumberofnodulationindifferentelevation

海拔Altitude(m)节瘤发生数量Amountofnodulation节瘤发生概率Percentageofnodulation’soccurrence(%)413056.2542101830.4343203155.00440017358.3344401041.18
2.4 样品采集与处理
显微镜(徕卡M-125、DM1000)观察样品,加工成小于1立方厘米样品,采用FAA固定液(38%福尔马林5 mL+冰醋酸5 mL+70%乙醇90 mL+甘油5 mL)固定24 h后,经过脱水、浸腊、包埋等工序后切片观察[14]。
扫描电子显微镜观察样品在中国农业科学院农产品加工所开展,节瘤及枝干大块完整组织采用FAA固定液后自然干燥样品经切片、喷金、干燥后观察[15]。
3 结果
3.1 节瘤结构特征
显微结构(图1)显示峨嵋蔷薇节瘤的形成主要是由于木质部的增长引起的,从节瘤组织显微结构体式显微镜下正常组织年轮纹路清晰,有明显的髓(图1:M),而节瘤组织中具有较多的轮纹结构,无髓结构(图1:N,O)。纵切节瘤组织(图1:P,Q)与正常枝干组织(图1:R)对比发现节瘤中形成多个植物肿瘤。扫描电镜观察发现节瘤的木质部导管被长方形的组织分割开,横切面方形细胞10~15 μm×25 μm,纵切25 μm×30 μm,随机分散一些不足1 μm的圆形孔纹(图1:B~D)。被分割开的导管和正常组织导管的直径无明显差异,直径为30 μm(图1:A,E),纵切未观测到较为完成的导管,其长度远大于导管横切直径。比较节瘤组织中导管与正常组织导管的孔纹结构,我们发现节瘤组织导管孔纹存在较大变化差异,而正常组织导管孔纹变化差异较小。节瘤木质部硬度远高于正常木质部,主要是由于大量的较小且紧密排列的近似长方形细胞组成。皮层加厚主要由于大量形成的木栓,节瘤上部和下枝干正常生长,部分会分生出若干枝条。
3.2 峨嵋蔷薇分布情况
在研究尺度范围内,随着海拔高度的增加峨嵋蔷薇种群密度(图2)无明显变化,呈现离散分布特点,拉萨小檗呈增加趋势,鬼箭锦鸡儿和小蓝雪花呈减少趋势。在海拔4 400 m处,不同种群密度变化较大,锦鸡儿种群密度急剧减小(图2),植被总盖度增加,峨嵋蔷薇盖度明显增加,植株高度提高,锦鸡儿盖度减少,植株高度无明显变化,其他物种的盖度和种群密度变化趋同(图3~4)。该尺度范围内,峨嵋蔷薇的重要值仅在最低海拔4 130 m处略低于锦鸡儿,在其他海拔梯度中均高于其他物种,是群落中的优势种(图5)。一定海拔高度后,样方受地理因素限制(被岩石峭壁阻隔)。峨嵋蔷薇种群数量急剧减少,香柏(Sabinaalpingivar.wilsonii(Rehd) Cheng ex L.K.FU)数量增加。

图2 不同海拔主要灌丛密度Fig.2 Densities of different shrubs at different altitudes

图3 不同海拔主要灌丛高度Fig.3 Height of different shrubs at different altitudes

图4 不同海拔5种主要灌丛盖度及总盖度Fig.4 Coverage and the whole coverage of different shrubs at different altitudes

图5 主要灌丛重要值Fig.5 Important values of different shrubs at different altitudes
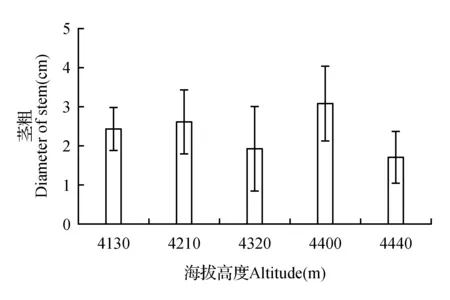
图6 节瘤下茎粗Fig.6 Diameter of stem under nodulation

图7 节瘤与植株高度对比Fig.7 Comparison between height of the rose and its nodulation
3.3 峨眉蔷薇群体灌丛数量及节瘤发生情况
峨嵋蔷薇节瘤是树干到一定高度后的主枝干膨大部分,发生在粗壮的枝条中(图6),发生情况(表2)新生枝条和细小枝条未发现有节瘤发生情况,(图1:M)为年轮数为9,该枝条为发现最细小枝干节瘤发生,可看出节瘤发生都大于9年龄的枝干上。节瘤发生高度随着植株高度增加而增加,随着植株高度降低而降低(图7)。通过对比节瘤下部正常枝干茎粗(图6)与植物盖度(图4),变化趋势一致。节瘤发生个数(表2)和盖度与高度极显著相关(P=0.008 1<0.01),节瘤发生个数Y和峨嵋蔷薇盖度X1和高度X2的线性关系为:y=542.810 57x1+0.432 34x2-73.218 22,而与海拔梯度、种群密度等其它因素间无明显相关性。因此节瘤的发生与峨嵋蔷薇植物自身的生长状况显著相关。
4 讨论
峨嵋蔷薇在大尺度范围内呈现区域分布特点,聚集分布在一定的区域,试验区小尺度范围随机分布,个体数量随着海拔升高变化较小,呈现离散分布特点,实地调查发现局部的坡度和湿度对植物分布影响较大,在局部坡度平缓的碎石地和坡度较大的坡地分布较多。参照物种多度格局马克明[16],峨嵋蔷薇种群密度和其他种群不显著相关,但其自身生长状况影响其他种群,其盖度增加显著降低锦鸡儿的盖度。峨嵋蔷薇在样地为一重要优势种和建群种,它的重要值在所有的灌木中最高,是群落中的重要组成物种。
峨嵋蔷薇作为重要物种,如何保持竞争优势的生存策略[16~17],研究发现其节瘤是树干一定高度的主枝干膨大部分,发生部位茎粗显著增加,节瘤表皮更加粗糙,节瘤上部主茎粗一般大于等于下部茎粗,同时部分节瘤部位大量分生处不同茎粗的新茎,在一个相当长的时间内,节瘤促进植物自身生长。该节瘤对该种群在环境中的生长与存在的不构成威胁,研究区域内随海拔增加节瘤发生数量与植物自身高度、盖度及灌木重要值之间线性关系良好,因此是一种长期进化中形成的结果。节瘤主要发生在成熟灌丛和灌丛中部分枝干,新生枝干未见发生节瘤,从某种程度上讲属于一种病变组织,但是该组织并未对该植物的生长和种群稳定构成存在一定的关系,相当一段时间内表现出促进植物增长。节瘤的木质部质地坚硬,密度较大,自然降解速度较慢,海拔4 400 m的灌丛中我们发现灌丛中较多已经死亡的带节瘤的枝干,该海拔灌丛生长旺盛,未见正常枝干死亡。峨嵋蔷薇作为一个的稳定种群,是一种长期进化过程中形成的自疏机制,通过发生节瘤的方式,使组织快速生长和死亡,实现个体的自疏,为新生树干的生长提供空间,作为一种生存策略[18]避免灌丛内部过密,为灌丛内部更新提供空间,保持自身的生命力和竞争力,保证灌丛生长优势和该空间尺度上的种群生物量的优势。
植物的结构是功能基础[19~20],体式显微镜下观察节瘤组织显微结构,发现正常组织年轮纹路清晰,而节瘤组织中具有较多的不规则轮纹结构,电镜观察发现节瘤的木质部导管被近似长方形的异变细胞的增长形成组织分割开,这长方形细胞对比正常组织导管孔纹少且小,且自身连接紧密,节瘤的形成是由于方形细胞的增长引起形成致密的木质,从而影响了植物疏导组织的结构和功能,实现其生态学功能。峨嵋蔷薇节瘤在实验考察区域内分布是有一定规律的,是自然状态下普遍形成的,而纵切节瘤可发现大量植物肿瘤,是否为部分形成层细胞在植物生长过程中失去分裂能力,而又在某些时间恢复,需要进一步研究。
1.Moulin L,Munive A,Dreyfus B,et al.Nodulation of legumes by members of theβ-subclass of Proteobacteria[J].Nature,2001,411(6840):948-950.
2.Long S R.Rhizobium-legume nodulation:life together in the underground[J].Cell,1989,56(2):203-214.
3.Alazard D.Stem and root nodulation inAeschynomenespp.[J].Applied and Environmental Microbiology,1985,50(3):732-734.
4.Gresshoff P.Plant Genetic Control of Nodulation inLegumes[J].Current Opinion in Plant Biology,2007,10(1):44-51.
5.中国科学院中国植物志编辑委员会.中国植物志:第37卷[M].北京:科学出版社,1985.
Flora of China Editorial Committee.Flora of China:vol.37[M].Beijing:Science Press,1985.
6.杨维力,陈封政,田军,等.峨眉蔷薇果实化学成分的研究[J].中国药学杂志,2000,35(7):445.
Yang W L,Chen F Z,Tian J,et al.Study on chemical constituents ofRosaoneienisisRolfe fruit[J].Chinese Pharmaceutical Journal,2000,35(7):445.
7.韦筱媚,高信芬,张丽兵.绢毛蔷薇复合体的分类学研究:峨眉蔷薇与绢毛蔷薇同种吗[J].植物分类学报,2008,46(6):919-928.
Wei X M,Gao X F,Zhang L B.Asystematic study ofRosasericea(Rosaceae)complex:AreR.omeiensisandR.sericeaconspecific[J].Journal of Systematics an Evolution,2008,46(6):919-928.
8.卢欣,仓决卓玛,次仁.建立西藏拉萨雄色市级自然保护区的意义及其可行性[J].西藏科技,2003,(11):26-28.
Lu X,Cangjue Z M,Ci R.Significance and feasibility of establishing nature reserve in Xiongse Lhasa city Tibet[J].Tibet Science and Technology,2003,(11):26-28.
9.周宁宁,唐开学,邱显钦,等.云南峨眉蔷薇天然群体的表型多样性[J].西南农业学报,2009,22(6):1732-1736.
Zhou N N,Tang K X,Qiu X Q,et al.Phenotypic diversity of natural populations inRosaomeiensisRolfe in Yunnan Province[J].Southwest China Journal of Agricultural Sciences,2009,22(6):1732-1736.
10.Hagihara A.Theoretical considerations on the C-D effect in self-thinning plant populations[J].Population Ecology,1999,41(2):151-159.
11.黎磊,周道玮,盛连喜.植物种群自疏过程中构件生物量与密度的关系[J].生态学报,2012,32(13):3987-3997.
Li L,Zhou D W,Sheng L X.Allometric relationship between mean component biomass and density during the course of self-thinning forFagopyrumesculentumpopulations[J].Acta Ecologica Sinica,2012,32(13):3987-3997.
12.王仁忠,方林,卢文祥,等.植物种群-3/2幂自疏定律假设的点评[J].东北师大学报:自然科学版,1998,(4):58-62.
Wang R Z,Fang L,Lu W X,et al.Some comments on the -3/2 power law of self-thinning[J].Journal of Northeast Normal University,1998,(4):58-62.
13.王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报:自然科学版,2010,33(2):312-316.
Wang Y S,Shangguan T L.Discussion on calculating method of important values[J].Journal of Shanxi University:Natural Science Edition,2010,33(2):312-316.
14.杨捷频.常规石蜡切片方法的改良[J].生物学杂志,2006,23(1):45-46.
Yang J P.Improvement of traditional paraffin section preparation methods[J].Journal of Biology,2006,23(1):45-46.
15.肖媛,刘伟,汪艳,等.生物样品的扫描电镜制样干燥方法[J].实验室研究与探索,2013,32(5):45-53,172.
Xiao Y,Liu W,Wang Y,et al.Drying methods of biological sample preparation for scanning electron microscope[J].Research and Exploration in Laboratory,2013,32(5):45-53,172.
16.马克明.物种多度格局研究进展[J].植物生态学报,2003,27(3):412-426.
Ma K M.Advances of the study on species abundance pattern[J].Acta Phytoecologica Sinica,2003,27(3):412-426.
17.刘艳芳,张艳茹,陈红.贡嘎山阔叶木本植物叶片解剖结构及其生存策略分析[J].西南大学学报:自然科学版,2015,37(5):66-72.
Liu Y F,Zhang Y R,Chen H.An analysis of the foliar anatomical structures of broadleaved woody plants in the Gongga mountains and their survival strategy[J].Journal of Southwest University:Natural Science Edition,2015,37(5):66-72.
18.Blom C W P M,Voesenek L A C J.Flooding:the survival strategies of plants[J].Trends in Ecology & Evolution,1996,11(7):290-295.
19.邓传远,辛桂亮,张万超,等.红树族植物次生木质部附物纹孔的电镜观测[J].植物学报,2015,50(1):90-99.
Deng C Y,Xin G L,Zhang W C,et al.SEM observations and measurements of vestured pits of the secondary xylem in the Tribe Rhizophoreae[J].Chinese Bulletin of Botany,2015,50(1):90-99.
20.展志岗.植物生长的结构—功能模型及其校准研究以corner型植物为例[D].北京:中国农业大学,2001.
Zhan Z G.Study on a structure-function model of plant growth and its calibration-corner-model plant as a case[D].Beijing:China Agricultural University,2001.
投稿须知
1.投稿前请登陆本刊网站(http://bbr.nefu.edu.cn/)认真阅读本刊投稿中心中的各项说明,并按照本刊要求修改和补充论文中的有关内容,使论文符合本刊要求。投稿时请附作者单位介绍信或全体作者签名,说明无泄密、署名无误和未一稿多投,网上投稿请签订版权转让协议。作者自留底稿及软盘备份。
2.投稿时,请登陆本刊网站在线提交论文。投稿后,可随时登陆查询稿件处理状况。
3.稿件如初审通过,请作者按要求邮寄已签名的《版权转让协议》。
4.稿件经过外审和编审后,如符合我刊要求,须交寄发表费,待发表费收到后,作者可登陆本刊网站后,在发稿状态下打印《稿件录用通知》。
5.作者文责自负。本编辑部对来稿可做必要修改。
6.稿件一经发表,酌致稿酬,并赠样刊2本。
7.凡在本刊发表的论文,如荣获省部级以上成果奖,请及时通知本编辑部,并提供获奖证书复印件。
《植物研究》编辑部
Natural Science Foundation of Xizang(Tibet) Autonomous Region(2015ZR-13-55)
introduction:MA Chao(1982—),male,master candidate,Mainly engaged in the study of medicinal plant.
date:2016-12-30
NodulationStructureinRosaomeiensisRolfeandItsRelationshipwithItself
MA Chao1,2PUBU Duoji1CHEN Bin1*XU Ai-Guo1JIANG Si-Ping1
(1.Tibet Plateau Institute of Biology,Lhasa 850001;2.Institute of Plant Protection,Chinese Academy of Agricultural Sciences,Beijing 100193)
We studied the occurrence of nodulation onRosaomeiensisRolfe and their regularity as well as the reasons of nodulation in the area of Xiongse Nature Reserve of Lasa in Tibet Autonomous Region, and checked the ecology effect of distribution ofR.omeiensison other shrubs Results show that: (1)The nodulation was organized by the hyperplasia of xylem, dense tissue forming irregular annulation without pulp. Its enlargement due to meristem tissue formation of plant tumors, which mainly formed by the square cells size of about 10-15 μm×25 μm×30 μm; (2)The occurrence of nodulation is closely related to the growth condition of its own. The relationship between the occurrence of nodulation, the density and the altitude gradient was not significantly great, while it has a significant correlation with the coverage and the height of the plant(P=0.0081<0.01); (3)Within the scale of the study area, nodulation can be found in over 9-yearR.omeiensis, as an important species of discrete distribution in the area with no correlation with other brush. To maintain the competitive advantage of shrub and whole population, it was achieved by self-thinning effect with nodulation. The nodulation ofR.omeiensisis formed in the long-term evolution process. The rapid growth and death of the individuals in the shrubs is realized through the nodulation. Through nodulation, the competitive advantage of brush and the entire population is kept.
RosaomeiensisRolfe;nodulation;importance value;self-thinning effect
西藏自治区地区自然科学基金(2015ZR-13-55)
马超(1982—),男,硕士研究生,主要从事药用植物研究。
* 通信作者:E-mail:32538503@qq.com
2016-12-30
* Corresponding author:E-mail:32538503@qq.com
Q949.751.8
A
10.7525/j.issn.1673-5102.2017.04.005
