光照对珙桐幼苗光合与生长特性的影响
2017-11-10姜瑞芳刘艳红
姜瑞芳 , 刘艳红
北京林业大学 林学院, 北京 100083
光照对珙桐幼苗光合与生长特性的影响
姜瑞芳 , 刘艳红*
北京林业大学 林学院, 北京 100083
为了解珙桐幼苗对光强的适应性, 以3年生珙桐幼苗为实验材料, 设置了全光照, 62%全光照及43%全光照3种光环境, 观测了110 d后珙桐幼苗光合特性及生物量分配的变化情况。结果表明: (1)珙桐幼苗净光合速率(Pn)日变化在全光照条件下呈双峰型, 有明显“光合午休”现象, 而遮阴条件下呈单峰型; (2)遮阴处理下最大光合速率(Pmax)、表观量子效率(AQY)、水分利用效率(WUE)及光能利用效率(LUE)均高于全光照, 光补偿点(LCP)和暗呼吸速率(Rd)均低于全光照; (3)遮阴显著降低珙桐幼苗根冠比, 增加了比叶面积(SLA)和总生物量。上述结果说明: 遮阴处理后, 珙桐幼苗在光合特性和生长特性上表现出对弱光一定程度的驯化适应, 对比三种遮阴条件可以得知, 适度的遮阴更有利于幼苗的正常生长, 甚至在较低光照(43%全光照)环境下有更高的光能利用率。
珙桐; 光照条件; 光合特性; 生长特性
1 前言
光照是植物进行光合作用的最重要能量来源,是影响植物生长和分布的重要环境因子之一[1]。不同种类的植物对光强的适应范围存在差异, 光强过低, 植物不能充分进行光合作用; 而光强过高, 植物又会发生光抑制。因此, 研究植物的最适宜光照条件, 有利于最大限度利用光能, 提高植物的光合效率。大量研究表明, 光强影响植物的形态特征、光合特性及生物量分配等, 如闽楠(Phoebe boumei)幼树的光合特性在不同光照条件下存在差异, 低光照限制生物量的积累[2]; 遮阴造成树莓(Rubus corchorifolius)蘖根苗数量的减少[3]。所以, 研究植物在不同光照下的适应性, 可以为其迁地保护提供重要的科学指导意义。
珙桐(Davidia involucrateBaill)是我国特有的珙桐科单型属珍稀濒危植物, 主要分布于四川、湖南、湖北、贵州和云南等地[4]。它不但具有园林价值, 也是我国出口创汇的重要树种之一[5]。但由于自然及人为原因, 珙桐资源越来越少, 分布范围也日益缩小。为了对珙桐进行更好的保护, 自20世纪70年代末开始, 我国开始对珙桐进行规模性地引种栽培[6]。研究发现, 珙桐幼苗生长缓慢, 具有喜阴湿的特性[7], 在强光照下叶片发黄, 并且伴有灼伤现象。可见, 光强过大可能是珙桐引种的限制因子, 对珙桐幼苗的光照强度适应范围的研究很有必要。韩素菊[8]等对珙桐幼苗在遮阴环境下的适应性已有研究, 并表明 50%遮光率有利于珙桐生长。本试验在前人的研究基础上, 通过设置不同遮阴处理以及选取与前人研究不同年龄段的珙桐幼苗, 深入探讨珙桐幼苗光合及生长特性对不同光照环境的响应情况,以期为珙桐幼苗的引种驯化、苗木栽培及迁地保护提供科学依据。同时, 珙桐作为园林植物, 探讨其对不同光照环境的适应性, 也可为园林植物在城市园林不同光照环境中的应用提供科学依据。
2 材料与方法
2.1 试验材料与处理方法
试验于 2015年在北京林业大学气象站进行, 供试材料为2014年3月从湖南武陵山区附近苗圃购进的2年生珙桐幼苗。2014年3月中旬将2年生珙桐幼苗移栽到口径23 cm、深21 cm的塑料花盆中, 土壤为潮褐土, 每盆栽种1株苗, 并定期进行浇水、喷药、除草等工作。待幼苗自然生长1 y后, 于2015年5月, 选取生长一致、无病虫害珙桐幼苗, 分别设置处理组(2个遮阴强度)和对照组。其中, 对照组为自然光(Y0, 透光率100%); 处理组1: 施加一层黑色遮阴网, 用数字照度计测得透光率约为62%(Y1); 处理组 2: 施加两层遮阴网, 测得透光率约为 43%(Y2)。遮阴网的高度距地面1.8 m。每个光照梯度10株幼苗,共30株, 按照常规技术管理, 试验处理110 d后, 测定不同光照环境下幼苗的光合特性及生长特性。
2.2 测定方法
2.2.1 叶片光合参数的测定
选取珙桐幼苗完全伸展的顶层叶片, 采用美国Li-Cor公司生产的Li-6400便携式光合仪, 选择配备红蓝光源叶室, 于2015年9月7日上午8: 00—12: 00测定光响应曲线, 通过光合仪的自动记录功能在光合有效辐射(PAR) 0—1600 μmol·m-2·s-1内进行连续测定, 光照强度梯度为 1600、1400、1200、1000、800、600、400、200、100、50、0 μmol·m-2·s-1, 记录珙桐幼苗的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(gs)和胞间 CO2浓度(Ci)等并计算气孔限制值(Ls), 每个处理测定3盆幼苗, 测量幼苗中上部发育健康的叶片, 每盆测定3—5片叶。于9月8日8: 00—18: 00测定光合日变化, 每隔2小时测定一次, 共测定6次, 即8: 00、10: 00、12: 00、14: 00、16: 00、18: 00, 每次测定6个叶片, 取平均值。
测定条件: 叶室温度控制在 20—25°C, 光照强度为600 μmol·m-2·s-1, CO2浓度为 400 μmol·mol-1, 流速为 500 μmol·s-1。测定公式为:

其中,Ls为气孔限制值,Ci为胞间CO2浓度,Ca代表气室中 CO2浓度为 400 μmol·mol-1

其中,WUE为水分利用效率,Pn为净光合速率,Tr为蒸腾速率
利用非直角双曲线模型对光响应曲线进行拟合, 其表达式为:

式中,Pn为叶片净光合速率,α为表观量子效率(AQY),Pmax为最大净光合速率,I为光合有效辐射,θ为曲线的凸度, 凸度越大, 说明曲线的弯曲程度越大,Rd为暗呼吸速率。并通过计算得到光饱和点(LSP)和光补偿点(LCP)。
叶片的瞬时光能利用效率(LUE)可用公式算出,

2.2.2 幼苗生长参数测定
9月9日, 各处理随机选取幼苗中上部生长良好的叶片15—20片, 用游标卡尺分别测量叶片的叶长(Leaf Length,LL)和叶宽(Leaf Width,LW), 取平均值。用Epson Perfection V700 Photo扫描仪进行叶面积的扫描, 每个处理扫描 15片, 然后用 Winfolia 2007进行叶面积的计算。每个处理随机选5株幼苗,将其小心挖出并洗净, 将根、茎、叶分离后按单株袋装, 带回实验室于80 ℃烘干至恒重, 用电子天平称量。求出如下参数: 叶长/叶宽(LL/LW), 比叶面积(specific leaf area,SLA, 总叶面积/总叶重), 总生物量(biomass), 叶生物量(leaf mass)、茎生物量(Stem mass)、根生物量(root mass)、根冠比(root to shoot ratio,R/S, 根生物量/地上部分生物量)。
2.3 数据处理
采用SPSS18.0和Excel 2010软件对数据进行统计分析并制表, 采用 one-way ANOVO分析不同光强下珙桐幼苗各参数的差异显著性。
3 结果与分析
3.1 遮阴对珙桐幼苗光合特性的影响
3.1.1 遮阴对珙桐幼苗叶片光合日变化的影响
由图1可知, 全光照下珙桐幼苗的净光合速率(Pn)日变化曲线呈“双峰型”, 即在10: 00和14: 00时出现峰值, 分别为 4.62 和 4.45 μmol·m-2·s-1; 而遮阴处理下均呈“单峰型”。Y1和Y2处理下,Pn峰值均出现在 10: 00, 分别为 5.73 和 6.42 μmol·m-2·s-1。3 种光照处理下, 蒸腾速率(Tr)日变化均在12: 00达到全天最高值, 分别为 1.45、2.05 和 2.72 mmol·m-2·s-1; 气孔导度(gs)日变化大体呈下降趋势, 胞间 CO2浓度(Ci)日变化均先下降后上升, 但出现低谷的时间不同: 在Y0处理下,Ci出现低谷的时间为10: 00, 其值为89.54 μmol·mol-1, 而在 YI和Y2 处理下, 出现低谷的时间为 12: 00, 分别为 60.20 和 102.53 μmol·mol-1。
3.1.2 遮阴对珙桐幼苗叶片光响应曲线的影响
自然光处理的珙桐幼苗最大光合速率(Pmax)、表观量子效率(α)及光饱和点(LSP)均低于遮阴处理的幼苗, 而光补偿点(LCP)和暗呼吸速率(Rd)则大于遮阴处理的珙桐幼苗(表1)。全光照条件下, 珙桐幼苗光补偿点LCP和Rd均最高, 分别为 120.64 μmol·m-2·s-1和 1.24 μmol·m-2·s-1; 而全光照条件下, 最大光合速率和表观量子效率最低, 分别为 2.48 μmol·m-2·s-1和0.01。Y1和Y2处理下, 珙桐幼苗叶片LCP下降至40.85和28.09, 但最大净光合速率较高, 分别为3.28和3.96, 表观量子效率均为0.02。

图1 不同遮阴处理下珙桐幼苗光合特征日变化差异Fig.1 Diurnal variation of photosynthesis of D.involucrata seedlings in different shading treatment
不同遮阴处理下Pn均高于全光照, Y2处理下Pn最高(图 2)。Pn随PAR增加而上升, 当PAR在 0400 μmol·m-2·s-1,Pn上升速度最快; 当PAR达到 400 μmol·m-2·s-1之后,Pn上升速度减缓;PAR>800 μmol·m-2·s-1,Pn值逐渐稳定。三种光环境下光能利用效率(LUE)的光响应过程基本相似, 均为明显的单峰曲线(图4)。不同遮阴处理下WUE均高于全光照, Y2处理下WUE最高(图3)。WUE随PAR增加而上升, 当PAR在 0—400 μmol·m-2·s-1,WUE上升速度最快;PAR>800 μmol·m-2·s-1,WUE值波动较为稳定。LUE随PAR增加先增大后减小, 当PAR< 200 μmol·m-2·s-1,LUE上升速度最快, 对光强响应敏感;在 100—200 μmol·m-2·s-1光强范围内, 达到峰值; 此后LUE逐渐下降, 当PAR>800 μmol·m-2·s-1,LUE变化较小。在Y0处理下LUE达到峰值后先迅速下降,之后又有所回升,PAR>400 μmol·m-2·s-1,LUE变化逐渐稳定(图4)。光强较弱时, 遮阴处理的幼苗LUE大于自然光处理, 表明遮阴的珙桐幼苗能高效地利用弱光环境。
3.1.3 光合特征各指标相关性分析
由表2可知, 珙桐幼苗叶片Pn与gs和WUE呈正相关, 且与WUE呈显著正相关, 与Ci呈显著负相关。其中Ci与Pn相关系数最高为–0.95, 说明Pn受Ci影响最大。珙桐幼苗叶片Ci与gs、WUE呈负相关, 与Tr呈正相关,gs与WUE呈负相关。
3.2 遮阴对珙桐幼苗生长特性的影响
3.2.1 珙桐幼苗叶性状的影响
遮阴对珙桐幼苗叶长、叶宽、比叶面积等有显著影响(表3)。叶长、叶宽、比叶面积均随着遮阴程度的增大而增加, Y2处理的珙桐幼苗叶长、叶宽显著高于 Y0处理(p<0.05), 与 Y1差异不显著。其中, 叶长是Y0处理的1.25倍, Y1处理的1.13倍; 叶宽是Y0处理的1.09倍, Y1处理的1.24倍。Y2处理下, 幼苗比叶面积显著高于 Y0和Y1(p<0.05), 分别是Y0处理和Y1处理的2.42倍和2.24倍。
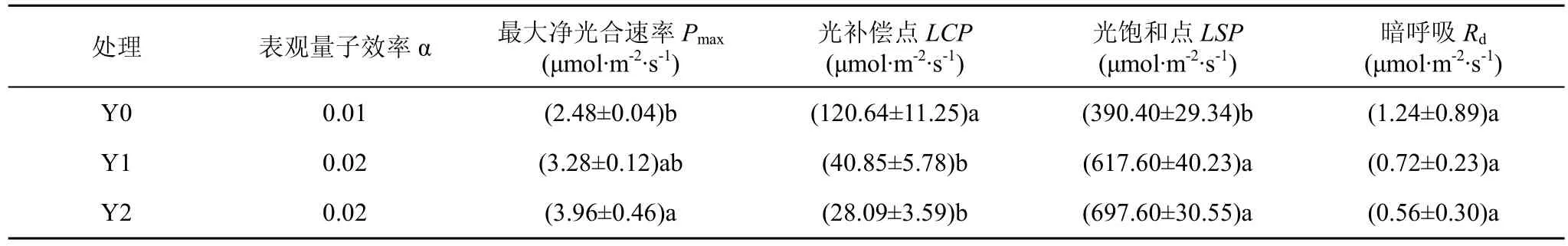
表1 遮阴处理对珙桐叶片光合作用特征参数的影响Tab.1 Effect of shading treatments on the photosynthetic characteristic parameters of D.involucrate seedlings
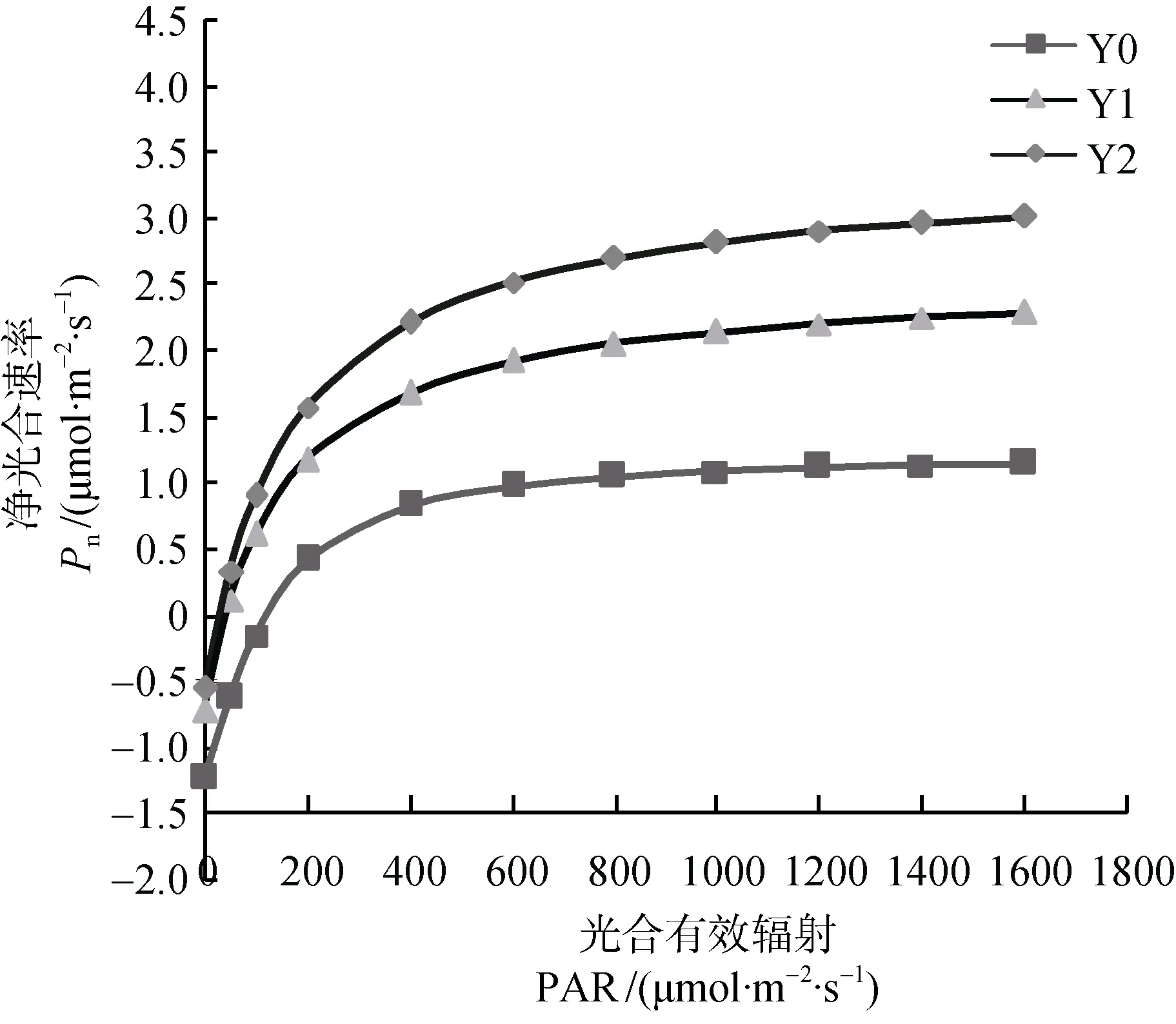
图2 不同遮阴处理下珙桐幼苗光合-光响应曲线Fig.2 Net photosynthetic rate-light response curves of D.involucrata seedlings in different shading treatments

图3 不同遮阴处理下珙桐幼苗水分利用效率光响应和光能利用效率光响应Fig.3 Water use efficiency-light response curves of D.involucrata seedlings in different shading treatments

图4 不同遮阴处理下珙桐幼苗光能利用效率光响应Fig.4 Light use efficiency-light response curves of D.involucrata seedlings in different shading treatments

表2 珙桐幼苗叶片光合作用生理指标的相关性系数Tab.2 Correlation coefficients of photosynthesis physiological indexes of D.involucrata seedlings

表3 遮阴处理对珙桐叶片性状的影响Tab.3 Effect of shading treatments on the leaf characteristics of D.involucrate seedlings
3.2.2 遮阴对珙桐幼苗生物量的影响
光照强度显著影响珙桐幼苗生物量积累及生物量在植株各部位的分配(表4)。随光照强度的减弱,珙桐幼苗的根、茎、叶生物量及根冠比均呈现先增加后降低的趋势, 且在全光照条件下珙桐幼苗的总生物量、根、茎、叶生物量均低于遮阴处理(表4)。Y1处理下珙桐幼苗的根、叶生物量显著高于Y0和Y2(p<0.05), 根冠比显著高于 Y2(p<0.05), 与对照(Y0)无差异。Y2处理下, 幼苗的根生物量和叶生物量与对照(Y0)基本一致, 根冠比则明显小于对照(Y0)处理。

表4 遮阴处理对珙桐幼苗生物量的影响Tab.4 Effect of shading treatments on the biomass of D.involucrate seedlings
4 讨论
4.1 光照对珙桐幼苗光合特性的影响
光合日变化能反映植物对特定光照环境的适应情况。全光照下, 珙桐幼苗叶片Pn日变化呈双峰曲线(图1), 说明珙桐幼苗有明显的“午休现象”。出现光合午休现象可能有两种原因: 一是随着光强和温度升高, 叶片的气孔关闭, 进入叶肉细胞的 CO2数量减少, 而使胞间CO2浓度(Ci)上升, 最终导致光合作用下降; 二是高光照可能引起 PSⅡ结构的破坏,降低RuBisCO酶的活性, 进而导致叶片净光合速率下降[9]。本研究中全光照下gs日变化呈下降趋势,Ci日变化先下降后上升(图1), 说明产生午休现象的原因更可能是受到气孔限制, 与姚毅等[10]的研究结果吻合。而在遮阴处理下,Pn日变化均呈单峰曲线, 且遮阴处理的净光合速率(Pn)均高于全光照, 说明遮阴有利于珙桐幼苗叶片气孔开放和 CO2的利用, 提高光合速率。此外, 宋天宇等[11]和韩素菊[8]研究发现25%全光照条件下珙桐幼苗Pn显著低于自然光下的幼苗, 说明过度遮阴会使珙桐幼苗光合作用受到限制, 选择适宜的光照条件更有利于幼苗生长。研究还发现, 不同遮阴处理下1年生珙桐幼苗Pn日变化曲线均为双峰型[14], 与本研究结果有出入, 这很可能与珙桐幼苗在不同的年龄阶段, 光合能力的大小存在差异有关, 姚毅等[10]研究结果也证实了这一点,发现 1年生珙桐的光能利用率较低, 臧润国等[12]对天山云杉的研究也发现不同龄级林木的Pn日变化有差异, 所以在引种时可考虑栽种树龄较大的幼苗,以提高光能利用效率。
光响应曲线特征对了解植物叶片的光化学效率有重要意义。最大光合速率(Pmax)也称为光合能力,大多数研究都认为, 生长在高光照环境中的植物具有较高的最大光合速率, 研究发现, 自然光处理的黄波罗幼苗Pmax高于遮阴处理[13], 闽楠幼树Pmax随光强的提高而增大[2]。而本实验呈现的结果与此相反, 遮阴处理的最大光合速率均高于全光照(表1),说明遮阴提高了叶片的光能利用效率(LUE), 张云等[14]对堇叶紫金牛的研究也有类似结果。光饱和点(LSP)和光补偿点可以反映植物叶片对光的利用能力[15-16], 遮阴处理下, 珙桐幼苗光补偿点(LCP)的降低有利于有机物的积累, 提高幼苗对弱光环境的耐受力, 同时较低的暗呼吸速率(Rd)可减少有机物的消耗, 是幼苗对弱光的一种适应性调节, 这与王宁宁[17]的研究结果一致, 但是过低的光照却会抑制珙桐幼苗的生长。
4.2 光照对珙桐幼苗叶性状和生物量的影响
植物叶片是对环境变化最敏感的器官, 叶片功能性状与植物碳的同化以及对资源的利用能力关系密切, 它可以反映植物适应环境变化形成的适应策略[18]。叶长宽比(LL/LW)及比叶面积(SLA)可以反映植物对特定环境的适应性和对资源的综合利用率[19-20]。弱光下, 植物通过增加叶片比叶面积(SLA)来增强光合能力[21-22]。对东北次生林主要树种的研究结果表明, 所有树种比叶面积均随光照强度的降低而增加[23]。本研究中Y2处理的比叶面积(SLA)显著高于Y0(表3), 也证实了这一结果。且Y2叶长宽比(LL/LW)也高于Y0(表3),LL/LW的增加, 表明叶片逐渐变长, 光合产物被用于伸展叶面积, 使得SLA增大, 更好地适应弱光环境。研究发现, 60%全光照处理增大了树莓的比叶面积, 提高了光合效率;而 40%全光照处理造成光照严重不足, 光合能力下降[3]。本实验中, 43%并未造成光照严重不足, 反而光合能力更强, 说明珙桐幼苗对弱光的适应性高于树莓, 在43%全光照环境下依然可以正常生长。
珙桐幼苗在遮阴处理下能产生更多的生物量,Y1、Y2总生物量均高于Y0, 且Y1根生物量和叶生物量显著高于 Y0(表4), 这表明低光照有利于珙桐幼苗生物量的积累, 具有较强耐阴性的红松(Pinus koraiensis)幼苗在适度遮阴条件也呈现这一规律[24]。生物量分配模式的改变是植物能在不同光照环境下生存的重要原因[25]。一般情况下, 光照条件受到限制时, 植物通过改变自身生物量的分配适应弱光环境, 将更多的资源分配到地上部分以获取更多的光照资源, 但也有研究表明, 化香(Platycarya longipes)幼苗倾向于把更多的生物量分配给叶片, 而麻栎(Quercus acutissima)则是分配给根部[26]。可见, 弱光环境下植物的生物量分配策略并不是单一的。根冠比(R/S)可以较准确反映植物地上部和地下部生物量的分配关系。Y1根冠比(R/S)与 Y0无差异, 但 Y2根冠比(R/S)却显著低于Y0(表4), 所以, 遮阴更有利于珙桐幼苗将更多的资源分配到地上部分, 一方面增强叶片对光能的捕获, 另一方面有利于珙桐幼苗支持结构和运输结构的构建。对蒙古栎(Quercus mongolica)的研究结果[27]以及对刺槐(Robinia pseudoacacia)的研究结果[28]均支持这一观点。幼苗在遮阴条件下生物量增加足以说明珙桐幼苗耐阴性较强, 但重度遮阴却不适宜幼苗的生长, 如宋天宇等[11]和王宁宁[17]分别研究了透光率为 25%和 5%—10%环境下幼苗的生长状况, 均证实过分遮阴不利于幼苗生长。因此, 了解幼苗叶片对不同光环境的响应, 对研究珙桐群落演替和生态恢复有很重要的现实意义。
综上所述, 珙桐幼苗在全光照条件下有明显的“光合午休”现象, 遮阴处理能够缓解“光合午休”现象。遮阴处理下珙桐幼苗的Pmax、AQY、WUE及LUE均高于全光照, 珙桐幼苗通过降低LCP、Rd的光合策略来适应光强的降低。遮阴后珙桐叶片在形态和光能利用效率方面均表现出对弱光环境的驯化适应,所以在珙桐幼苗期可以选择遮阴的环境以保证其正常生长; 在用作园林植物时, 可选择有隐蔽的地方,如种植在高大建筑物旁。另外, 本研究中没有设置43%全光照以下的光强, 也没有对不同龄级珙桐幼苗耐阴能力进行比较, 是本试验方案设计的缺陷所在, 今后的研究应考虑设置低于 43%全光照的光照处理和不同龄级耐阴性的对比, 以更加深入揭示珙桐幼苗对不同强度光照的响应。
[1]SALGADO-LUARTE C, GINAOLI E.Herbivory may modify functional responses to shade in seedlings of a light-demanding tree species[J].Functional Ecology, 2011 25(3): 492–499.
[2]王振兴, 朱锦懋, 王健, 等.闽楠幼树光合特性及生物量分配对光环境的响应[J].生态学报, 2012, 32(12):3841–3848.
[3]王晓冬, 贺国强, 赵利群,等.遮阴对树莓光合特性和果实性状的影响[J].植物研究, 2014, 34(5): 599–603.
[4]禹玉婷, 徐刚标, 汪晓萍.珙桐研究进展[J].经济林研究, 2006, 24(4): 92–94.
[5]胡进耀, 苏智先, 黎云祥.珙桐生物学研究进展[J].中国野生植物资源, 2003, 22(4): 15–19.
[6]张家勋, 李俊清.珙桐的天然分布和人共同引种分析[J].北京林业大学学报, 1995, 17(1): 25–30.
[7]司继跃, 雷妮娅, 等.珙桐研究综述[J].科学技术与工程, 2009, 13(9), 3713–3719.
[8]韩素菊.遮荫对珙桐幼苗光合特性及生长特征的影响研究[J].四川林业科技, 2014, 05(05): 45–48.
[9]朱巧玲, 冷佳奕, 叶庆生.黑毛石斛和长距石斛的光合特性[J].植物学报, 2013, 48(2): 151–159.
[10]姚毅, 刘艳红, 胡耀升, 等.珙桐一年生苗光合特性的研究[J].生态科学, 2012, 31 (4): 384–389.
[11]宋天宇, 刘艳红, 姚毅.土壤氮素与光照对盆栽珙桐幼苗光合参数的影响[J].生态科学, 2011, 30 (3): 243–249.
[12]臧润国, 刘华, 张新平, 等.天山中部天然林分中不同龄级天山云杉光合特性[J].林业科学, 2009, 45 (5): 60–68.
[13]王凯, 朱教君, 于立忠, 等.遮阴对黄波罗幼苗的光合特性及光能利用效率的影响[J].植物生态学报, 2009, 33(5):1003–1012.
[14]张云, 夏国华, 马凯, 等.遮阴对堇叶紫金牛光合特性和叶绿素荧光参数的影响[J].应用生态学报, 2014, 25(7):1940–1948.
[15]张旺峰, 樊大勇, 谢宗强, 等.濒危植物银杉幼树对生长光强的季节性光合响应[J].生物多样性, 2005, 13(5):387–397.
[16]秦舒浩, 李玲玲.遮光处理对西葫芦幼苗形态特征及光合生理特性的影响[J].应用生态学报, 2006, 17(4): 653–656.
[17]王宁宁.珙桐苗木光合特性对干旱、光照强度和二氧化碳浓度的响应[D].北京: 北京林业大学, 2010.
[18]MCINTYRE S, LAVOREL S, LANSBERG J, et al.Disturbance response in vegetation: towards a global perspective on functional traits[J].Journal of Vegetation Science, 1999, 10(5): 621–630.
[19]WILSON P J, THOMPSON K, HODGSON J G.Specific leaf area and leaf dry matter content as alternative predictors of plant strategies[J].New Phytologist, 2010,143(1): 155– 162.
[20]WRIGHT I J, WESTOBY M, REICH P B.Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span [J].Journal of Ecology, 2002, 90(3): 534–543.
[21]SU J K, YU D J, KIM T C, et al.Growth and photosynthetic characteristics of blueberry (Vaccinium corymbosum cv.Bluecrop) under various shade levels[J].Scientia Horticulturae, 2011, 129(3): 486–492.
[22]LUSK C H.Leaf area and growth of juvenile temperate evergreens in low light: species of contrasting shade tolerance change rank during ontogeny[J].Functional Ecology, 2004, 18(6): 820–828.
[23]李明财, 朱教君, 孙一荣.东北次生林主要树种比叶面积对光照强度的响应[J].生态学杂志, 2009, 28(8):1437–1442.
[24]吕晋慧, 王玄, 冯雁梦, 等.遮荫对金莲花光合特性和叶片解剖特征的影响[J].生态学报, 2012, 32(19): 6033–6043.
[25]金鑫, 胡万良, 丁磊, 等.遮阴对红松幼苗生长及光合特性的影响[J].东北林业大学学报, 2009, 37(9): 12–13.
[26]MONTGOMERY R.Relative importance of photosynthetic physiology and biomass allocation for tree seedling growth across a broad light gradient[J].Tree Physiology, 2004,24(2): 155–167.
[27]杨莹, 王传华, 刘艳红.光照对鄂东南2种落叶阔叶树种幼苗生长、光合特性和生物量分配的影响[J].生态学报,2010, 30 (22): 6082–6090.
[28]许中旗, 黄选瑞, 徐成立, 等.光照条件对蒙古栎幼苗生长及形态特征的影响[J].生态学报, 2009, 29(3): 1121–1128.
[29]徐飞, 郭卫华, 徐伟红, 等.不同光环境对麻栎和刺槐幼苗生长和光合特征的影响[J].生态学报, 2010, 30(12):3098–3107.
Effects of light intensity on the photosynthesis and growth characteristics ofDavidia involucrata seedlings
JIANG Ruifang, LIU Yanhong*
College of Forestry of Beijing Forestry University, 100083,China
Davidia involucrataBaill is a rare and endangered plant and has been listed as first degree national protective plant.In order to understand the light adaptability ofD.involucrate, the effects of different light intensities (sunlight, 62% sunlight and 43% sunlight) on leaf photosynthetic characteristics and biomass allocation ofD.involucratewere studied.The net photosynthetic rate (Pn) ofD.involucratevaried as a bimodal curve with a ‘midday depression’ phenomenon in full bright light,and as a unimodal curve in the shading treatments.With shade, plants had higher photosynthetic rate (Pmax), apparent quantum yield(AQY), water use efficiency(WUE) and light use efficiency(LUE), lower light-compensation point and dark respiration rate.Plants grown in the shade treatment had greaterSLAand biomass, lowerR/T.We concluded thatD.involucratehad the ability to adjust to shade in leaf photosynthetic and growth characteristics.Therefore,D.involucrateseedlings are shade tolerant.
Davidia involucrataBaill; light intensity; photosynthetic characteristics; growth characteristics
10.14108/j.cnki.1008-8873.2017.05.015
Q948.11
A
1008-8873(2017)05-114-07
姜瑞芳, 刘艳红.光照对珙桐幼苗光合与生长特性的影响[J].生态科学, 2017, 36(5): 114-120.
JIANG Ruifang, LIU Yanhong.Effects of light intensity on the photosynthesis and growth characteristics ofDavidia involucrataseedlings[J].Ecological Science, 2017, 36(5): 114-120.
2016-08-16;
2016-10-20
北京市共建重点学科项目“生态学”(201306-201406);国家重点研发项目:极小种群野生植物回归技术研究与示范(2016YFC0503106)
姜瑞芳, (1990—), 女, 硕士研究生, 主要从事生物多样性方面研究, E-mail: ecojrf@163.com
*通信作者:刘艳红(1965—), 女, 教授, 博士生导师, 主要从事森林生态学, 生物多样性保护方面研究, E-mail: liuyh@bjfu.edu.cn
