土豆抗性淀粉的体外消化与发酵
2017-11-06李晓文刘瑞兴易翠平
谢 涛,祝 红,李晓文,刘瑞兴,易翠平,*
1. 湖南工程学院化学化工学院 (湘潭 411104)2. 长沙理工大学化学与生物工程学院 (长沙 410015)
2017-07-03
谢 涛,男,1970年出生,教授,主要从事再生资源与食品、生物化工研究。
*通讯作者:易翠平,女,1973年出生,教授,主要从事粮食深加工研究。
土豆抗性淀粉的体外消化与发酵
谢 涛1,祝 红2,李晓文2,刘瑞兴2,易翠平2,*
1. 湖南工程学院化学化工学院 (湘潭 411104)2. 长沙理工大学化学与生物工程学院 (长沙 410015)
研究了土豆回生淀粉(RS3)和交联淀粉(RS4)的体外消化动力学与体外人粪分批厌氧发酵。结果表明:土豆回生淀粉(RS3)和交联淀粉(RS4)中缓慢消化淀粉(SDS)与抗性淀粉(RS)的含量显著增加,两者属于低血糖指数(pGI)食物。以土豆回生淀粉(RS3)和交联淀粉(RS4)为唯一碳源发酵时,活的厌氧菌总数和双歧杆菌数都比原淀粉、Novelose 230(RS2)的要高,但乳酸杆菌数都出现不同程度的下降。四种淀粉样品都能促进短链脂肪酸(SCFAs)总量的增加,其中土豆回生淀粉(RS3)、交联淀粉(RS4)与Novelose 230(RS2)特别有利于促进丁酸的生成。土豆回生淀粉(RS3)与交联淀粉(RS4)能够改善人体肠道菌群的分布、促进SCFAs特别是丁酸的生成,具有开发成为新的益生素的潜力。
土豆;回生淀粉(RS3);交联淀粉(RS4);体外消化与发酵
根据生物可利用性,淀粉可分为快速消化淀粉(RDS)、缓慢消化淀粉(SDS)和抗性淀粉(RS)三类[1]。RDS在小肠内快速消化后引起血糖水平急剧上升,SDS在小肠内完全但缓慢消化,RS在小肠中不能消化。RS和SDS具有许多独特功能,已引起了国内外广大研究者的极大兴趣和广泛关注,成为食品科学界、动物营养学界的研究热点[2-6]。根据RS的形态及理化性质,又分为RS1、RS2、RS3和RS4,能大量制备的是后2种[7-9]。RS在人体大肠内被不同的微生物发酵,涉及各种代谢过程,包括厌氧微生物降解有机质产生代谢能用于菌体生长,引起微生物菌群的变化[10]。物理与化学改性淀粉已被广泛应用于食品工业中,其中有些具有酶抗性。RS3、RS4能阻止淀粉颗粒的溶胀,提高其对热、剪切力和pH值的稳定性,具有相同的生理功能。RS主要的末端代谢产物是短链脂肪酸(SCFAs)、气体(CO2, CH4, H2)和热量。SCFAs对宿主能产生重要的生理作用,如降低胞内pH值,促进矿物质吸收,减少结肠内次级代谢产物胆汗酸的生成,以及影响大肠内微生物菌群分布等[11-14]。在SCFAs中,丁酸是大肠细胞优先的能量来源,能抑制肠癌细胞的扩散[11]。有关RS促进SCFAs生产与改善微生物菌群分布的研究始终不断,RS作为唯一碳源对丁酸的生成有很强的影响[15-16]。
当今,国人的日常饮食已从传统的富碳水化合物向富蛋白质转变,肉和奶制品的消费急剧增加,肠癌发病率也快速增加[17]。众所周知,某些癌的发病率与膳食的改变直接相关。许多研究已聚焦在胃肠疾病的防治方面,特别是功能性淀粉资源的开发利用[10,13,15]。本文旨在以Novelose 240(RS2)为对照,分析土豆回生淀粉(RS3)和交联淀粉(RS4)的体外消化动力学行为,并以其为唯一碳源,研究体外人粪厌氧发酵中肠道菌群的变化与SCFAs的生产。
1 材料与方法
1.1材料与试剂
淀粉:土豆淀粉购自四川友嘉食品有限公司,Novelose 240(RS2)购自Sigma-Aldrich公司。
营养培养基:9.240 g NaHCO3、7.125 g Na2HPO4·12H2O、0.470 g NaCl、 0.450 g KCl、0.100 g Na2SO4、0.055 g CaCl2、0.047 g MgCl2和0.400 g 尿素溶解于990 mL碳酸盐缓冲液中,并与10 mL微量元素溶液(含36.8 mg FeSO4·7H2O、19.00 mg MnSO4·7H2O、4.40 mg ZnSO4·7H2O、1.20 mg CoCl2·6H2O、1.98 mg CuSO4·5H2O和0.17 mg Mo7(NH4)6O24·4H2O)混匀。
分离培养基:TP选择性培养基(Bifidobacteria sp)、BL非选择性培养基(总厌氧菌)和LBS选择性培养基(Lactobacilli)购自美国Life Tech公司。
酶制剂:淀粉转葡萄苷酶(400 AGU,Novozymes)、猪胰液素(P7545; Sigma-Aldrich)、转化酶(200 000 SU/g; Novozymes)。
其他试剂均为分析纯。
1.2土豆回生淀粉(RS3)的制备
参照文献[18-19]的方法。用蒸馏水配制30%的淀粉乳液,调节pH为6.0,沸水浴30 min后,110 ℃湿热处理40 min。冷却,4 ℃放置24 h,80 ℃烘干,粉碎过120目筛得回生抗性淀粉(RS3)。
1.3土豆交联淀粉(RS4)的制备
称取1 kg土豆淀粉溶解在1 500 mL水中制成淀粉浆,再加入100 g硫酸钠溶解混匀。淀粉浆用10%(w/v)HCl溶液调至pH 2,加热至50 ℃振荡维持10 h。冷至室温,用5%(w/v)NaOH溶液调至pH 10。再加入100 g三聚磷酸钠(STPP)/三偏磷酸钠(STMP)(99∶1)混合物,于50 ℃反应10 h,用稀HCl溶液调至pH 5,过滤、洗涤,干燥至水分含量12%(w/w),粉碎过120目筛得交联抗性淀粉(RS4)。
1.4体外消化及动力学分析
参照文献[20-21]的方法,略做改动。称取50 mg重结晶淀粉粉末放入15 mL离心管,用2.5 mL含0.02%叠氮钠、pH 5.2的盐酸-醋酸钠缓冲液混匀,预热至37℃。将8 g猪胰α-淀粉酶溶于10 mL上述缓冲液中,于37 ℃孵育10 min,1 500 g离心得到的上清液即为α-淀粉酶溶液。使用前在α-淀粉酶溶液中混入125 μL(300 U/mL)淀粉葡萄糖酶即得混合酶液。5份200 μL混合酶液与100 μL淀粉乳液的混合液配制后,立即置于37 ℃往复式振荡水浴(170次/min)中依次反应0 min、20 min、60 min、120 min和180 min,加入96%的乙醇溶液900 μL钝化酶活性并使之沉淀。沉淀在4 ℃贮放1 h,4 ℃离心(5 000 g)10 min。上清液中的还原糖含量采用葡萄糖氧化酶/过氧化物酶(GOPOD)试剂盒测定。
快速消化淀粉(RDS)、缓慢消化淀粉(SDS)和抗性淀粉含量由0 min(G0)、20 min(G20)、120 min(G120)时的葡萄糖含量以及样品的初始干重(S)按以下公式计算而得:%RDS=(G20-G0)×0.9×100/S,%SDS=(G120-G20)×0.9×100/S,%RS=1-(%RDS+%SDS) 。
动力学方程C=C∞×(1-e-kt)可用于描述淀粉的酶解过程,C指还原糖含量,C∞指180 min时的还原糖浓度,k是水解速率常数,t是水解时间(h),以完全糊化的冷冻干燥淀粉为对照。相对消化率(Dr)由某一时间待测样水解曲线积分面积除以对照样水解曲线积分面积再乘100计算而来。预期血糖指数pGI= 8.198 + 0.862×Dr计算。
1.5体外发酵过程
1.5.1混合酶液配制
2.8 mL淀粉转葡萄苷酶(400 AGU,Novozymes)用蒸馏水稀释至8 mL(酶活力为140 AGU/mL)。4只离心管中每只放入1.8 g猪胰液素(P7545; Sigma-Aldrich),分别用20 mL蒸馏水悬浮混匀,离心管振动10 min后于1 500×g离心10 min。从每只离心管中取出13.5 mL上清液合并得54 mL,再与6 mL稀淀粉转葡萄苷酶溶液、4 mL转化酶(200 000 SU/g; Novozymes)混合。
1.5.2酶解消化
称取1 g淀粉样品放入50 mL带盖试管中,加入0.1 mol/L的醋酸盐缓冲液(pH 5.2)20 mL混合。将带盖试管放入沸水浴中振荡糊化10 min,取出于冰浴中冷却至37 ℃,放入5粒玻璃珠(Φ 5 mm; Sigma-Aldrich),再移入37 ℃水浴中平衡20 min。加入5 mL混合酶液,带盖试管置于水平振荡摇床(振幅35 mm,160次/min)上消化6 h。之后移入85 ℃水浴中维持15 min钝化酶。在带盖试管中加入95%的乙醇300 mL沉淀未消化淀粉,离心收集未消化淀粉,干燥粉碎过120目筛用于厌氧发酵。
1.5.3厌氧发酵
参照文献[22]的方法,略做改动。取年龄23~31岁健康男子(此前至少3个月以上未注射抗生素,没有预先服用已知的益生元或益生菌,没有胃肠病史)的粪便,贮存于充满CO2的气瓶中。粪便按1∶5(w∶v)用营养培养基混合均匀,并用6层医用纱布过滤。称取100 mg混合酶液消化的样品(唯一碳源)放入15 mL带盖试管中,加入10 mL粪便滤液。同时制作一份加入粪便滤液的空白(100 mg未加淀粉样品的混合酶液)。在37 ℃水浴中鼓入CO2以维持发酵所需的厌氧环境。为使接种物分布均匀,每12 h振动带盖试管20 s。
1.5.4活细菌计数
用厌氧稀释液将种子培养液稀释至合适的浓度,接种每个选择性培养基,然后在好氧或厌氧条件下培养。培养36 h,每12 h活菌计数1次,以l g(CFU/mL)表示。
1.5.5短链脂肪酸(SCFA)分析
参照文献[12]的方法测定,SCFA浓度以mmol/L表示,总SCFAs含量以乙酸、丙酸与丁酸的总和表示。
所有数据为3个平行实验的平均值,且采用SPSS 20.0进行显著性分析。
2 结果与讨论
2.1体外消化动力学
土豆淀粉体外消化的动力学分析结果见表1。与土豆原淀粉相比,它经湿热回生处理与磷酸化交联反应后则有了很大变化,快速消化淀粉(RDS)含量分别减少了79.66%和93.86%,缓慢消化淀粉(SDS)分别增加了3.95和4.63倍,尤其是抗性淀粉(RS)分别增加了4.48倍和5.29倍。与土豆原淀粉比,由于其回生淀粉与交联淀粉中功能性成分SDS、RS[2-6]急剧增加,因而相对消化率(Dr)急剧下降,血糖指数(pGI)大大减小。从pGI值来看,土豆原淀粉属典型的高pGI食物(pGI>75),而3种抗性淀粉则属于低pGI食物(pGI<55),适合于糖尿病、肥胖症类病人食用[23-24]。
2.2肠道菌群的变化
由表2可见,空白对照由于未加淀粉作为碳源,厌氧菌总数在前24 h变化不大,至36 h时则显著下降,而以土豆原淀粉、RS2、RS3与RS4作为唯一碳源时,厌氧菌总数在第24 h时都达到最大(比空白对照有显著增加),然后厌氧菌总数减少可能是因淀粉基质和其它营养物质被消耗所致。双歧杆菌和乳酸杆菌活细胞计数结果见图1。由图1(a)可看出,在发酵到第12 h时,以RS3与RS4为唯一碳源时,双歧杆菌的增殖最快,其次为RS2, 最后是原淀粉与空白对照;第12 h后双歧杆菌数量变化出现相反的结果,以RS3与RS4为唯一碳源时, 双歧杆菌数量稍有增加,而后基本保持不变,其余3种情况则持续下降,特别是RS2在36 h后降至最低。由图1(b)也可看出,以RS3与RS4为唯一碳源时,乳酸杆菌数量在第12 h时稍有增加而后基本保持不变,发酵到第36 h时,出现较快下降;而以土豆原淀粉与RS2为唯一碳源时,乳酸杆菌数量的变化规律基本与之相似,只是变化的速度稍快些。这些结果与Wang等[16]研究一致。大多数乳酸杆菌与双歧杆菌不能利用淀粉,因为它们不能产生淀粉水解酶类。然而,RS2、RS3及RS4的未消化部分是如何促进肠道菌群生长的,到目前为止尚不知其原因[22]。人粪培养物是复杂的,存在许多相互关联且相互作用的不同菌株,或许另外一些细菌可以降解RS2、RS3与RS4的未消化部分,并足够支持乳酸杆菌和双歧杆菌的生长。
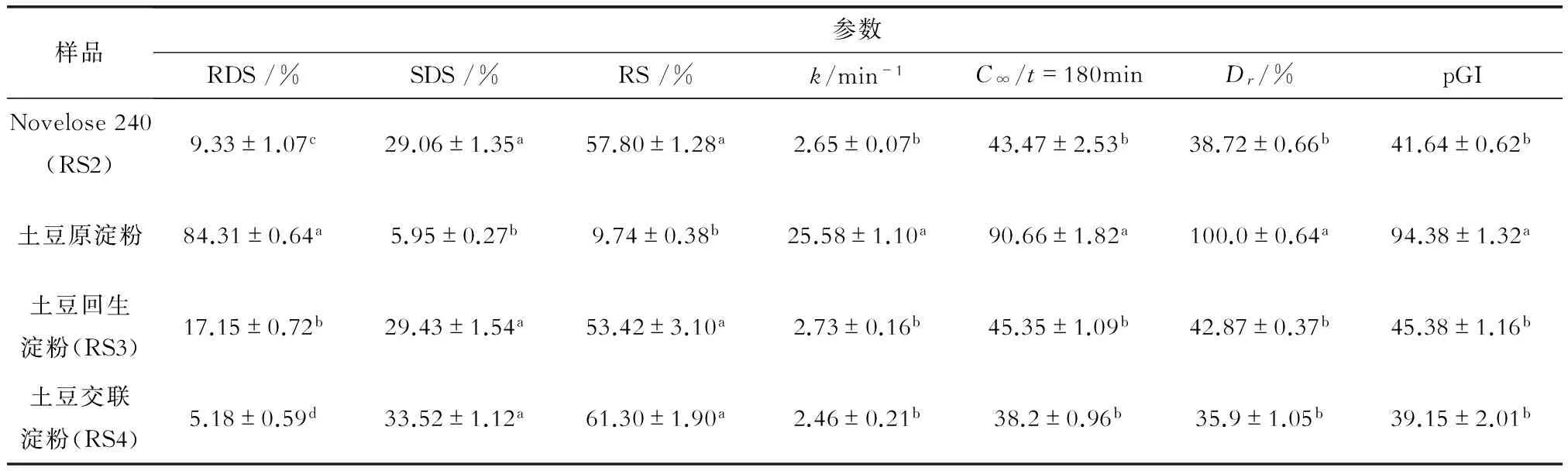
表1 磷酸化交联淀粉的体外消化与血糖指数
注:不同字母表示存在显著差别(p<0.01),相同字母表示差别不显著(p>0.05),下同。
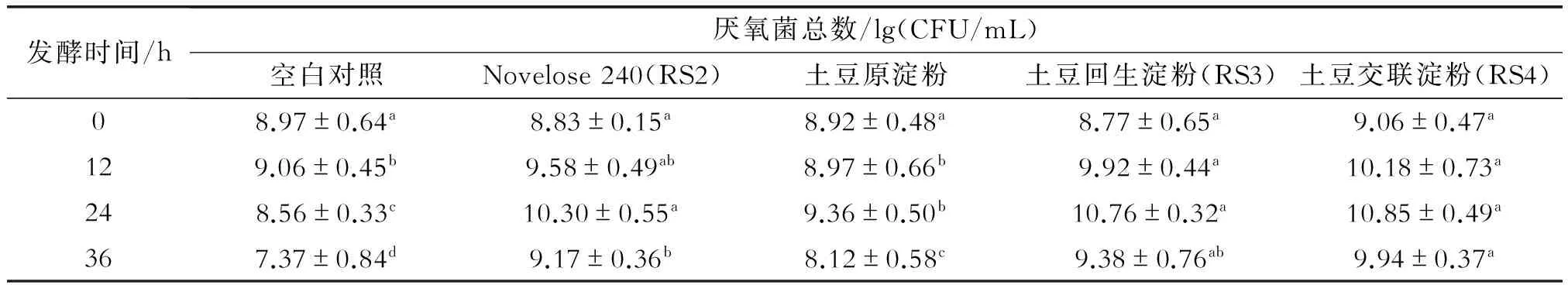
表2 大便接种物分批发酵过程中总厌氧细菌数量的变化
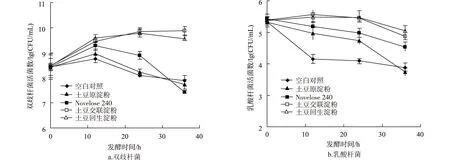
图1 大便接种物分批发酵过程中双歧杆菌与乳酸杆菌计数的变化
2.3短链脂肪酸的产生
除空白对照在整个发酵过程中总SCFAs浓度很低且产量没有增加外,以其它样品为唯一碳源时SCFAs的生成均有持续增加(图2)。在发酵36 h后,以RS2、RS3与RS4为唯一碳源时,乙酸/丙酸/丁酸的摩尔比分别为41/12/26、36/22/21、38/21/20,总SCFAs产量分别为79.38 mmol/L、78.16 mmol/L、78.33 mmol/L,而土豆原淀粉的乙酸/丙酸/丁酸摩尔比也达到34/27/11、总SCFAs产量为69.03 mmol/L(表3)。与空白对照以生产乙酸为主比较,以其它样品为唯一碳源时生产乙酸的比例显著下降约22%左右,但生产丁酸的比例却明显增加,除土豆原淀粉的稍低(16.83%)外,RS2、RS3与RS4分别达到31.82%、28.78%和27.43%,说明它们是有利于促进丁酸的生成[1-2,10-11]。

图2 大便接种物分批发酵过程中总SCFAs的变化
3 讨论
许多文献[20-24]报道:当pGI<55时为低pGI食物,pGI在55~75时为中等pGI食物,pGI>75时为高pGI食物。高pGI的食物进入胃肠后消化快、吸收率高,葡萄糖释放快,葡萄糖进入血液后峰值高;低pGI食物在胃肠中停留时间长,吸收率低,葡萄糖释放慢,葡萄糖进入血液后的峰值低,下降速度慢。本研究中,土豆原淀粉经变性处理后,土豆回生淀粉(RS3)和交联淀粉(RS4)中SDS、RS的含量明显增加,两者与Novelose 240(RS2)都属于低pGI食物,而土豆原淀粉属高pGI食物。
目前,以淀粉和非淀粉等多聚糖为唯一碳源进行体外人粪培养物厌氧发酵的报道已有许多[1,13,16,22],而很少有回生淀粉(RS3)和交联淀粉(RS4)在这方面的研究。回生淀粉(RS3)和交联淀粉(RS4)结构牢固,抗溶胀与抗糊化能力强,因而比原淀粉更能抗酶解消化。但对其抗消化性的说法仍很不统一,有人认为大多数RS在24 h内被完全发酵利用[25],有人报道只有85%[26],甚至仅有60%[14]。本研究中,尽管RS与原淀粉之间的消化率有些不同,但3种RS样品在24 h内90%以上已被消化(数据未列出),而其消化率不同可能与基质来源及浓度、接种物浓度及用量、稀释倍数、培养基、样品预处理条件等因素有关[15]。随着发酵继续,菌种总量减少,可能是由于淀粉基质的消耗、SCFAs的积累所导致。
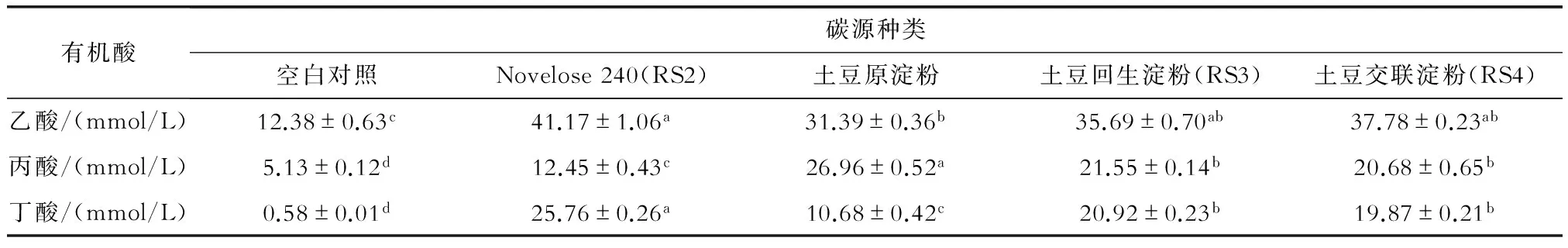
表3 大便接种物分批发酵36 h后短链脂肪酸的摩尔浓度
在所有样品中,土豆回生淀粉(RS3)和交联淀粉(RS4)均表现出比Novelose 240(RS2)要高的总厌氧细菌水平,而且与其它样品相比双歧杆菌数量也因两者有很大增加,双歧杆菌是RS3、RS4发酵最主要的菌株,这与已有研究一致[16,25,27]。在本研究中,土豆回生淀粉(RS3)和交联淀粉(RS4)都比其它样品对乳酸杆菌的生长更有效。双歧杆菌和乳酸杆菌是益生菌,因为它们能增强人体对病原体感染的抵抗力,产生酶和维生素,调整宿主免疫反应[28]。例如,只有双歧杆菌和丁酸梭菌能有效利用高直链玉米淀粉,在含有高直链玉米淀粉的培养基中具有很高的比生长速率,分别能降解40%和80%的直链淀粉[27]。
淀粉或碳水化合物分子中抗酶解的部分最终在人体大肠内被发酵和代谢成SCFAs。Wyatt等[26]报道,以玉米RS3为唯一碳源的人粪体外厌氧发酵产生的SCFAs中,乙酸/丙酸/丁酸的摩尔比高达80/9/8。Schwiertz等[12]报道,豌豆RS3能引起总SCFAs的增加,尤其是排泄物中丁酸浓度显著增高。本研究以土豆回生淀粉(RS3)与交联淀粉(RS4)为唯一碳源进行体外人粪分批厌氧发酵36 h后,乙酸/丙酸/丁酸的摩尔比分别约为36/22/21、38/21/20,丁酸能稳定生成,而且与Novelose 240(RS2)为唯一碳源时SCFAs的生产差别不大,这是因为所有样品都用混合酶液消化处理,且使用相同的数量用于发酵。
总之,土豆回生淀粉(RS3)与交联淀粉(RS4)属于低pGI食物,能促进双歧杆菌、乳酸杆菌的生长和提高SCFAs的生产。与Novelose 240(RS2)一样,它们可能给人体结肠产生有益的影响,具有开发成为新益生因子的前景。
[1] Englyst H N, Kingman S M, Cummings J H. Classification and measurement of nutritionally important starch fractions [J]. European Journal of Clinical Nutrition [J], 1992, 46: 33-50.
[2] Fuentes Aaragozae A C, Riquelme Navarrete M J, Sánchez Zapata E, et al. Resistant starch as functional ingredient: A review [J]. Food Research International, 2010, 43: 931-942.
[3] Rodríguez Cabezas M E, Camuesco D, Arribas B, et al. The combination of fructoologosaccharides and resistant starch shows prebiotic additive effects in rats[J]. Clinical Nutrition, 2010, 29: 832-839.
[4] 何 健, 王 韧, 张 昊, 等. 改性蜡质玉米淀粉抗消化组分的理化性质及分子结构表征[J]. 食品与机械, 2016, 32(5): 1-4.
[5] Zhang G, Hamaker B R. Slowly digestible starch: Concept, mechanism, and proposed extended glycemic index [J]. Critical Reviews in Food Science and Nutrition, 2009, 49(10): 73-85.
[6] Lee C J, Kim Y. Slowly digestible starch from heat-moisture treated waxy potato starch: preparation, structural characteristics, and glucose response in mice [J]. Food Chemistry, 2012, 133(4): 1222-1229.
[7] Lehmann U, Jacobasch G, Schmiedl D. Characterization of resistant starch type III from banana [J]. Journal of Agricultural and Food Chemistry, 2002, 50: 5236-5240.
[8] Mutungi C, Rost F, Onyango C, et al. Crystallinity, thermal and morphological characteristics of resistant starch type Ⅲ produced by hydrothermal treatment of debranched cassava starch[J]. Starch/Stärke, 2009, 61: 634-645.
[9] Pongjanta J, Utaipattanaceep A, Naivikul O. Debranching enzyme concentration effected on physicochemical properties and α-amylase hydrolysis rate of resistant starch type III from amylase rice starch[J]. Carbohydrate Polymers, 2009, 78: 5-9.
[10] Higgins J A, Brown I L. Resistant starch: A promising dietary agent for the prevention/treatment of inflammatory bowel disease and bowel cancer [J]. Curr Opin Gastroenterol, 2013, 29: 190-194.
[11] Topping D L, Clifton P M. Short chain fatty acids and human colonic function: Roles of resistant starch and non-starch polysaccharides [J]. Physiol Rev, 2001, 81: 1031-1064.
[12] Schwiertz A, Lehmann U, Jacobasch G, et al. Influence of resistant starch on the SCFA production and cell counts of butyrate producing Eubacterium spp. in the human intestine [J]. J Appl Microbiol, 2002, 93: 157-162.
[13] Hylla S, Gostner A, Dusel G, et al. Effect of resistant starch on the colon in healthy volunteers: possible implications for cancer prevention [J]. Am J Clin Nutr, 1998, 67: 136-142.
[14] Edward C A, Gibson G, Champ M, et al. In vitro method for quantification of the fermentation of starch by human faecal bacteria [J]. J Sci Food Agr,1996, 71: 209-217.
[15] Le Bay G, Michel C, Blottiere H M, et al. Enhancement of butyrate production in the rat caecocolonic tract by long-term ingestion of resistant potato starch [J]. Brit J Nutr, 1999, 82: 419-426.
[16] Wang X, Brown I L, Khaled D, et al. Manipulation of colonic bacteria and volatile fatty acid production by dietary high amylose maize (amylomaize) starch granules [J]. J Appl Microbiol, 2002, 93: 390-397.
[17] Kim D S, Kang S W. Mortality and potential years of life lost of cololectal cancer between Korea and OECD countries before and after the year 2000 [J]. J Korea Acad Industr Coop Soc, 2012, 13: 5261-5270.
[18] 曾红华, 谢 涛, 杨 莉. 几种薯类与豆类抗性淀粉的的抗消化性及其益生效应[J]. 中国粮油学报, 2012, 27(11): 30-34.
[19] 谢 涛, 张淑远. 重结晶红薯淀粉体外消化前后益生作用与结构变化[J]. 农业机械学报, 2013, 44 (8): 203-208.
[20] Mutungi C, Onyango C, Rost F, et al. Structural and physicochemical properties and in vitro digestibility of recrystallized linear α-D-(1→4) glucans derived from mild-acid modified cassava starch[J]. Food Research International, 2010, 43(4): 1144-1154.
[21] Miao M, Jiang B, Jin Z Y, et al. Impact of mild acid hydrolysis on structure and digestion properties of waxy maize starch[J]. Food Chemistry, 2011, 126: 506-513.
[22] Bae C H, Park M S, Ji G E, et al. Effects of phosphorylated cross-linked resistant corn starch on the intestinal microflora and short chain fatty acid forming during in vitro human fecal batch culture [J]. Food Sci Biotechnol, 2013, 22(6): 1649-1654.
[23] Lehmann U, Robin F. Slowly digestible starch—its structure and health implications: A review[J]. Trends in Food Science and Technology, 2007, 18(7): 346-355.
[24] Zhang G, Hamaker B R. Slowly digestible starch: concept, mechanism, and proposed extended glycemic index[J]. Critical Reviews in Food Science and Nutrition, 2009, 49(10): 73-85.
[25] Englyst H N, Macfarlane G T. Breakdown of resistant starch and readily digestible starch by human gut bacteria [J]. J Sci Food Agr, 1986, 37: 699-706.
[26] Wyatt G M, Horn N. Fermentation of resistant food starches by human and rat intestine bacteria [J]. J Sci Food Agr, 1988, 44: 281-288.
[27] Wang X, Conway P L, Brown I L. In vitro utilization of amylopectin and high-amylose maize (amylomaize) starch granules by human colonic bacteria [J]. Appl Environ Microbiol, 1999, 65: 4848-4854.
[28] Takahashi T, Nakagawa E, Nara T, et al. Effects of orally ingested Bifidobacteria longum on the mucosal IgA response of mice to dietary antigens [J]. Biosci Biotechnol Biochem, 1998, 62: 10-15.
Theinvitrodigestibilityandfermentationofpotatoresistantstarch
Xie Tao1, Zhu Hong2, Li Xiaowen2, Liu Ruixing2, Yi Cuiping2,*
1. College of Chemical Engineering, Hunan Institute of Engineering (Xiangtan 411104 ) 2. College of Chemistry and Biology Science, Changsha University of Science & Technology (Changsha 410015)
Theinvitrodigestible kinetics and human fecal batch fermentation of retrograded starch (RS3 type) and cross-linked starch (RS4 type) from potato native starches were focused on. The results show that there are higher quantities of slowly digestible starch (SDS) in potato RS3 and higher quantities of resistant starch (RS) in potato RS4. Both potato RS3 and RS4 are low glycemic index (pGI) foods. When RS3 or RS4 is provided as the sole carbon source in aninvitrohuman fecal batch fermentation experiment, the viable counts of total anaerobic bacteria and bifidobacteria are higher for the RS3 and RS4 starches than for RS2 starch (Novelose 240). However, the viable counts of lactobacilli are lower for the RS3 and RS4 starches than for RS2 starch. The total short chain fatty acids (SCFAs) content increases during the fermentation analysis of all four starch samples. The production of butyric acid is favored in medium containing RS3 or RS4 as the carbon source, compared with that containing RS2. These results show that potato RS3 and RS4 can improve human intestinal flora profiles and promote SCFAs formation, especially butyric acid. Thus, these starches have the potential for development as new probiotics.
potato; retrogradated starch (RS3); cross-linked starch (RS4);invitrodigestibility and fermentation
TS235.2
A
1672-5026(2017)05-035-06
