华北地区猪繁殖与呼吸综合征病毒变异分析
2017-10-23焦文强白献晓徐引弟王克领郎利敏朱文豪李海利张青娴张立宪王治方
焦文强,白献晓,徐引弟,王克领,郎利敏,朱文豪,李海利,张青娴,张立宪,游 一,王治方,许 峰
(河南省农业科学院 畜牧兽医研究所,河南 郑州 450002)
华北地区猪繁殖与呼吸综合征病毒变异分析
焦文强,白献晓*,徐引弟,王克领,郎利敏,朱文豪,李海利,张青娴,张立宪,游 一,王治方,许 峰
(河南省农业科学院 畜牧兽医研究所,河南 郑州 450002)
为了解华北地区猪繁殖与呼吸综合征病毒(PRRSV)的遗传变异情况,通过对2014—2016年从河南、河北、山东、山西4省采集的疑似猪繁殖与呼吸综合征(PRRS)病料进行检测,将扩增得到的PRRS阳性样品的ORF5基因进行测序,分析华北地区2014—2016年PRRSV的变异情况。结果表明,2014—2016年华北地区流行毒株与国内外代表性毒株核苷酸同源性在84.6%~99.8%;抗原位点比对结果显示,变异株中和表位第37—44位氨基酸为SH(I/L/F)QLIY(N/K),诱骗表位27—30位氨基酸为(V/A)L(V/A)N;系统进化树分析表明,以CH-1a为代表的变异株、以VR-2332为代表的经典毒株以及NADC30毒株在华北地区同时存在。可见,华北地区PRRSV流行情况复杂。
猪繁殖与呼吸综合征病毒; ORF5; 序列分析; 进化
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种以母猪流产、早产,仔猪发生呼吸道疾病为特征的烈性传染病[1-4]。该病于1987年首次在美国报道,之后迅速蔓延至世界上大多数养猪国家[5]。我国学者郭宝清于1996年首次从病料中分离到PRRSV,证实了PRRS在我国的存在[6]。2006年,我国大多数养猪省份暴发了以母猪高烧、流产为主要特征的高致病性蓝耳病(HP-PRRS),并一直延续至今,严重威胁着我国养猪业的健康发展,其病原被认定为PRRSV变异株[7-8]。
PRRSV属于动脉炎病毒科(Arterivirus)动脉炎病毒属(Arteriviridae) 成员,分为2个基因型:以VR-2332株为代表的北美洲型和以LV株为代表的欧洲型。PRRSV基因组为单股正链RNA,长度约为15 kb,含9个开放阅读框 (ORFs),编码2个非结构蛋白(NSPs) 和7个结构蛋白(SP)。其中,ORF5编码的GP5蛋白是变异最大的结构蛋白,成为分析PRRSV遗传变异和进化的首选对象[9-10]。接种疫苗是目前防控PRRS最有效的办法,但PRRSV在免疫压力下发生变异,规避中和抗体的中和作用。PRRSV整个生命周期中缺乏真核生物中复制酶5′—3′的校正作用,使得PRRSV基因组存在较大可能的错配现象。此外,PRRSV普遍存在缺失、插入以及重组现象[11-13],以及由于某种不确定因素导致病毒生命周期意外终止而产生大量的准种,使得PRRSV同源重组、变异、突变情况变得异常复杂[14-15]。同时PRRSV不同毒株间的交叉保护力非常差,且疫苗研发相对于病毒变异往往处于滞后的状态。因此,针对PRRSV主要结构蛋白,如GP5进行遗传变异检测就显得十分重要。
为深入了解华北地区PRRSV遗传变异情况,对2014年11月至2016年8月从河南、河北、山西、山东规模化猪场采集的样品进行检测,扩增PRRSV ORF5基因并进行测序分析,旨在为有效指导临床PRRS的防控工作提供参考。
1 材料和方法
1.1供试病料
于2014年11月至2016年8月共采集河南、山西、河北、山东等地区疑似病料553份,包括肺脏、淋巴结、血液等。
1.2主要试剂
TaqDNA聚合酶、鼠源反转录酶(M-MLV)、RNA酶抑制剂、pMD18-T Simple载体、DL2000 Marker均购自大连宝生物工程有限公司;Trizol试剂购自Invitrogen公司,琼脂糖购自Oxoid公司,其他试剂均为国产分析纯。
1.3引物设计
参考GenBank中已发表的PRRSV毒株ORF5序列的保守区序列,应用Primer Premier 5.0软件设计引物,ORF5-F:5′-CCATTCTGGTGGCAATTTGA-3′, ORF5-R:5′-GGCATATATCATCACTGGCG-3′。引物由生工生物工程(上海)股份有限公司合成。
1.4PRRSVORF5基因的克隆与测序
按照Trizol试剂操作说明书提取疑似样品的RNA,随即合成cDNA,反应条件为42 ℃ 1.5 h,70 ℃ 10 min。以cDNA为模板进行PCR扩增,反应程序为:94 ℃预变性5 min;94 ℃ 30 s,57 ℃ 30 s,72 ℃ 1 min,35个循环;最后72 ℃延伸10 min。扩增结束后,取5 μL PCR产物于1.0%琼脂糖凝胶中电泳检测扩增结果,回收与预期大小一致的PCR产物,并与pMD18-T Simple载体连接,4 ℃连接过夜后转化至JM109感受态细胞,蓝白斑筛选后挑取单克隆进行培养并提取质粒,质粒送生工生物工程(上海)股份有限公司进行测序。使用Meg Align软件将本研究得到的PRRSV序列与GenBank中收录的PRRSV序列进行核苷酸和氨基酸的比对,使用MEGA 5.0软件构建系统进化树。
2 结果与分析
2.1PRRSVORF5基因的扩增结果
从图1可以看出,PCR扩增PRRSV ORF5可得到750 bp大小的片段,符合预期大小。553份待检样品中,有127份样品检测为PRRSV阳性,阳性率高达22.97%。

M为DL 2000 Marker;1、2为待检样品图1 PRRSV ORF5扩增结果
2.2PRRSVORF5基因序列比对
本研究得到的16株(HEB-HD、HENAN-JIAOZ、HENAN-XINX、HENAN-XINX3、HEN-AY、HEN-AY2、HEN-KF、HEN-NY2、HEN-NY、HEN-PDS、HEN-PY、HEN-XC、HEN-ZHUMD、HEN-ZK、SHANDONG-DM、SHANXI-JINC)PRRSV 毒株均为北美洲型,与国内外分离毒株核苷酸同源性在84.6%~99.8%,与NADC30毒株核苷同源性在84.7%~96.4%(图2)。
2.3PRRSVORF5抗原位点比对结果
从图3可以看出,ORF5的突变主要集中在第3~17位氨基酸、第23~39位氨基酸、第121~128位氨基酸、第161~170位氨基酸。其中,位于第37—44位氨基酸的中和表位,第39位氨基酸I大多突变为L或者F,第44位氨基酸存在N→K的突变,其氨基酸为SH(I/F/L)QLIY(N/K);位于第27—30位氨基酸的诱骗表位,其氨基酸为V(A)LA(V)N;第13位氨基酸中存在R→Q的突变。
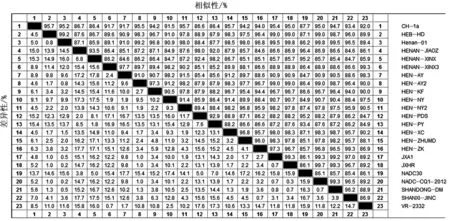
图2 ORF5核苷酸序列比对结果

图3 PRRSV ORF5主要抗原位点比对结果
2.4基于PRRSVORF5构建的系统进化树
由图4可知,本研究得到的毒株均为北美洲型毒株,且HEN-AY、HEN-NY、HENAN-XINX3与VR-2332毒株在一个分支,表明HEN-AY、HEN-NY、HENAN-XINX3属于经典毒株;HEN-PDS、HEN-PY、HENAN-JIAOZ、HENAN-XINX与NADC30毒株在一个分支,表明HEN-PDS、HEN-PY、HENAN-JIAOZ、HENAN-XINX为PRRSV NADC30毒株;其他毒株与CH-1a毒株在一个分支,均为高致病毒株。以CH-1a为代表的变异株、以VR-2332为代表的经典毒株以及NADC30毒株同时存在,表明华北地区PRRSV流行情况日益复杂。
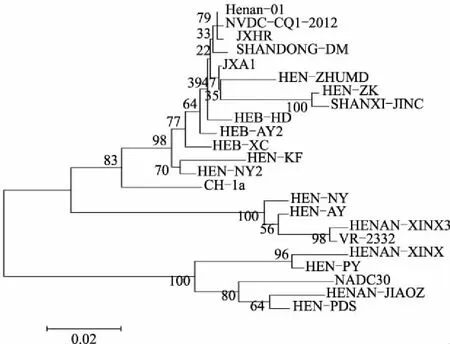
图4 基于ORF5构建的系统进化树
3 结论与讨论
将本研究得到的PRRSV ORF5核苷酸序列与国内外毒株的序列进行比对,结果显示,核苷酸同源性在84.6%~99.8%,分离到的流行毒株均为北美洲型毒株,且不同区域间的差异较大。R13被认为与PRRSV的毒力存在某种潜在的联系,抗原位点的比对结果发现,分离株第13位氨基酸发生了R→Q的突变,在NADC30毒株中也存在同样的突变,该位点的突变与周峰等[16]的研究相一致。
位于第37—44位氨基酸的中和性抗原位点目前已经确定存在于北美洲型PRRSV毒株中,中和抗体的识别位点是第38、42、43、44位氨基酸,而第39、40、41位氨基酸被认为是结合位点[17]。本研究结果显示,分离株的中和抗体识别位点第44位氨基酸存在N→K突变,而中和抗体结合位点的第39位氨基酸存在I→L突变以及I→F突变。第27—30位氨基酸的诱骗表位可以推迟中和抗体的产生,其氨基酸为V(A)LA(V)N,这些氨基酸的突变对于表位的影响还有待于进一步研究[18]。这些位点的突变可能是PRRSV在疫苗压力下产生的应变,也可能是病毒自身随机重组产生,其突变可能导致中和抗体不能有效识别和结合PRRSV,从而导致免疫失败。关于R13突变对于病毒毒力的影响,需要进一步研究。
系统进化树分析显示,HEN-AY、HEN-NY、HENAN-XINX3与VR-2332毒株处于一个分支,HEN-PDS、HEN-PY、HENAN-JIAOZ、HENAN-XINX与NADC30毒株处于同一个分支,其他毒株均为PRRSV高致病毒株。综合上述情况,经典毒株与高致病毒株同时存在,加之新近流行毒株NADC30的出现[19],使得华北地区PRRSV流行情况日益复杂,PRRS的防控更加困难。针对目前这种复杂的情况,单一的针对经典毒株、高致病毒株或者NADC30毒株的疫苗很难达到令人满意的防控效果。采用基因工程方法,开发针对多种毒株、不同抗原位点的新型疫苗显得极为迫切。
[1] 司朝朝,史西保,李恭贺,等.猪繁殖与呼吸障碍综合征病毒非结构蛋白nsp1α的表达及活性检测[J].河南农业科学,2015,44(1):126-129.
[2] 赵孟孟,冯丽丽,张二芹,等.猪繁殖与呼吸综合征病毒NSP9蛋白的亚细胞定位与功能预测[J].河南农业科学,2016,45(12):138-142.
[3] 侯婕,李睿,马红芳,等.猪繁殖与呼吸综合征病毒入侵受体唾液酸黏附素的表达与纯化[J].河南农业科学,2016,45(3):130-134.
[4] 孟帆,姚敬明,吴忻,等.猪群暴发猪瘟与蓝耳病混合感染的诊断与防制[J].山西农业科学,2010,38(6):66-68.
[5] Keffaber K K.Reproductive failure of unknown etiology[J].Am Assoc Swine Pract Newsl,1989,1:1-9.
[6] 郭宝清,陈章水,刘文兴,等.从疑似PRRS流产胎儿分离PRRSV的研究[J].中国畜禽传染病,1996(2):1-4.
[7] Tian K G,Yu X L,Zhao T Z,etal.Emergence of fatal PRRSV variants:Unparalleled outbreaks of a typical PRRS in China and molecular dissection of the unique hallmark[J].PLoS One,2007,2(6):e526.
[8] Zhou L,Yang H C.Porcine reproductive and respiratory syndrome in China[J].Virus Res,2010,154:31-37.
[9] Key K F,Haqshenas G,Guenette D K,etal.Genetic variation and phylogenetic analysis of the ORF5 gene of acute porcine reproductive and respiratory syndrome virus isolated[J].Vet Microbiol,2001,83(3):249-263.
[10] Chen J,Liu T,Zhu C G,etal.Genetic variation of Chinese PRRSV strains bases on ORF5 sequence [J].Biochem Genet,2006,44(9/10):425-435.
[11] Goldberg T L,Hahn E C,Weigel R M,etal.Genetic geographical and temporal variation of porcine reproductive and respiratory syndrome virus in Illinois[J].J Gen Vi-rol,2000,81: 171-179.
[12] Meng X J.Heterogeneity of porcine reproductive and respiratory syndrome virus:Implications for current vaccine efficacy and future vaccine development[J].Vet Microbiol,2007,74(4): 309-329.
[13] 高志强,郭鑫,杨汉春,等.猪繁殖与呼吸综合征病毒缺失变异株的基因组特征[J].畜牧兽医学报,2005,36(6):578-584.
[14] Stadejek T,Stankevicius A,Storggaard T,etal.Identification of radically different variants of porcine reproductive and respiratory syndrome virus in Eastern Europe: Towards a common ancestor for European and American virus[J].J Gen Virol,2002,83:1861-1873.
[15] 周海范,夏平安,崔保安,等.猪繁殖与呼吸综合征病毒河南分离株ORF5基因的克隆与变异分析[J].中国兽医学报,2007,27(6):810-813.
[16] 周峰,常洪涛,赵军,等.2012—2013年猪繁殖与呼吸综合征病毒河南流行株的分离鉴定及分子流行病学调查[J].中国兽医学报,2014,34(9):1398-1404,1410.
[17] 王金凤.河北省2007—2011年猪繁殖与呼吸综合征病毒的分子流行病学调查[D].保定:河北农业大学,2011.
[18] 韩一超,刘文俊,姚敬明,等.猪繁殖与呼吸综合征病毒Shanxi-6株全基因序列分析[J].华北农学报,2014,29(2):62-65.
[19] 王林建,郭振华,乔松林,等.河南地区NADC30-like PRRSV毒株的增殖特性与遗传进化分析[J].河南农业科学,2017,46(3):122-128.
Variation Analysis of Porcine Reproductive and Respiratory Syndrome Virus Isolates in Northern China
JIAO Wenqiang,BAI Xianxiao*,XU Yindi,WANG Keling,LANG Limin,ZHU Wenhao,LI Haili, ZHANG Qingxian,ZHANG Lixian,YOU Yi,WANG Zhifang,XU Feng
(Institute For Animal Husbandary and Veterinary Research,Henan Academy of Agricultural Sciences,Zhengzhou 450002,China)
In order to elucidate the evolution of the porcine reproductive and respiratory virus(PRRSV) in northern China from 2014 to 2016,suspicious samples collected from Henan,Hebei,Shandong and Shanxi province were subject to PRRSV diagnosis,the positive samples were further subject to RT-PCR to obtain the sequence of ORF5.The sequence acquired were further aligned with the ORF5 sequence available in GenBank to construct phylogenetic tree.Also,antigenic sites of ORF5 compared to those of other isolates to analyse whether the antigenic sites has changed or not.Sequence comparision showed that the homology of ORF5 gene obtained in the present study was up to 84.6%—99.8%.Antigenic site comparison showed that neutralizing epitipes of the HP-PRRSV located at 37—44 were SH(I/L/F)QLIY(N/K),decoy epitope located at 27—30 were (V/A)L(V/A)N.Phylogenetic tree showed that HP-PRRSV with the representive strain CH-1a,together with classic PRRSV represented by VR-2332 and NADC30 strain have been circulating in northern China.In conclusion,the PRRSV has been increasingly complicated.
porcine reproductive and respiratory syndromevirus(PRRSV); ORF5; sequence analysis;evolution
S855.3
A
1004-3268(2017)10-0128-04
2017-04-28
河南省科技攻关项目 (162102110050,162102110042);河南省农业科学院自主创新基金项目(2016ZC51,2017ZC49)
焦文强(1982-),男,河南新乡人,助理研究员,博士,主要从事分子病毒学方面研究。E-mail:wenqiangjiao@163.com
*通讯作者:白献晓(1963-),男,河南南阳人,研究员,主要从事动物传染病诊断方面的研究。E-mail:1030410210@qq.com
