AANAT基因编码区多态性与家兔生长性状的关联分析
2017-10-23刘卜玮陈仕毅贾先波赖松家
刘卜玮,王 杰,陈仕毅,贾先波,赖松家
(四川农业大学动物遗传育种研究所,四川成都 611130)
AANAT基因编码区多态性与家兔生长性状的关联分析
刘卜玮,王 杰,陈仕毅,贾先波,赖松家*
(四川农业大学动物遗传育种研究所,四川成都 611130)
为研究五羟色胺-N-乙酰基转移酶(AANAT)基因编码区(CDS区)多态性和家兔生长性状的关联性,以天府黑兔、伊拉兔和新西兰兔作为实验材料,对AANAT基因的外显子进行PCR扩增,直接测序检测家兔AANAT基因外显子的单核苷酸多态性。结果表明:在3个兔品种的CDS区上均发现有5个完全连锁突变位点,分别是C.132 T>C、C.153T>C、C.159 A>T、C.234 A>G、C.291 A>G,形成了野生型TTTTAAAAAA、杂合型TCTCATAGAG、突变型CCCCTTGGGG 3种基因型,突变型新西兰兔在56、70、84日龄的体重显著高于野生型和杂合型(P<0.05),而突变型伊拉兔和天府黑兔在70、84日龄的体重显著高于杂合型和野生型(P<0.05)。综上可知,AANAT基因可作为肉兔生长性状的候选基因。
家兔;AANAT;单核苷酸基因多态性;生长性状
我国是养兔大国,也是兔肉出口大国。家兔是一种非常重要的经济动物,它既可以食用,也可以作为工业原料、观赏动物、医用模型等。五羟色胺-N-乙酰基转移酶(AANAT)是褪黑激素(Melatonin,MT)合成过程的限速酶[1]。MT在生物体内具有调控动物的生长、应激、炎症、昼夜节律、视网膜信号以及发情泌乳等重要功能[2-4]。近些年来研究发现,在AANAT基因mRNA的5'UTR上面有内部核糖体进入位点(IRES),昼夜节律的调节依靠IRES的活性,在夜间,核内不均一核糖核蛋白Q(hnRNP Q)作用于IRES,使其活性显著增加,影响AANAT基因转录以及MT的合成[5]。目前,AANAT基因在家畜上面的研究比较少,但是作为MT的限速酶,其对家畜的生产可能起着重要作用。本研究将对家兔AANAT基因的编码区单(CDS区)核苷酸多态性和兔的生长性状进行关联性分析,从而为家兔的分子育种工作提供指导。
1 材料与方法
1.1 实验材料 选取四川农业大学育种场的天府黑兔122只,成都金堂育种场的伊拉兔124只,新西兰兔281只作为实验材料。在90日龄对家兔进行耳组织采样,样品放在含有75%酒精的1.5 mL离心管中,带回实验室,-80℃保存。
1.2 实验方法
1.2.1 耳组织DNA的提取 将耳组织样从-80℃冰箱中取出并解冻,从离心管中取出耳组织并用无菌滤纸擦干,用眼科剪剪去耳组织表面的兔毛,然后剪碎耳组织放入新的EP管中备用;使用TIANGEN公司的TIAN amp Genomic DNA Kit 基因组DNA提取试剂盒按照说明书步骤提取耳组织的DNA,放置在-20℃冰箱备用。
1.2.2 目的基因的扩增 使用Primer Premier 6根据NCBI网站家兔AANAT基因(GeneID:100357535)外显子序列设计引物,再由成都擎科生物公司合成引物,引物序列如表1所示。
确定引物退火温度,PCR反应体系采用10 μL体系:Mix 5 μL,ddH2O 3.2 μL,上、下游引物各0.4 μL,DNA 1 μL。PCR反应程序:95℃ 5 min,95℃30 s,55~63℃ 30 s,72℃ 1 min,39个扩增循环,72℃10 min,4℃保存。然后将PCR产物进行凝胶电泳检测,选取最亮条带对应的温度为引物的退火温度。
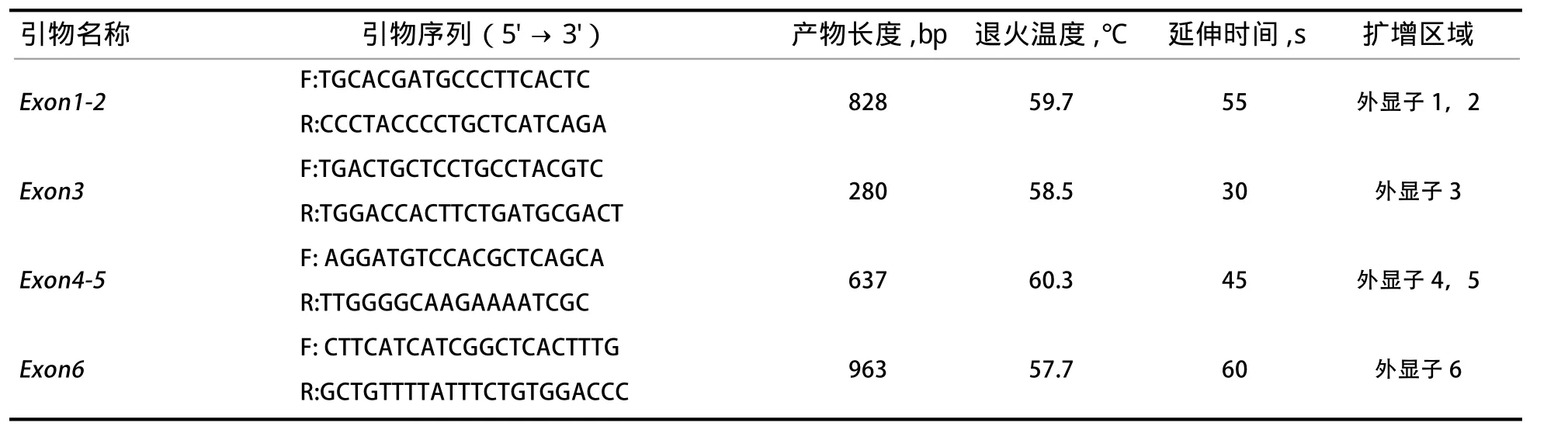
表1 AANAT基因引物序列
混管测序,按照各个引物的退火温度进行PCR反应,将PCR产物进行电泳检测,每对引物选取16个条带明亮的PCR产物,每个PCR产物取4 μL,每4个PCR产物混成一管送至成都擎科生物公司进行测序。
分型,混管测序后,发现第1、2、3外显子无突变,4、5、6外显子上面有突变,把527个DNA样品进行PCR反应,并送至成都擎科生物有限公司进行测序。
2 结果与分析
2.1 序列分析 使用Seqman软件查看测序结果,并和NCBI上AANAT基因序列做比较,混管测序发现,AANAT基因外显子1、2、3上面没有发现突变位点,在外显子4、5、6的CDS区发现5个同义突变位点;再对527个PCR产物进行单管测序,经统计表明,CDS区的5个突变位点完全连锁;利用codonw计算RSCU(同义密码子相对使用度),结果见表2和图1。
2.2 对AANAT基因类型和家兔生长性状的关联性分析 由表3可知,突变型新西兰兔在56、70、84日龄的体重显著高于野生型和杂合型(P<0.05),而突变型伊拉兔和天府黑兔在70、84日龄的体重显著高于杂合型和野生型(P<0.05)。各个突变位点的多态信息含量(PIC)达到中度多态(0.25
3 讨 论
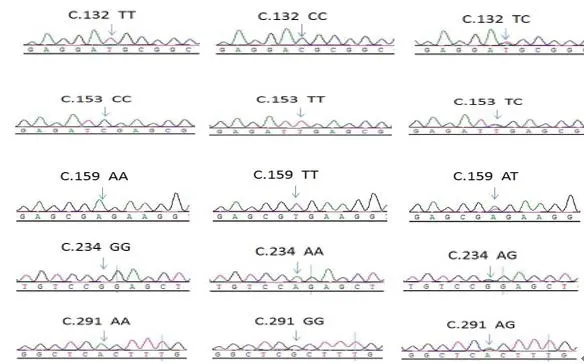
图1 AANAT基因的突变位
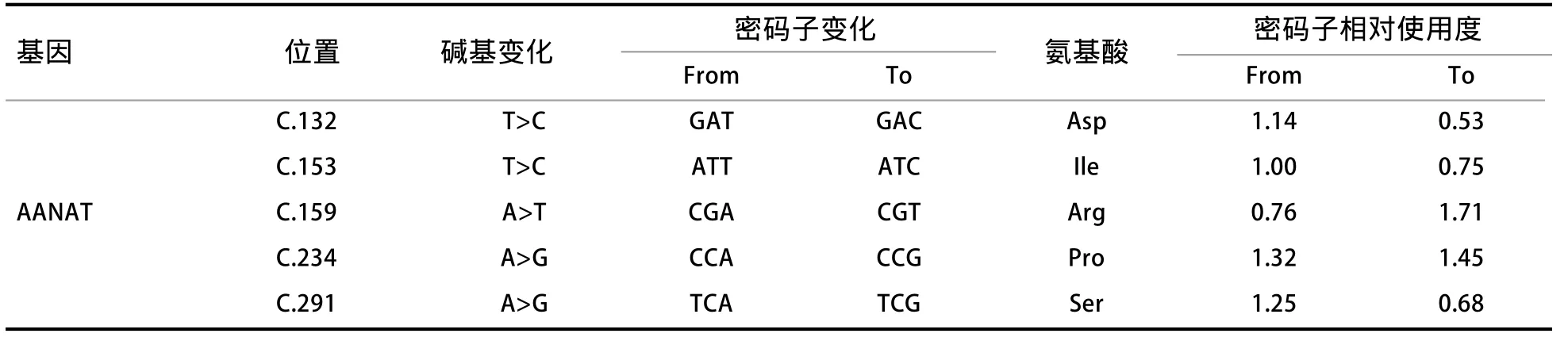
表2 AANAT基因CDS的突变位点
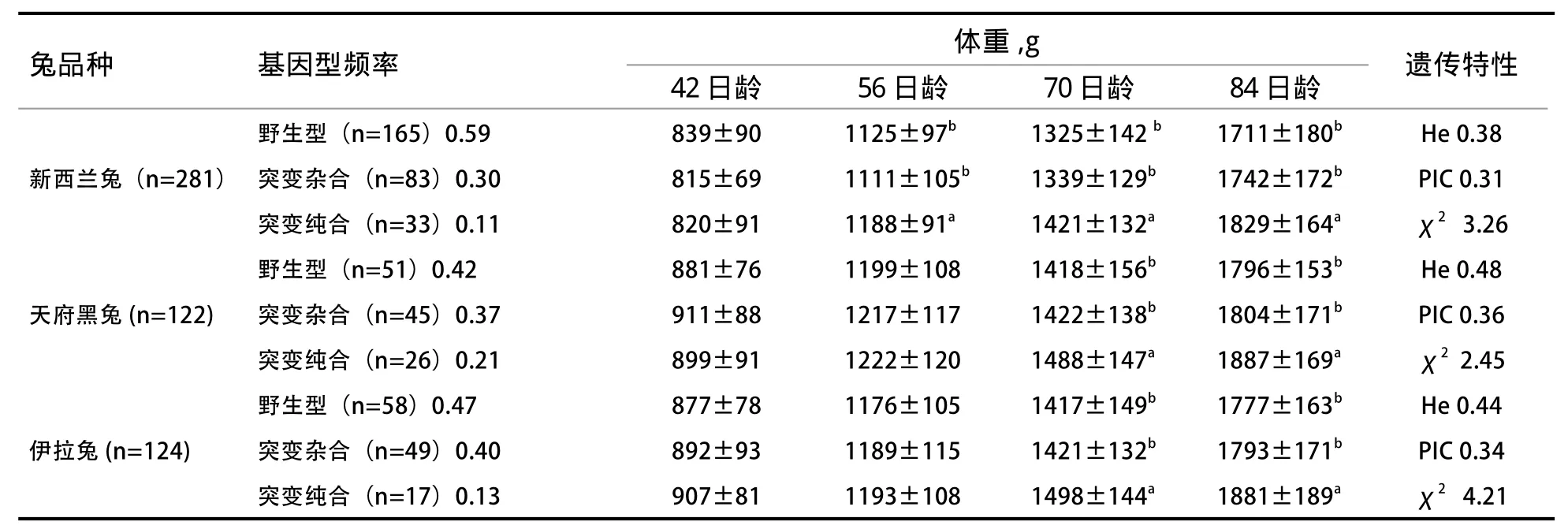
表3 AANAT基因类型和家兔生长性状的关联性分析
动物的生长受很多基因控制,这种现象称为微效多基因现象,如FTO[6]、IGFBP-5[7]、TBC1D1[8]等,这些基因对家兔的生长都起着调控作用。在动物的松果体中,AANAT基因编码1个乙酰转移酶超家族蛋白[9],对MT的合成起着重要的调控作用,是MT合成过程的限速酶[10]。MT合成是以色氨酸为原料,通过羟化、脱羧、乙酰化、氧甲基化最后合成MT,在脊椎动物中MT的主要作用有调节生物节律[11]、促进褐色脂肪生成[12]、抑制促性腺激素释放激素(GnRH)的释放[13]等,从而调控动物的生长以及性成熟[14-15]。在家畜中,Kashani等[16]研究发现,AANAT基因的表达跟脂肪代谢有直接关系,在山羊的日粮中添加螺旋藻提取物,使AANAT基因mRNA的表达量上升,提高了不饱和脂肪酸的含量,不利于山羊肌间脂肪的沉积。郝园丽[17]研究发现,AANAT基因的多态性对鸡的产蛋和开产日龄有显著影响,-708和-615位点连锁,单倍型TA使母鸡开产日龄延后10.3 d,-619和-595位点连锁,GC单倍型使母鸡的中期产蛋提高10.6枚。郭振刚等[18]研究发现,AANAT基因的第1个外显子129 bp处G>C、132 bp处C>T、141 bp处C>T,AA基因型对产绒量和绒长度有极显著的正效应,第2个外显子590 bp处C>T也对产绒量、绒长度和褪黑素含量有极显著正效应。
本实验在伊拉兔、天府黑兔和新西兰兔基因的CDS区中均发现5个完全连锁的同义突变多态位点,分别是C.132T>C、C.153 T>C、C. 159 A>T、C.234 A>G、C.291A>G,同义突变虽然没有改变蛋白质中氨基酸的组成,但是近年来的研究也表明,编码同一种氨基酸的不同密码子存在着使用偏好,这可以对基因的转录效率、mRNA的剪切以及空间结构、翻译效率等产生影响,从而影响蛋白质的结构、功能以及合成效率[19-21]。本实验仅研究了AANAT基因CDS区上面的突变位点和家兔生长性状的相关性,突变位点对AANAT基因的mRNA、蛋白质以及MT的表达量影响有待进一步验证。本实验发现,突变型新西兰兔在56、70、84日龄的体重显著高于野生型和杂合型,而突变型伊拉兔和天府黑兔在70日龄和84日龄的体重显著高于杂合型和野生型,伊拉兔、天府黑兔和新西兰兔群体AANAT基因的CDS突变属于中度多态(0.25<PIC<0.5),AANAT基因可以作为家兔生长性状的候选基因,AANAT基因CDS区上面的突变对家兔的生长有调节作用,本研究结果为进一步研究家兔育种提供科学依据。
4 结 论
本研究结果表明,家兔AANAT基因编CDS区的C.132 T>C、C.153 T>C、C. 159 A>T、C.234 A>G、C.291 A>G位点的多态性与家兔的生长性状显著相关。
[1] Huang Z, Liu T, Jimo B. N-terminal residues regulate proteasomal degradation of AANAT[J]. J Pineal Res, 2010,48(3):290-296.
[2] Reiter R J. Pineal melatonin: cell biology of its synthesis and of its physiological interactions.[J]. Endocr Rev, 1991,12(2):151-180.
[3] Kim T D, Woo K C, Cho S, et al. Rhythmic control of AANAT translation by hnRNPQ in circadian melatonin production.[J]. Genes Dev, 2007, 21(7):797-810.
[4] Zilberman-Peled B, Ron B, Gross A, et al. A possible new role for fish retinal serotonin- N -acetyltransferase-1(AANAT1): Dopamine metabolism[J]. Brain Res, 2006, s 1073-1074(1):220-228.
[5] Kim T D, Woo K C, Cho S, et al. Rhythmic control of AANAT translation by hnRNP Q in circadian melatonin production[J]. Genes Dev, 2007, 21(7):797.
[6] 高莲. 家兔FTO基因多态性及其与生产性状的相关性分析[D]. 雅安:四川农业大学, 2012.
[7] 李月英. 家兔IGFBP-5基因SNP多态性及其与生长性能的关联分析[D]. 雅安:四川农业大学, 2012.
[8] Yang Z J, Fu L, Zhang G W, et al. Identification and association of SNPs in TBC1D1 gene with growth traits in two rabbit breeds[J]. Asian-Austr J Anim Sci, 2013,26(11):1529-1535.
[9] Forrest R H, Hickford J G H, Hogan A, et al. Polymorphism at the B3-Adrenergic receptor locus: associations with birth weight, growth rate, carcass composition and cold survival[J]. Anim Genet, 2003, (34):19-25.
[10] Costa A S H D, Pires V M R, Fontes C M G A, et al.Expression of genes controlling fat deposition in two genetically diverse beef cattle breeds fed high or low silage diets[J]. BMC Vet Res, 2013, 9(1): 118.
[11] Guo B, Kongsuwan K, Greenwood P, et al. A gene expression estimator of intramuscular fat percentage for use in both cattle and sheep[J]. J Anim Sci Biotechnol,2014, 5(4):1-12.
[12] Holman B W B, Kashani A, Malau-Aduli A E O. Effects of Spirulina (Arthrospira platensis) supplementation level and basal diet on liveweight, body conformation and growth traits in genetically divergent Australian dual-purpose lambs during simulated drought and typical pasture grazing[J]. Small Rum Res, 2014,120(1):6-14.
[13] Arianna S, Patricia H P, María D C R, et al. Melatonin inhibits GnRH-1, GnRH-3 and GnRH receptor expression in the brain of the european sea bass, dicentrarchus labrax[J]. Int J Mol Sci, 2013, 14(4):7603-7616.
[14] Madgwick S, Evans A C O A P. Treating heifers with GnRH from 4 to 8 weeks of age advanced growth and the age at puberty[J]. Theriogenology, 2005, 63(8):2323-2333.
[15] 卢玉发, 廖清华. 褪黑素对肉鸡生长性能及免疫功能的影响[J]. 饲料工业, 2008, 29(13):37-39.
[16] Kashani A, Holman B W B, Nichols P D, et al. Effect of dietary supplementation with Spirulina on the expressions of AANAT, ADRB3, BTG2 and FASN genes in the subcutaneous adipose and Longissimus dorsi muscle tissues of purebred and crossbred Australian sheep[J]. J Anim Sci Biotechno , 2015, 57(1):1-8.
[17] 郝园丽. 寿光鸡AANAT基因5’调控区的多态性矿藏与产蛋性状的关联分析[D]. 泰安:山东农业大学, 2015.
[18] 郭振刚. 辽宁绒山羊褪黑激素合成酶(AA-NAT、TPH)基因克隆及遗传变异的研究[D]. 长春:吉林农业大学,2014.
[19] Chamary J V, Hurst L D. Evidence for selection on synonymous mutations affecting stability of mRNA secondary structure in mammals[J]. Genome Biol, 2005,(6): R75.
[20] Sauna Z E, Chava K S. Understanding the contribution of synonymous mutations to human disease[J]. Nat Rev Genet, 2011,12(10):683-691.
[21] Wu X, Tronholm A, Cáceres EF, et al. Evidence for deep phylogenetic conservation of exonic splice-related constraints: splice-related skews at exonic ends in the brown alga ectocarpus are common and resemble those seen in humans[J]. Genome Biol Evol, 2013, 5:1731-1745.
Association Analysis between Polymorphism of Coding Sequence of AANAT Gene and Growth Traits of Rabbits
LIU Bu-wei, WANG Jie, CHEN Shi-yi, JIA Xian-bo, LAI Song-jia*
(Institute of Animal Genetics and Breeding, Sichuan Agricultural University, Sichuan Chengdu 611130, China)
To study the association between Coding sequence(CDS)of five HT -N- acetyltransferase gene ( AANAT )polymorphism and growth traits in rabbits. In the study we used direct sequencing method to detect single nucleotide polymorphism of exons of AANAT gene region with Tianfu black rabbit, IRA rabbit and New Zealand rabbit as experimental materials. There were 5 complete linkage mutation sites(C.132 T>C, C.153T>C, C. 159 A>T, C.234 A>G, C.291A>G) in the CDS of AANAT gene in the three rabbit breeds. 3 genetypes(wild type: TTTTAAAAAA, heterozygous: TCTCATAGAG mutant type: CCCCTTGGGG ) were formed. In New Zealand, the 56, 70 and 84 days body weight of mutant type was significantly higher than those of other types rabbits and in the IRA rabbit and Tianfu black rabbit (P<0.05),70 and 84 days body weight were higher (P<0.05). The AANAT gene can be used as a candidate gene for growth traits in rabbit.
Rabbit ; AANAT gene; SNP; Growth performance
S829.2
A
10.19556/j.0258-7033.2017-10-025
2017-03-23;
2017-04-26
四川省科技支撑计划项目(2016NYZ0046、2016NYZ0050)
刘卜玮(1993-),男,四川南充人,硕士研究生,研究方向为动物遗传育种与繁殖,E-mail:liubuwei@yeah.net
*通讯作者:赖松家,教授,研究方向为生物技术与兔遗传育种,E-mail: laisj5794@gmail.com
