干旱胁迫对高羊茅航天诱变新品系生理特性的影响及综合评价
2017-10-21李娟雷霞王小利牟琼杨春燕吴佳海
李娟,雷霞,王小利,牟琼,杨春燕,吴佳海
(贵州省农业科学院草业研究所,贵州 贵阳 550006)
干旱胁迫对高羊茅航天诱变新品系生理特性的影响及综合评价
李娟,雷霞,王小利,牟琼,杨春燕,吴佳海*
(贵州省农业科学院草业研究所,贵州 贵阳 550006)
为揭示高羊茅航天诱变新品系对干旱胁迫的响应和适应,培育出耐旱性强的新品种。以高羊茅航天诱变新品系12份及其亲本黔草1号高羊茅和贵州主推品种水城高羊茅为材料,采用盆栽试验,在人工气候室进行干旱胁迫处理,测定可溶性蛋白、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性、叶绿素(Chl)生理指标,并应用隶属函数法和灰色关联法对其生理指标及14份高羊茅材料耐旱性进行综合评价。结果表明,可溶性蛋白质随着干旱胁迫程度的增加总体呈上升趋势,SOD、POD、CAT含量随着干旱胁迫程度增加总体呈下降趋势,叶绿素含量随干旱胁迫程度加剧呈下降趋势。5个生理指标在14份材料间均存在显著性差异。利用隶属函数法对14份高羊茅材料进行综合耐旱性评价,SP5-32的平均隶属函数值最大,为0.533,耐旱性顺序为:SP5-32>SP5-75>水城高羊茅>SP5-60>SP5-94>SP5-97>SP5-89>SP5-71>SP5-7>SP5-85>SP5-42>SP5-5>黔草1号高羊茅>SP5-88。利用灰色关联性分析法得出各耐旱指标与耐旱性的关联序为:POD>可溶性蛋白质>SOD> CAT>Chl,关联度在0.55~0.65之间。因此,SP5-32更加适应干旱地区,干旱胁迫下保护酶活性大小、可溶性蛋白质含量与叶绿素含量对高羊茅材料的耐旱性是大致相同的。
高羊茅;干旱胁迫;航天诱变;生理特性;综合评价
我国干旱半干旱地区面积占国土面积的47%,干旱已成为全球自然灾害之一[1-2],可使其植物生长受到抑制、产量下降[3]。高羊茅(Festucaarundinacea)又称苇状羊茅,是禾本科羊茅属一种多年生冷季型草种[4-5]。是重要的饲用、城市、运动场绿化植物,生长迅速、饲用品质优良、利用年限长,在市场经济中显示出明显的社会、生态、环境效益,因而越来越受到人们的重视。然而,在当今水资源日益缺乏的时候,人们首先会满足生活用水和粮食灌溉的需求,对于草建植等环境景观等方面的需求就会受到限制[6],因此,通过各种不同的选育方法,培育出耐旱新品种,成为高羊茅育种中的当务之急。
在自然条件下,由于外界环境的变化较小和遗传结构的相对稳定性,植物本身发生自发突变的频率极低,加之传统的育种方式周期长,分子育种经费高、难度大等问题[7]。将高科技的空间技术与常规农业育种相结合,形成航天诱变育种,是利用太空特殊环境,如空间宇宙射线、微重力、高真空、弱磁场等因素,对农作物种子的诱变作用产生变异,再返回地面选育新种质、新材料,培育新品种的作物育种新技术。近年来牧草和草坪草在航天诱变育种方面发展迅速,先后搭载了红豆草(Onobrychisviciaefolia)、苜蓿(Medicagosativa)、草地早熟禾(Poaspp.)、结缕草(Zoysiajaponica)、狗牙根(Cynodondactylon)等[8-11],并对搭载材料的形态学、细胞学和分子水平的变异进行了研究,通过航天诱变已筛选、培育出很多优良品种,航天诱变作为一种新的育种方法已受到国内外遗传育种界的重视[12-13]。航天诱变育种既能明显改良作物某些农艺性状,又能获得地面育种难以得到的重要经济性状产生的突破性突变。本研究基础正是基于对黔草1号高羊茅种子经“实践八号”育种卫星搭载处理,在开展多年的“地-空”对比试验研究,发现有很多优良突变植株,经多年多代选择,初步筛选出不同的优良材料,深入系统开展高羊茅航天诱变育种。选取不同高羊茅航天诱变新品系进行干旱胁迫试验,研究其对干旱胁迫的生理响应和调节机制,为西南地区高羊茅的逆境生理和育种提供理论依据。
1 材料与方法
1.1试验材料
选取多年生黔草1号高羊茅种子,于2006年9月9日经“实践八号”育种卫星搭载处理,卫星共运行355 h,航程900多万km,卫星运行轨道为倾角63°,近地点180 km、远地点460 km,在轨运行15 d后,在四川遂宁回收,留轨舱进行3 d留轨试验。选取经搭载诱变后育成材料SP5-88、SP5-42、SP5-94、SP5-75、SP5-5、SP5-85、SP5-60、SP5-89、SP5-71、SP5-32、SP5-7、SP5-97及其亲本黔草1号高羊茅和贵州主推品种水城高羊茅共14份材料。
1.2试验设计
各试验材料在贵州省草业研究所人工气候室进行分株繁殖,2013年4月23日采用盆栽方式。选用塑料花盆(花盆规格:15 cm×15 cm×10 cm),取田间表土(黄壤),粉碎混匀,不做灭菌处理,每盆装干土0.7 kg。每盆移栽6~7个分蘖枝,剪去地上部分,定期浇水,以保证幼苗的正常生长,置于温度为25 ℃、光照为4000 lx的人工气候室中。待植株生长正常,叶片长到20 cm长时,每盆定植5个分蘖枝,进行干旱胁迫。1盆为一个重复,每个材料12盆,设3个水分处理,4次重复,完全随机区组排列。处理A(无水分胁迫)为田间持水量的85%~80%;处理B(中度水分胁迫)为田间持水量的60%~55%;处理C(重度水分胁迫)为田间持水量的35%~30%。处理前,每盆加水至土壤达到饱和状态,然后使其自然干旱至设定标准,并维持。处理中,为了降低土壤表面水分蒸发,在盆土表面覆保鲜膜,并且定期移动植物的位置,使每个植物呈随机排列。待各水分梯度形成后开始干旱胁迫,水量用称重法控制,于每天17: 00向盆中插孔补充水分,在保持设定的含水量15 d后进行生理指标的测定。在早晨8: 00-9: 00选取相同部位的成熟健壮叶片进行生理指标的测定,每个指标重复测定3次。
1.3测定指标与方法
各生理生化指标参照李合生[14]的方法测定。可溶性蛋白质采用考马斯亮蓝G-250比色法;超氧化物歧化酶(SOD)采用氮蓝四唑光还原法;过氧化物酶活性(POD)采用愈创木酚法;过氧化氢酶(CAT)活性采用钼酸铵显色法;叶绿素含量的测定采用浸提法。
1.4耐旱性综合评价——隶属函数值法
本研究根据测定的各项生理生化指标,采用模糊数学中的隶属函数值法进行耐旱性综合评价[15-18]。如果某一指标与耐旱性呈正相关,则采用反隶属函数计算,公式为:
X(u1)=(X-Xmin)/(Xmax-Xmin)
(1)
如果某一指标与耐旱性呈负相关,则采用反隶属函数计算,公式为:
X(u2)=1-(X-Xmin)/(Xmax-Xmin)
(2)
式中,X(u1)、X(u2)为隶属函数值,X为干旱胁迫下各材料某一指标的测定值,Xmin、Xmax分别为所有参试材料某一指标的最小值和最大值。将所有参试材料的抗旱隶属函数值加起来,求其平均值,用平均值大小综合评价各材料耐旱性的强弱。
1.5灰色关联性分析
灰色关联度分析被广泛应用于农业和林业研究中[15]。将各种耐热指标的平均隶属函数值作为参考数列,记为X0,以各耐热指标的平均值为比较数列,记为Xi(i=0,1,2,…,n), 则参考数列X0={X0(1),X0(2),…,X0(n)}, 比较数列Xi={X1(1),X1(2),…,X1(n)}。利用公式(3)对原始数据进行无量纲化处理,
(3)
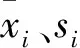
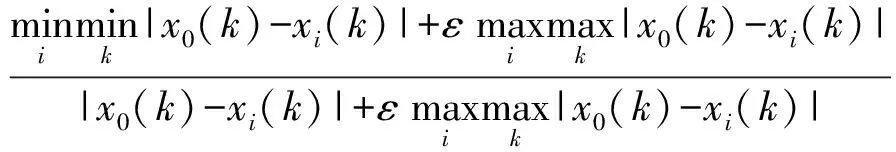
(4)
(5)
式中:εi(k)为比较数列xi对参考数列x0在第k点的关联系数,ri为比较数列xi与参考数列x0的关联度,ε为分辨系数,一般取值范围为0∽1,本试验取值为0.5[13-15]。i,k=1,2,…,n。
1.6数据统计
试验每个处理、各指标测定均重复3次,采用Excel 2003和SPSS 19.0统计软件对数据进行统计分析,选择单因素方差分析(one way ANOVA)进行差异显著性检验。
2 结果与分析
2.1可溶性蛋白质含量
图1表明,14份高羊茅航天诱变材料可溶性蛋白质含量随田间持水量减少呈上升趋势,其中SP5-88、SP5-94、SP5-60三份材料呈先升高后下降的趋势,在中度胁迫时达到最大值,分别是1.285、1.421、0.954 g/L,比无水分胁迫升高了0.279、0.569、0.459 g/L;SP5-75、SP5-5、SP5-85、SP5-89、SP5-71呈先下降后升高的趋势,中度胁迫时达到最低,分别是0.601、1.191、0.697、1.018、0.665 g/L,比无水分胁迫时降低了0.224、0.128、0.089、0.462、0.196 g/L;SP5-42、SP5-32、SP5-7、SP5-97、黔草1号高羊茅、水城高羊茅呈上升趋势。由图1还发现,由于材料抗旱性的差异,随着干旱程度的增加,可溶性蛋白质变化趋势呈现3种情形,但总体趋势都是趋于上升。SP5-88、SP5-42、SP5-94、SP5-75、SP5-5、SP5-85、SP5-60、SP5-32、SP5-7、SP5-97、黔草1号高羊茅和水城高羊茅12份材料重度胁迫比无水分胁迫分别上升了0.133、0.454、0.229、0.144、0.066、0.153、0.171、0.346、0.402、0.205、0.220、0.404 g/L,SP5-89和SP5-71材料重度胁迫比无水分胁迫分别下降了0.177和0.006 g/L。

图1 干旱胁迫对可溶性蛋白质含量的影响Fig.1 Effect of drought stress on the soluble protein content 1:黔草1号高羊茅F. arundinacea cv. Qiancao No.1; 2:水城高羊茅F. arundinacea cv. Shuicheng.大写字母不同,表示同一试材在不同胁迫间差异显著(P<0.05);小写字母不同,表示同一胁迫下不同试材间差异显著(P<0.05)。下同。Different capital letters indicate significant differences in the same experimental material under different treatments (P<0.05); Different small letters indicate significant differences between different experimental materials in the same treatment (P<0.05).The same below.
2.2SOD活性
图2显示,14份高羊茅航天诱变材料SOD活性随田间持水量减少呈下降的趋势,其中SP5-88、SP5-94、SP5-60、SP5-7四份材料呈先下降后上升的趋势,在中度胁迫时达到最小值,分别是19.461、18.073、27.906、27.042 U/mg,比无水分胁迫下降了9.211、11.346、24.913、22.534 U/mg;SP5-75、SP5-5、SP5-85、SP5-89、SP5-71呈先升高后下降的趋势,中度胁迫时达到最大,分别是34.721、19.953、36.858、26.864、38、197 U/mg,比无水分胁迫时升高了13.024、0.983、5.203、12.073、9.954 U/mg;SP5-42、SP5-32、SP5-97、黔草1号高羊茅、水城高羊茅呈下降趋势。表明由于材料抗旱性的差异,随着干旱程度的增加,SOD活性变化趋势呈现3种情形,但总体趋势都是趋于下降。SP5-88、SP5-42、SP5-94、SP5-5、SP5-85、SP5-60、SP5-32、SP5-7、SP5-97、黔草1号高羊茅、水城高羊茅11份材料重度胁迫比无水分胁迫分别下降了5.004、11.668、9.636、1.838、6.974、10.755、13.356、20.837、3.309、3.845、8.370 U/mg。SP5-75、SP5-89、SP5-71材料重度胁迫比无水分胁迫分别上升了1.576、3.948、0.444 U/mg。

图2 干旱胁迫对SOD活性的影响Fig.2 Effect of drought stress on the SOD activity
2.3POD活性
干旱胁迫下14份高羊茅航天诱变材料POD活性变化如图3所示。结果表明,14份高羊茅航天诱变材料POD活性随田间持水量减少呈下降的趋势,其中SP5-88、SP5-94、SP5-60、水城高羊茅4份材料呈先下降后上升的趋势,在中度胁迫时达到最小值,分别是31.532、28.509、49.326、37.106 U/mg,比无水分胁迫下降了10.334、23.668、16.752、21.961 U/mg;SP5-75、SP5-5、SP5-85、SP5-89、SP5-71呈先升高后下降的趋势,中度胁迫时达到最大,分别是69.048、37.439、60.003、42.476、70.248 U/mg,比无水分胁迫时升高了14.368、1.898、10.440、14.687、47.135 U/mg;SP5-42、SP5-32、SP5-7、SP5-97、黔草1号高羊茅呈下降趋势。表明:随着干旱程度的增加,POD活性呈现3种变化趋势,但总体趋势都是趋于下降。SP5-88、SP5-42、SP5-94、SP5-75、SP5-5、SP5-85、SP5-60、SP5-32、SP5-7、SP5-97、黔草1号高羊茅、水城高羊茅12份材料重度胁迫比无水分胁迫分别下降了4.230、23.011、7.515、11.920、2.640、4.226、3.181、48.072、39.392、5.762、7.982、21.307 U/mg。SP5-89、SP5-71材料重度胁迫比无水分胁迫分别下降了-7.561、-31.568 U/mg。

图3 干旱胁迫对POD活性的影响Fig.3 Effect of drought stress on the POD activity
2.4CAT活性
图4所示,14份高羊茅航天诱变材料CAT活性随田间持水量减少呈下降的趋势,其中SP5-94、SP5-60、SP5-7、水城高羊茅四份材料呈先下降后上升的趋势,在中度胁迫时达到最小值,分别是6.849、7.170、7.709、7.808 U/mg,比无水分胁迫下降了4.322、2.979、2.503、5.906 U/mg;SP5-42、SP5-75、SP5-5、SP5-85、SP5-89、SP5-71、SP5-32、SP5-97、黔草1号高羊茅呈先升高后下降的趋势,中度胁迫时达到最大,分别是9.599、18.645、8.135、10.737、11.353、13.814、15.472、6.374、8.435 U/mg,比无水分胁迫时升高了1.556、7.440、0.066、4.743、4.729、0.884、2.856、0.577、1.658 U/mg;SP5-88呈下降趋势。表明:随着干旱程度的增加,CAT活性呈现3种变化趋势,但总体趋势都是趋于下降。SP5-88、SP5-94、SP5-75、SP5-5、SP5-71、SP5-32、SP5-7、SP5-97、水城高羊茅9份材料重度胁迫比无水分胁迫分别下降了2.128、2.086、2.786、0.539、3.031、2.921、1.808、1.626、5.337 U/mg。SP5-42、SP5-85、SP5-60、SP5-89、黔草1号高羊茅材料重度胁迫比无水分胁迫分别上升了-1.391、-0.728、-5.167、-2.155、-0.533 U/mg。
2.5叶绿素含量
由图5可知,14份高羊茅航天诱变材料叶绿素含量随田间持水量减少呈下降的趋势,其中SP5-75、SP5-5两份材料呈先下降后上升的趋势,在中度胁迫时达到最小值,分别是1224.612、765.501 mg/g,比无水分胁迫下降了515.825、377.949 mg/g;SP5-88、SP5-60、SP5-71、黔草1号高羊茅、水城高羊茅呈先升高后下降的趋势,中度胁迫时达到最大,分别是793.687、998.301、803.752、1259.609、1985.653 mg/g,比无水分胁迫时升高了65.352、145.270、13.320、44.439、179.984 mg/g;SP5-42、SP5-94、SP5-89、SP5-32、SP5-97呈下降趋势;SP5-85、SP5-7呈上升趋势。表明:随着干旱程度的增加,叶绿素含量呈现4种变化趋势,但总体都是趋于下降。SP5-88、SP5-42、SP5-94、SP5-75、SP5-5、SP5-89、SP5-71、SP5-32、SP5-97、黔草1号高羊茅、水城高羊茅11份材料重度胁迫比无水分胁迫分别下降了237.742、231.353、428.756、113.186、53.599、360.832、166.283、582.001、217.291、656.996、554.477 mg/g;SP5-85、SP5-60、SP5-73材料重度胁迫比无水分胁迫分别上升了221.390、114.010、197.483 mg/g。
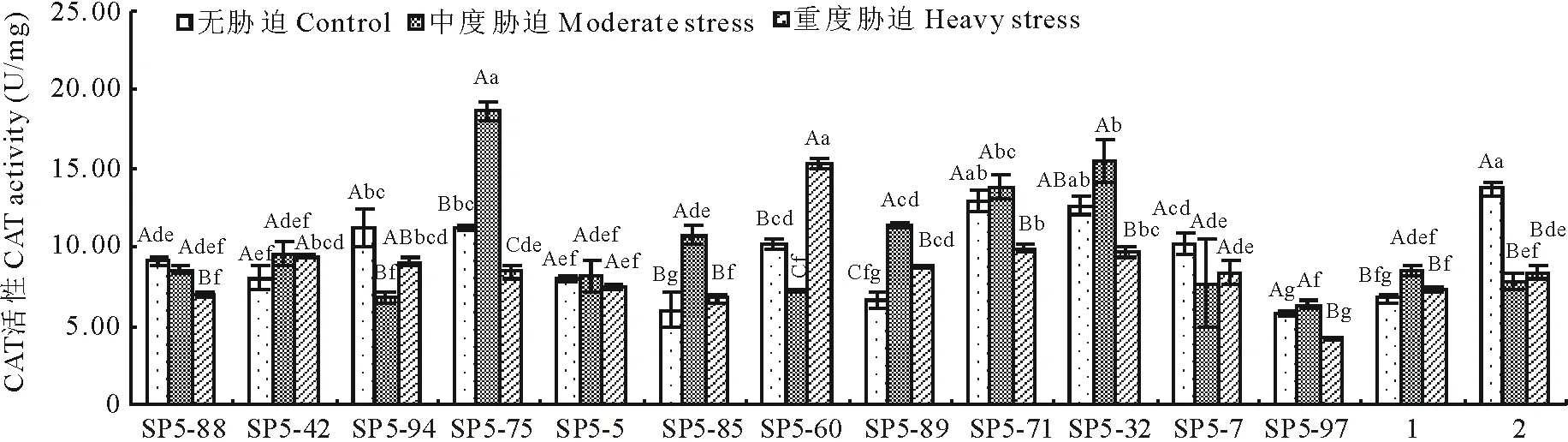
图4 干旱胁迫对CAT活性的影响Fig.4 Effect of drought stress on the CAT activity
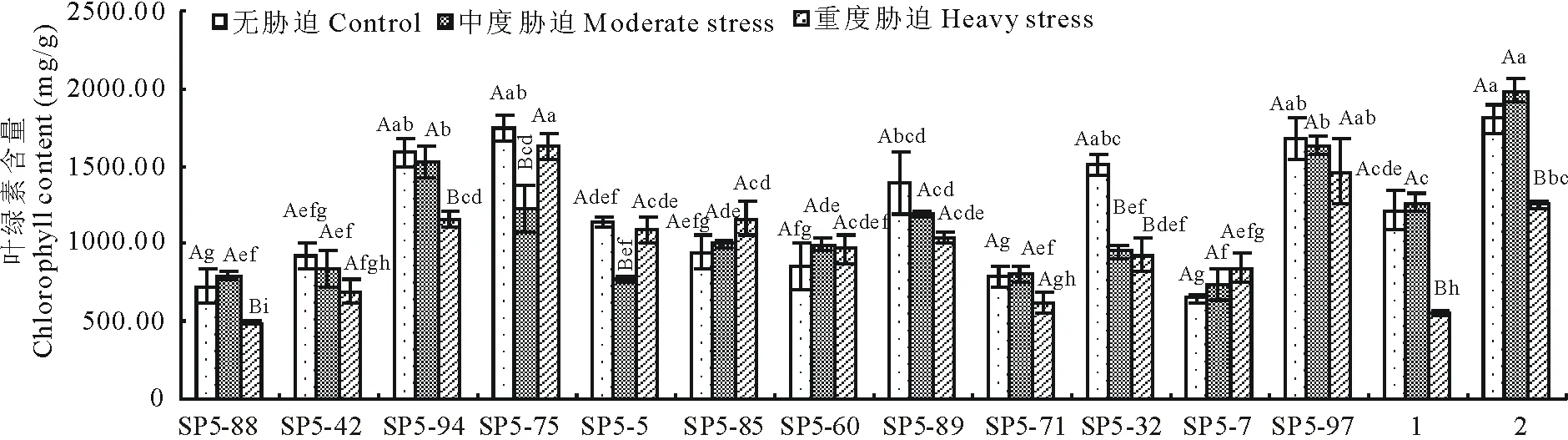
图5 干旱胁迫对叶绿素含量的影响Fig.5 Effect of drought stress on the Chl content
2.6抗旱性评价——隶属函数值法
植物的耐旱性是由多种指标综合作用的遗传表现,单一的指标对植物耐旱性的判断往往有一定的局限性,难于判断植物对干旱的综合适应能力,必须用多指标进行综合评价[19-20]。以可溶性蛋白质、SOD、POD、CAT、叶绿素5个指标为依据,计算各指标的隶属函数值,并进行耐旱性综合评价见表1。综合抗旱性强弱顺序为:SP5-32>SP5-75>水城高羊茅>SP5-60>SP5-94>SP5-97>SP5-89>SP5-71>SP5-7>SP5-85>SP5-42>SP5-5>黔草1号高羊茅>SP5-88。除SP5-88,其他诱变材料的抗旱性都大于黔草1号高羊茅,其中SP5-32的平均隶属函数值最大,为0.533,SP5-88的平均隶属函数值最小,为0.297,黔草1号高羊茅平均隶属函数值也小,为0.328,说明航天诱变可以产生好的抗旱材料,且SP5-32材料产生的变异,更加适应干旱地区。
2.7耐旱指标的相关关系评价——灰色关联性分析
灰色关联分析是对于系统之间的因素,随发展趋势的相似或相异程度,来衡量关联程度的一种分析方法,意图透过一定的方法,去寻求系统中各因素之间的数值关系。因此,灰色关联度分析对于一个系统发展变化态势提供了量化的度量,非常适合动态历程分析。
由于各指标对耐旱性的贡献大小不同,按各指标的相对重要程度求出关联度,才能准确、全面地评价其耐旱性[19]。将14份材料5个指标的平均隶属函数值与14份材料的抗旱指标看作是一个灰色系统,平均隶属函数值作为数列X0,可溶性蛋白质、SOD、POD、CAT、叶绿素5个指标作为比较数列Xi,14份材料的抗旱指标的平均值及标准化处理后的结果见表2,利用公式(4)、(5)计算各抗旱指标和抗旱性的关联系数、关联度,并根据大小排列出关联序。关联度大小可表示某一抗旱性指标对干旱的敏感程度,关联度越大,表明对干旱的敏感程度越高,反之则越低。从表3可以看出,各抗旱指标与耐旱性的关联序为:POD>可溶性蛋白质>SOD>CAT>Chl。5个指标的关联度都在0.55~0.65之间,分别是0.610、0.599、0.642、0.596、0.584,通过关联分析,说明干旱胁迫下保护酶活性大小、可溶性蛋白质含量与叶绿素含量对高羊茅材料的耐旱性是大致相同的。
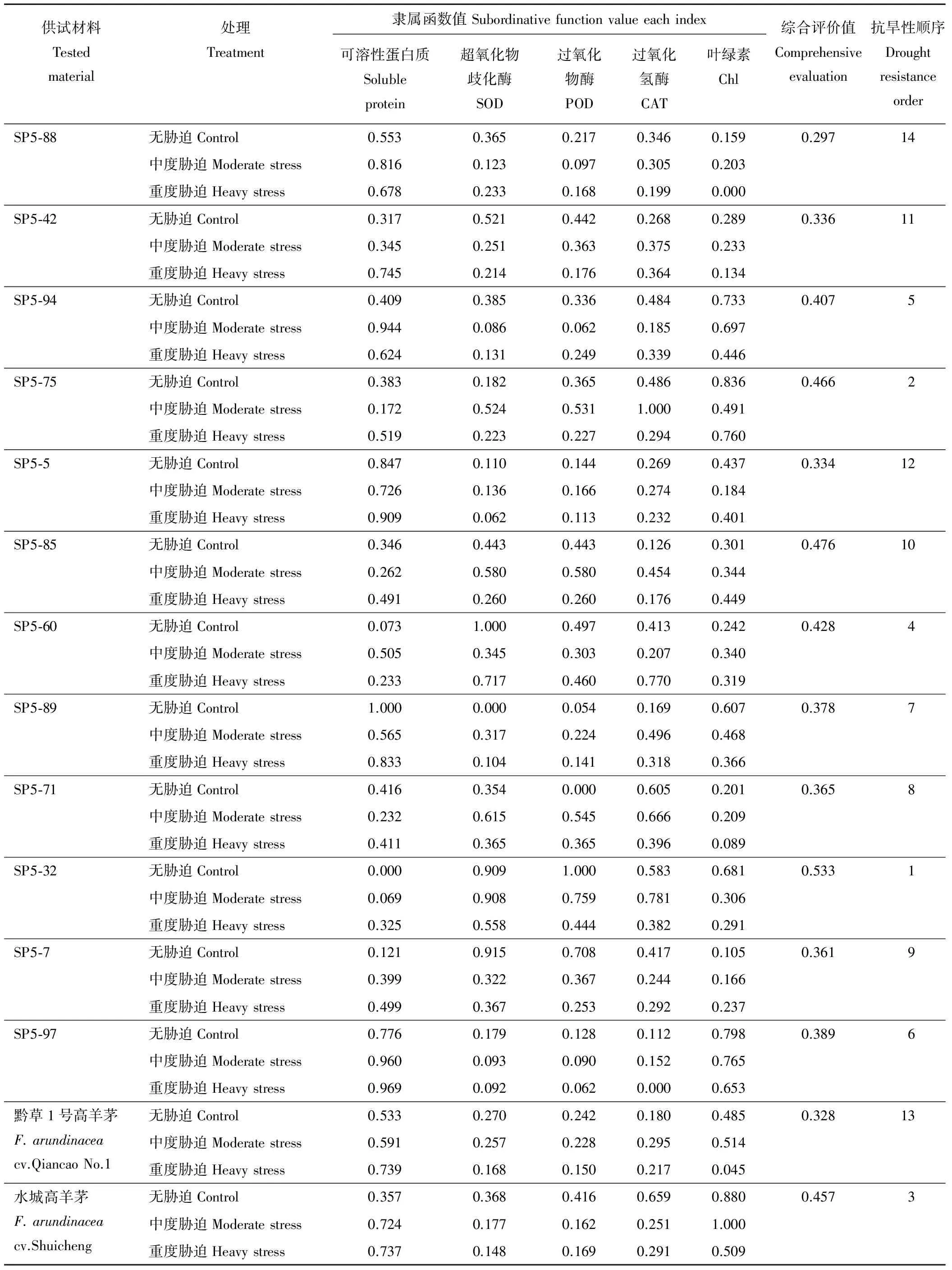
表1 抗旱性综合评价Table 1 Drought resistance comprehensive evaluation
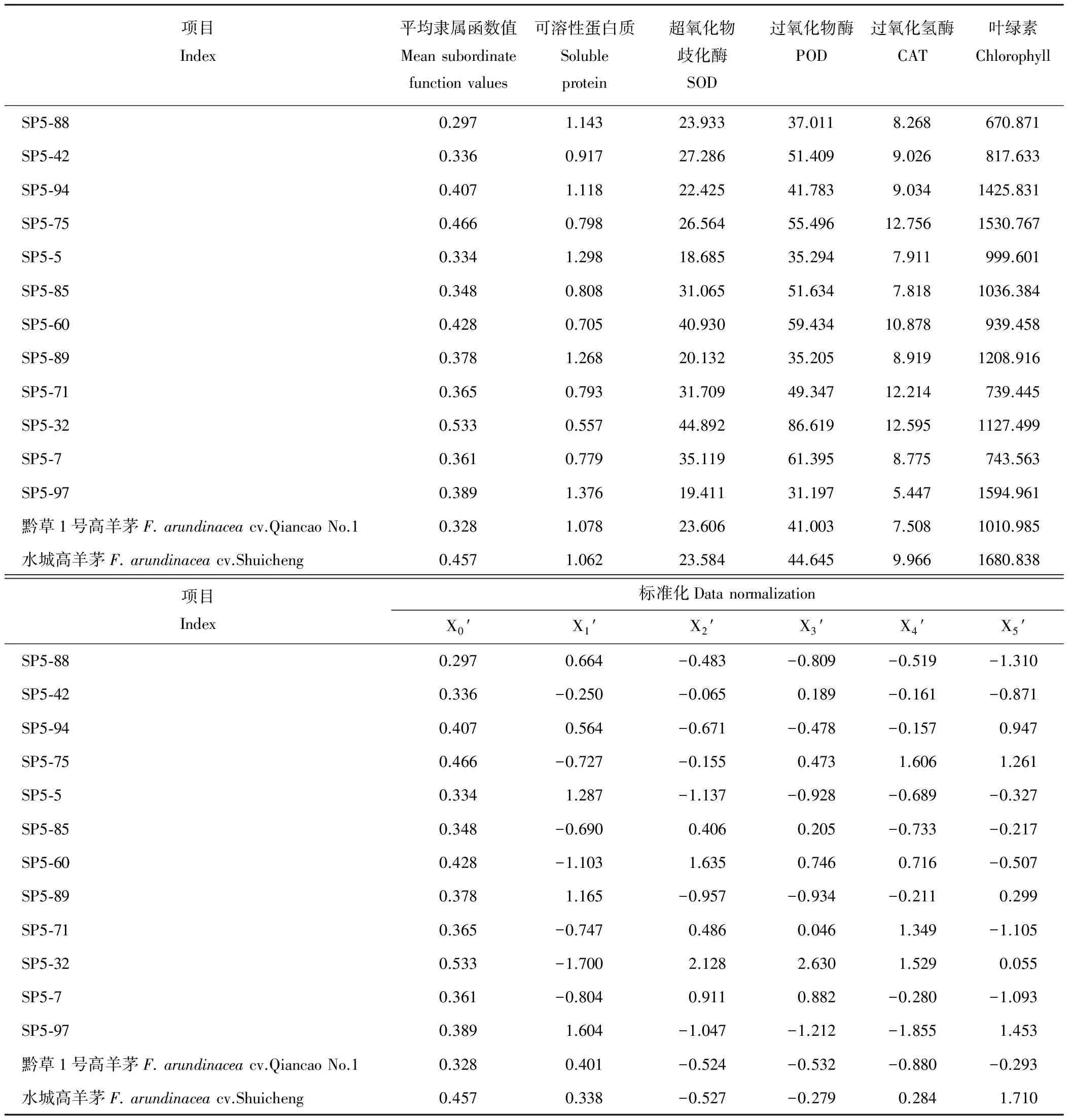
表2 抗旱指标及标准化处理Table 2 Drought resistance indexes and data normalization
3 讨论与结论
蛋白质是生物体内重要的大分子之一,受多种因素的影响。变化的环境因子或逆境胁迫包括干旱、水涝、盐渍等环境条件都会影响植物体内蛋白质(酶)等分子遭受破坏,植物为了避免造成伤害,会诱导产生一些抗逆蛋白,这些新增蛋白的种类和含量与植物的耐旱性密切相关[21-22]。可溶性蛋白增多可帮助维持植物细胞较低的渗透势,提高细胞内溶质浓度,降低水势,使细胞能从外界吸收水分,从而抵抗干旱胁迫导致的伤害[23]。供试14份材料,在受到干旱胁迫时可溶性蛋白质总体呈上升趋势,可能是植物体内产生更多的可溶性蛋白质或者细胞内一些不溶性蛋白质转变为可溶性蛋白质,以抵抗缺水的威胁,使细胞内正常的新陈代谢得以维持[24-25];每个时期的变化趋势又因材料不同而有差异,这说明,可溶性蛋白质的变化趋势与植物本身的特性或胁迫所处的不同阶段有关,从而使其变化趋势有所差异;在重度胁迫下,抗旱性强的品种可能启动与适应干旱胁迫有关的基因,诱导产生新的胁迫诱导蛋白,从而在干旱条件下能够维持正常的产量或尽量减少产量损失;抗旱性弱的品种,新的蛋白质不能产生,只能随着胁迫程度的加剧而阶段性的促进原有蛋白质的合成和分解,不能继续合成和分解时则不能维持正常的可溶性蛋白质浓度[21,26],从而使可溶性蛋白质含量下降。
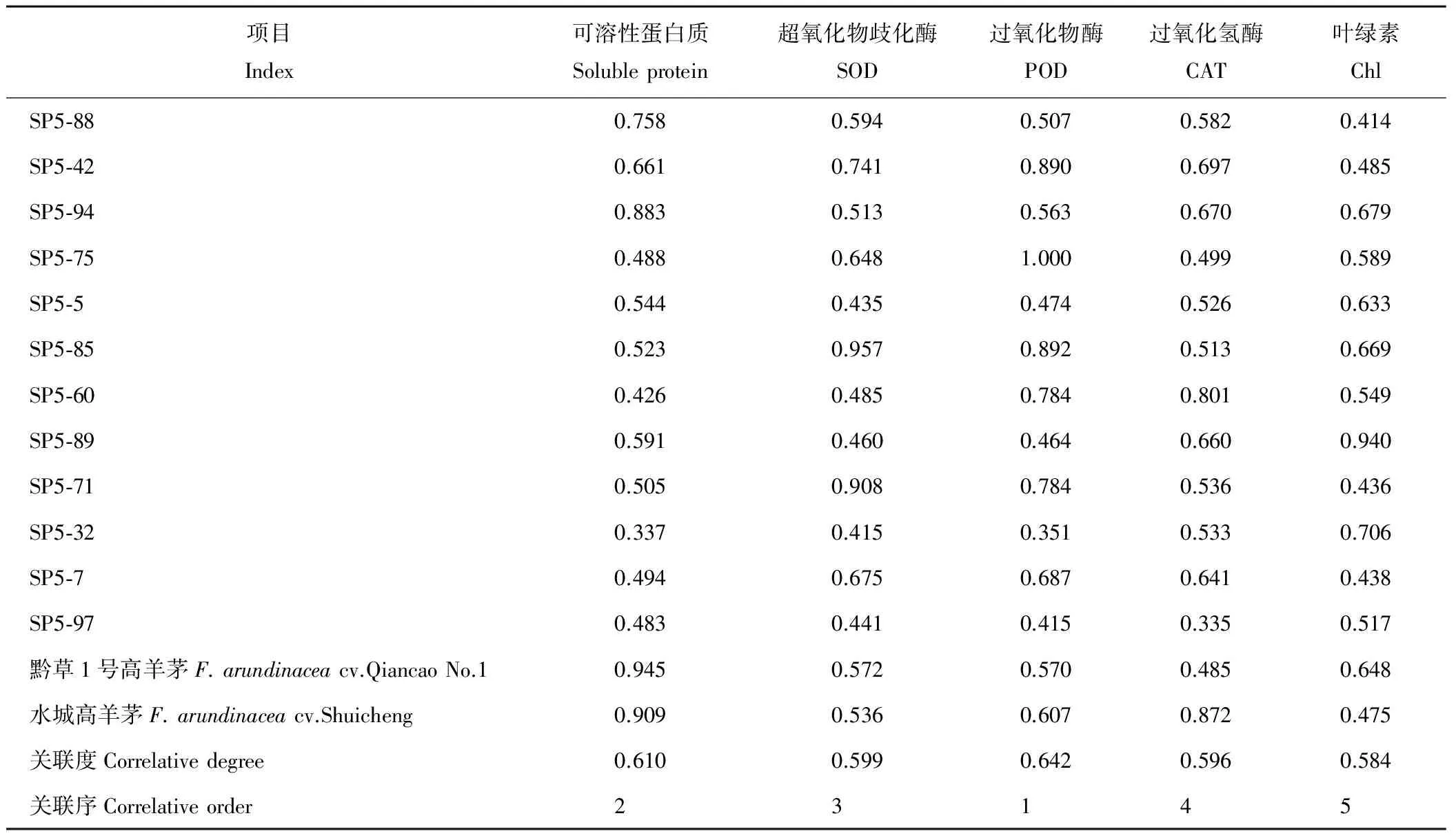
表3 抗旱性与各指标的关联系数、关联度、关联序Table 3 Drought resistance and correlative modulus, correlative degree and correlative order
在逆境条件下,SOD、POD、CAT等活性的变化有利于植物度过困境,是植物体内重要的活性氧清除酶[27]。在正常的生理条件下,细胞内自由基的产生和消除处于动态平衡状态,自由基水平很低,不会造成对植物的伤害;植物在逆境条件下,平衡被打破,细胞内产生过量的自由基,会引发或加剧膜脂过氧化而造成生物膜系统的损伤,对植物造成伤害,严重时会导致死亡。供试14份材料SOD、POD、CAT随干旱胁迫程度的加剧总体呈下降趋势,但其变化趋势又因材料间的差异而不同,造成SOD、POD、CAT对干旱胁迫程度的响应不一致,可能是过多的自由基超出机体的清除能力,影响物质的正常代谢,对生物体造成伤害,也可能是活性氧清除系统遭到破坏导致SOD、POD、CAT活性的降低[28]。轻度胁迫下降,重度胁迫又上升,可能是植物对轻度胁迫的一种适应性反应,使能量消耗减少,通过正常的生理调节起作用[29],胁迫程度加剧,活性氧清除系统诱导SOD、POD、CAT活性的大幅增加,以此来抵御外界胁迫的适应性反应。随着胁迫程度的加剧,有的又呈下降趋势可能是由于活性氧生成迅速且含量急剧增多,也可能在重度干旱条件下,植物无法发挥清除活性氧的能力,最终抑制了植物的生长[30]。
叶绿体是植物进行光合作用的主要场所,在干旱胁迫下叶绿素含量直接受叶片含水量的影响。在干旱胁迫下,叶片含水量下降,叶绿体结构遭到破坏,从而导致植物体内叶绿素含量下降。供试14份材料叶绿素含量随干旱胁迫程度加剧呈下降趋势。但变化趋势又不同。轻度胁迫出现上升趋势,可能是由于轻度干旱胁迫促进了叶片对水分的吸收和贮藏,也可能是植物对环境因子的补偿和超补偿效应[31],当干旱胁迫程度加剧,叶绿素含量下降,可能是重度胁迫引起植物体内生理生化改变,使叶绿素合成受阻,降解加快。
植物的抗旱性是对土壤干旱和大气干旱的适应能力[32],不同植物对干旱的抗性只是程度上的差异,在不同的干旱胁迫下可以区分不同植物的抗旱能力,若在植物对干旱的耐受范围,植物会对干旱胁迫迅速产生应激机制,对植物的伤害就会产生一定的缓冲作用,使其在干旱条件下的伤害降到最低[33-34],若超越植物对干旱的耐受范围,几乎所有的植物的抗旱力就会消失。干旱条件下,植物能够调控与抗旱有关的基因表达,随之产生一系列形态、生理生化等方面的变化来显示出其抗旱能力[19,35]。本试验测定干旱胁迫下14份高羊茅材料随着干旱胁迫程度不同,可溶性蛋白质含量、SOD含量、POD含量、CAT含量、叶绿素含量5个指标的变化,研究表明,可溶性蛋白质随着干旱胁迫程度的加剧总体呈上升趋势,SOD、POD、CAT含量、叶绿素含量呈下降趋势。5个生理指标在14份材料间均存在显著性差异。近年来常采用主成分分析法、聚类分析法、模糊数学中的隶属函数法等,来评价多个指标共同作用的结果,减少单个指标对材料评价的片面性[28]。本研究采用隶属函数法,对14份高羊茅航天诱变新品系进行耐旱性综合评价,得出耐旱性顺序为:SP5-32>SP5-75>水城高羊茅>SP5-60>SP5-94>SP5-97>SP5-89>SP5-71>SP5-7>SP5-85>SP5-42>SP5-5>黔草1号高羊茅>SP5-88。同时采用灰色关联性分析法得出各耐旱指标与耐旱性的关联序为:POD>可溶性蛋白质>SOD>CAT>Chl。本研究只对高羊茅航天诱变新品系5个生理生化指标进行了测定,还需从水分、光合、激素等方面进行进一步的研究。
References:
[1] Peng K S, Xu X B, Hu J H,etal. The harm of drought to the west region and its prevention strategy. Journal of Shijiazhuang University of Economics, 2002, 25(3): 257-262.
彭珂珊, 徐宣斌, 胡晋辉, 等. 干旱是西部地区生态系统受损的关键因素. 石家庄经济学院学报, 2002, 25(3): 257-262.
[2] Jiang H, Bi Y F, Chen L X,etal. Physiological characteristics of alfalfa under dry-farming conditions. Acta Agrestia Sinica, 2012, 20(6): 1077-1080.
姜华, 毕玉芬, 陈连仙, 等. 干旱条件下紫花苜蓿生理特性的研究. 草地学报, 2012, 20(6): 1077-1080.
[3] Tang Z C. Response and adaptability of plant to water stress. Plant Physiology communications, 1983, 98(4): 1-7.
汤章程. 植物对水分胁迫的反应和适应性. 植物生理学通讯, 1983, 98(4): 1-7.
[4] Yang C Y, Zhong L, Li C Q,etal. Research progress of tall fescue in genetic breeding. Guizhou Agricultural Science, 2013, 41(6): 13-16.
杨春燕, 钟理, 李辰琼, 等. 高羊茅遗传育种研究进展. 贵州农业科学, 2013, 41(6): 13-16.
[5] Zhou F, Effect of different treatment on four tall fescue varieties seed germination. Southwest Horticulture, 2006, 2: 25-26.
邹芬. 不同处理对四个高羊茅品种种子发芽的影响. 西南园艺, 2006, 2: 25-26
[6] Wang Y, Guo H L, Chen J B,etal. Preliminary evaluation of drought resistance for the hybrid zoysiagrass ‘Suzhi No.1’. Acta Prataculturae Sinica, 2016, 25(5): 30-39.
汪毅, 郭海林, 陈静波, 等. 国审品种‘苏植1号’杂交结缕草抗旱性初步评价与分析. 草业学报, 2016, 25(5): 30-39.
[7] Yang H S, Chang G Z, Zhou X H. Performance of Hangmu No.1 alfalfa in the Lanzhou region. Acta Prataculturae Sinica, 2015, 24(9): 138-145.
杨红善, 常根柱, 周学辉. 航天诱变航苜1号紫花苜蓿兰州品种比较试验. 草业学报, 2015, 24(9): 138-145.
[8] Xu Y Y, Jia J F, Niu B T. The influences of space condition on three legume forages. Chinese Journal of Space Science, 1996, 16(Suppl): 136-141.
徐云远, 贾敬芬, 牛炳韬. 空间条件对3种豆科牧草的影响. 空间科学学报, 1996, 16(增刊): 136-141.
[9] Han L, Sun Z Y, Qian Y Q,etal. Change in biological characteristics ofPoapratensiscarried by Shenzhou-3 spaceship. Pratacultural Science, 2004, 21(4): 17-19.
韩蕾, 孙振元, 钱永强, 等. 神州三号飞船搭载对草地早熟禾生物学特性的影响. 草业科学, 2004, 21(4): 17-19.
[10] Hu H G, Liu J X, Guo H L. Spaceflight mutation breeding of china and its application in grass. Acta Prataculturae Sinica, 2006, 15(1): 15-21.
胡化广, 刘建秀, 郭海林. 我国植物空间诱变育种及其在草类植物育种中的应用. 草业学报, 2006, 15(1): 15-21.
[11] Zhang Y Q, Jia W L, Yang L L,etal. A study on character mutation of60Co ray irradiation inFestucaarundinacea. Acta Prataculturae Sinica, 2005, 14(4): 65-71.
张彦芹, 贾炜珑, 杨丽莉, 等.60Co辐射高羊茅性状变异研究. 草业学报, 2005, 14(4): 65-71.
[12] Xuan J P, Guo A G, Liu J X,etal. Analysis on morphological vaniation of mutants ofCynodondactyloninduced by radiation. Acta Prataculturae Sinica, 2005, 14(6): 107-111.
宣继萍, 郭爱桂, 刘建秀, 等. 狗牙根辐射诱变后代外部性状变异分析. 草业学报, 2005, 14(6): 107-111.
[13] Duther F R,Hess E L,Halstead R W.Progress in plant research in space. Advances in Space Research, 1994, 14(8): 159-162.
[14] Li H S, The Principles and Techniques of Plant Physiology and Biochemistry Experiment[M]. Beijing: Higher Education Press, 2000: 109-261.
李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000: 109-261.
[15] Huang C L, Chen X, Gao G L. Physiological response of seedlings of three azales species of drought stress and ebaluation of drought resistance. Scientia Silvae Sinicae, 2011, 47(6): 48-55.
黄承玲, 陈训, 高贵龙. 3种高山杜鹃对持续干旱的生理响应及抗旱性评价. 林业科学, 2011, 47(6): 48-55.
[16] Wei Y S, Liang Z S, Shan L. Comprehensive evaluation on alfalfa drought resistance traits by subordinate function values analysis. Pratacultural Science, 2005, 22(6): 33-36.
魏永胜, 梁宗锁, 山仑, 等. 利用隶属函数值法评价苜蓿抗旱性. 草业科学, 2005, 22(6): 33-36.
[17] Zhong P F, Su S P, Li Y. Comprehensive evaluation of drought resistance ofReaumuriasoongoricafrom four geographical populations. Acta Prataculturae Sinica, 2011, 20(5): 26-33.
种培芳, 苏世平, 李毅. 4个地理种群红砂的抗旱性综合评价. 草业学报, 2011, 20(5): 26-33.
[18] Zhao L L, Chen C, Wang P C. A comprehensive assessment of drought resistance ofLotuscorniculatusduring seed germination stage. Guizhou Agricultural Sciences, 2012, 40(9): 56-60.
赵丽丽, 陈超, 王普昶. 百脉根种子萌发期抗旱性综合评价. 贵州农业科学, 2012, 40(9): 56-60.
[19] Marie Pierre R, Vartavian N, Sallantin M,etal. Characterization of a novel protein induced by progressive or rapid drought and salinity inBrassicanapusleaves. Plant Physiology, 1992, 100: 1486-1493.
[20] Huang C B, Zeng F J, Lei J Q. Comparison of drought resistance among three Calligonum in the Southern Fringe of the Taklamakan Desert. Acta Prataculturae Sinica, 2014, 23(3): 136-143.
黄彩变, 曾凡江, 雷加强. 塔克拉玛干沙漠南缘3个沙拐枣种的抗旱特性比较. 草业学报, 2014, 23(3): 136-143.
[21] Liu E E, Wang P H, Guo Z F. Drought-induced proteins in plants. Plant Physiology Communications, 2001, 37(2): 155-160.
刘娥娥, 汪沛洪, 郭振飞. 植物的干旱诱导蛋白. 植物生理学通讯, 2001, 37(2): 155-160.
[22] Wu Z H, Zeng F H, Ma S J,etal. Effects of exogenous ABA on soluble protein inCynodondactylonunder PEG stress. Acta Prataculturae Sinica, 2004, 13(5): 75-78.
吴志华, 曾富华, 马生健, 等. ABA对PEG胁迫下狗牙根可溶性蛋白质的影响. 草业学报, 2004, 13(5): 75-78.
[23] Xiong L M, Schumaker K S, Zhu J K. Cell signaling during sold drought and salt stress. Plant Cell, 2002, 14: 165-183.
[24] Han J M. The responses of rice seedlings to the osmotic stress and relation to the osmotic adjustment. Journal of Agricultural University of Hebei, 1990, 13(1): 17-21.
韩建民. 抗旱性不同的水稻品种对渗透胁迫的反应及其与渗透调节的关系. 河北农业大学学报, 1990, 13(1): 17-21.
[25] Li D Q, Zou Q, Cheng B S. Osmotic adjustment and osmotica of wheat cultivars with different drought resistance under soil drought. Acta Phytophysiologica Sinica, 1992, 18(1): 37-43.
李德全, 邹琦, 程炳嵩. 土壤干旱下不同抗旱性小麦品种的渗透调节和渗透调节物质. 植物生理学报, 1992, 18(1): 37-43.
[26] Zhao T H, Shen X Y, Yang D G,etal. Effect of water stress on proteins in seedlings leaves of different drought-resistant Maize Hybrids. Journal of Shenyang Agricultural University, 2002, 33(6): 408-410.
赵天宏, 沈秀瑛, 杨德光, 等. 水分胁迫对不同抗旱性玉米幼苗叶片蛋白质的影响. 沈阳农业大学学报, 2002, 33(6): 408-410.
[27] Ma Y J, Ma R, Cao Z Z,etal. Effects of PEG stress on physiological characteristics of theLespedezaseedling leaves. Journal of Desert Research, 2012, 32(6): 1662-1668.
马彦军, 马瑞, 曹致中, 等. PEG胁迫对胡枝子幼苗叶片生理特性的影响. 中国沙漠, 2012, 32(6): 1662-1668.
[28] Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, 2010, 48(12): 909-930.
[29] Yao Y C, Wang Y N, Zhao X F. Study on the charaters of SOD eneymes in seedling of date plum persimmon under different low teperature. Journal of Beijing Agricultural College, 1994, 9(2): 49-55.
姚允聪, 王有年, 赵晓芳. 低温条件下君迁子幼苗SOD同工酶谱带与活性研究. 北京农学院学报, 1994, 9(2): 49-55.
[30] Hu Y B, Cheng J. Research on alleopathy of aqueous extract fromAstragalusstrictus. Acta Prataculturae Sinica, 2013, 22(6): 136-142.
胡远彬, 程俊. 茎直黄芪水浸提液化感作用研究. 草业学报, 2013, 22(6): 136-142.
[31] Zou C J, Han S J, Xu W D,etal. Eco-physiological responses ofPrceamongolicaecotypes to drought stress. Chinese Journal of Applied Ecology, 2003, 14(9): 1446-1450.
邹春静, 韩士杰, 徐文铎, 等. 沙地云杉生态型对干旱胁迫的生理生态响应. 应用生态学报, 2003, 14(9): 1446-1450.
[32] Aygul Abduwayit, Wang Z, Muhtar Zari,etal. Effect of drought stress on plant growth and photosynthetic characteristics of three alfalfa varieties. Chinese Agricultural Science Bulletin, 2016, 32(14): 7-12.
阿衣古力·阿布都瓦依提, 王铮, 木合塔尔·扎热, 等. 干旱胁迫对3个苜蓿品种的生长及光合特性的影响. 中国农学通报, 2016, 32(14): 7-12.
[33] Sun L X, Sun P, Cai S Z,etal. Physiological responses of four cold-season turfgrasses to oxidative stress of SO2. Acta Prataculturae Sinica, 2014, 23(4): 237-244.
孙凌霞, 孙萍, 蔡仕珍, 等. 四种冷季型草坪草对二氧化硫胁迫的生理响应研究. 草业学报, 2014, 23(4): 237-244.
[34] Xue X D, Dong X Y, Duan Y X,etal. A comparison of salt resistance of three kinds ofZoysiaat different salt concentrations. Acta Prataculturae Sinica, 2013, 22(6): 315-320.
薛秀栋, 董晓颖, 段艳欣, 等. 不同盐浓度下3种结缕草的耐盐性比较研究. 草业学报, 2013, 22(6): 315-320.
[35] Dasgupta J, Beweley J D. Variations in protein synthesis in different regions of greening leaves of barley seedling and effects of imposed water stress. Journal of Experimental Botany, 1984, 35: 1450-1459.
EffectsofdroughtstressonthephysiologicalcharacteristicsofnewlinesofFestucaarundinaceainducedbyspaceflightandtheircomprehensiveevaluation
LI Juan, LEI Xia, WANG Xiao-Li, MOU Qiong, YANG Chun-Yan, WU Jia-Hai*
PratacultureInstituteofGuizhouacademyofAgriculturalSciences,Guiyang550006,China
In this study, the ecophysiological characteristics of 14 lines ofFestucaarundinacea, including some induced by spaceflight, were studied in a pot experiment. The plants were subjected to drought stress conditions, and physiological and biochemical indices including soluble protein content, chlorophyll (Chl) content, and the activities of superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) activity were determined. These indexes of the 14F.arundinaceawere used to quantitatively evaluate their drought tolerance. The soluble protein content and Chl content increased, and the activities of SOD, POD, and CAT decreased under drought conditions. The drought resistance of the 14F.arundinacealines was calculated using the subordinate function, and the lines were ranked, from most drought tolerant to least drought tolerant, as follows: SP5-32>SP5-75>F.arundinaceacv. Shuicheng>SP5-60>SP5-94>SP5-97>SP5-89>SP5-71>SP5-7>SP5-85>SP5-42>SP5-5>F.arundinaceacv. Qiancao No.1>SP5-88. The average subordinate function of SP5-32, the most drought tolerant line, was 0.533. A grey correlative analysis was conducted to determine which indices were most strongly related to drought resistance. The indices were ranked, from strongest correlation with drought resistance to weakest, as follows: POD activity>soluble protein content>SOD activity>catalase activity>Chl content (correlation coefficients, 0.55-0.65). Therefore, SP5-32 is more suitable for cultivation in drought-affected areas as its antioxidant enzyme activity, soluble protein content, and chlorophyll content are unaffected by drought stress.
Festucaarundinacea; drought stress; space mutation; physiological characters; comprehensive evaluation
10.11686/cyxb2016502http//cyxb.lzu.edu.cn
李娟, 雷霞, 王小利, 牟琼, 杨春燕, 吴佳海. 干旱胁迫对高羊茅航天诱变新品系生理特性的影响及综合评价. 草业学报, 2017, 26(10): 87-98.
LI Juan, LEI Xia, WANG Xiao-Li, MOU Qiong, YANG Chun-Yan, WU Jia-Hai. Effects of drought stress on the physiological characteristics of new lines ofFestucaarundinaceainduced by spaceflight and their comprehensive evaluation. Acta Prataculturae Sinica, 2017, 26(10): 87-98.
2016-12-28;改回日期:2017-04-19
贵州省联合基金(黔科合J字LKN[2013]24号),贵州省重大专项(黔科合重大专项字[2014]6017号)和贵州省高层次创新型人才培养(黔科合人才[2016]4024号)资助。
李娟(1982-),女,甘肃平凉人,副研究员,硕士。E-mail: xixingyue@126.com
*通信作者Corresponding author. E-mail: 1848266168@qq.com
