毛竹向杉木林扩展过程中叶功能性状的适应策略*
2017-10-14刘广路范少辉唐晓鹿刘希珍
刘广路 范少辉 唐晓鹿 刘希珍
(国际竹藤中心 竹藤科学与技术重点实验室 北京 100102)
毛竹向杉木林扩展过程中叶功能性状的适应策略*
刘广路 范少辉 唐晓鹿 刘希珍
(国际竹藤中心 竹藤科学与技术重点实验室 北京 100102)
【目的】 揭示毛竹和杉木叶功能性状变化规律,探索毛竹林扩展过程中毛竹和杉木叶片的适应特性和生存对策,为毛竹林合理调控与生态经营提供依据。【方法】 沿毛竹向杉木林扩展方向设置3个10 m×50 m调查样带,每一样带平均划分为10个5 m×10 m样方,在每个样方内选取不同年龄的标准毛竹和杉木,选取叶片测定比叶面积(SLA)、叶干物质含量(LDMC)及叶片中碳(C)、氮(N)、磷(P)含量,并计算其比值。【结果】 1) 毛竹与杉木比叶面积(除Ⅰ度竹)与毛竹比例呈负相关关系,干物质含量与毛竹比例呈正相关关系。Ⅰ、Ⅱ、Ⅲ和Ⅳ度平均比面积分别为291、215、207、213 m2·kg-1,平均干物质含量分别为0.38、0.40、0.42、0.42 g·g-1,均显著高于杉木(115 m2·kg-1与0.34 g·g-1,P< 0.05)。2) Ⅰ度竹和Ⅳ度竹叶片N、P含量随着毛竹比例增加呈增加的趋势,Ⅱ、Ⅲ度竹和杉木叶片N、P含量呈降低的趋势。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均叶碳含量分别为46.5%、46.1%、45.5%和46.0%,氮含量分别为27.6、22.3、21.7和20.8 g·kg-1,磷含量分别为1.6、1.4、1.2和1.2 g·kg-1。其中,毛竹叶氮、磷含量显著高于杉木(分别为13.6和1.0 g·kg-1)。3) 毛竹SLA,LDMC与叶C、N、P含量,C∶N,N∶P等性状之间存在显著或极显著相关关系。【结论】 毛竹叶片具有比杉木叶片更高的SLA和叶N、P含量,且其向杉木扩展过程中,不同年龄毛竹的SLA,LDMC与叶C、N、P含量及C∶N,N∶P等叶功能性状采取的适应策略不同,具有互补效应,能够比杉木更有效地利用资源。
扩展; 比叶面积; 叶片养分含量; 干物质含量; 适应策略
Abstract: 【Objective】 This study aims to reveal the changing patterns of the leaf functional traits of moso bamboo and Chinese fir, and explore the adaptability and survival strategy of moso bamboo and Chinese fir during the expansion of moso bamboo to Chinese fir forest. 【Method】 Three 10 m×50 m transects were set along the expansion direction from moso bamboo to Chinese fir plantation, and each transect was divided into 10 subplots (a size of 5 m×10 m). In each subplot, standard culms/stems of moso bamboo at different ages and Chinese fir were selected. Leaves were collected on these trees to measure the specific leaf area (SLA), leaf dry matter content (LDMC), leaf carbon content (C), leaf nitrogen content (N), leaf phosphorus content (P) and calculate the ratios. 【Result】 1) During the expansion process of moso bamboo to Chinese fir, SLA of moso bamboo and Chinese fir was significantly negatively related with the ratio of moso bamboo, except Ⅰ du bamboo, while LDMCs were significantly positively related with the ratio of moso bamboo. The mean SLAs of Ⅰ, Ⅱ, Ⅲ and Ⅳ bamboo were 291, 215, 207 and 213 m2·kg-1, respectively, and the LDMC of Ⅰ, Ⅱ, Ⅲ and Ⅳ bamboo was 0.38, 0.40, 0.42 and 0.42 g·g-1, respectively. The two parameters were significantly higher than those of Chinese fir (115 m2·kg-1for SLA and 0.34 g·g-1for LDMC,P< 0.005). 2) For Ⅰ and Ⅳ bamboo, leaf C, N and P contents of moso bamboo showed an increasing trend with the ratio of moso bamboo. Meanwhile, a decreasing trend with the ratio of moso bamboo was observed for Ⅱ, Ⅲ and Chinese fir leaves. Leaves of Ⅰ, Ⅱ, Ⅲ and Ⅳ had 46.5%, 46.1%, 45.5% and 46.0% C contents, respectively, 27.6, 22.3, 21.7 and 20.8 g·kg-1N content, and 1.6, 1.4, 1.2, and 1.2 g·kg-1for P content. The N and P contents of moso bamboo were significantly higher than those of Chinese fir (13.6 and 1.0 g·kg-1, respectively). 3) There were significant or extreme significant relationships between bamboo SLA, LDMC and leaf C, N, P contents, C∶N and N∶P. 【Conclusion】 Compared to Chinese fir, moso bamboo had larger SLA, and higher N and P contents, and during its expansion to Chinese fir, the main leaf functional traits of moso bamboo at different ages adapted differently with a complementary effect, which can make use of the resources more effectively.
Keywords: expansion; specific leaf area; nutrient content in leaf; dry matter content in leaf; survival strategy
叶片是植物进行光合作用和物质生产的主要器官,对环境变化极为敏感。叶功能性状是植物资源利用策略的指标,可反映不同物种或种源在长期进化过程中对生存环境的适应和竞争能力(Liuetal., 2013),并随着立地条件、温度、降水等环境因子的变化而发生变化(Wrightetal., 2005)。目前,国内外对植物叶功能性状的研究主要集中在不同物种的叶功能性状特征(任书杰等, 2012; Hanetal., 2005; 王晶苑等, 2011; Xiaetal., 2014)、环境因子对植物叶性状的影响(安海龙等, 2015; 杨冬梅等, 2012)、植物性状对干扰的响应(李西良等, 2014)等方面,其中Wright等(2004)提出的植物叶经济型谱(主要是叶片的化学、结构以及生理特征)是最具影响力的成果(陈莹婷等, 2014)。当前,有关叶功能性状的研究基本涵盖了不同尺度的代表性乔木、灌木和草本植物(毛伟等, 2012),但有关毛竹(Phyllostachyseduis)叶功能性状的报道较少。
毛竹属于单轴散生型竹种,具有向周边环境扩展的强大能力(施建敏等, 2014)。毛竹林扩展在提供更多竹资源的同时,也出现了植物群落物种多样性降低等现象(白尚斌等, 2013; 林倩倩等, 2014; 杨清培等, 2015),开展毛竹扩展策略研究,可为合理调控毛竹林扩展过程提供科学依据。当前,关于毛竹扩展对原生植被和土壤的影响以及毛竹扩展过程中的适应策略等方面已有一些研究(吴家森等, 2008; 宋庆妮等, 2013),毛竹林在扩展过程中,土壤养分含量发生显著变化,这些变化通常会导致林分生产力下降。在毛竹向阔叶林扩展过程中,毛竹通过增加细根生长速率和周转率可获得更强的竞争力(刘骏等, 2013),同时毛竹叶功能性状也发生了一系列适应性改变以适应变化的环境条件(刘希珍等, 2015)。杉木(Cunninghamialanceolata)作为我国最重要的人工林树种之一,其适宜生长的立地条件与毛竹基本一致,在生产中存在着很多由毛竹扩展形成的竹杉混交林,但关于毛竹向杉木林扩展策略的研究较少,史纪明等(2013)对不同扩展方向毛竹的大小和密度进行了分析,刘烁等(2011)研究了毛竹、竹-杉和杉木林光照变化规律,毛竹向杉木扩展过程中毛竹、杉木叶功能性状的研究未见报道。
表征植物叶功能性状的指标通常有植物比叶面积(specific leaf area,SLA)、叶干物质含量(leaf dry mass content,LDMC)和叶片C、N、P含量及比值,其中,植物比叶面积和叶干物质含量是最重要的指标,可以解释90%以上植物叶片性状随环境变化而发生的变异(Rocheetal.,2004)。本研究以毛竹-杉木扩展界面为研究对象,对毛竹和杉木叶的比叶面积、叶干物质含量和叶片主要养分元素及计量比值进行研究,以阐明毛竹扩展过程中杉木和毛竹叶片的叶功能性状变化规律,揭示毛竹向杉木林扩展策略,为毛竹林合理调控与生态经营提供科学依据。
1 研究区概况
研究区位于福建永安竹林生态系统定位观测研究站天宝岩国家级自然保护区观测点(117°28′03″—117°35′28″E, 25°50′51″—26°01′20″N),属于戴云山余脉,中低山地貌,海拔高580~1 604.8 m,中亚热带东南季风气候。年均气温15 ℃,绝对最低温度-11 ℃,绝对最高温度40 ℃; 年均相对湿度80%以上,无霜期290天左右,土壤主要为红壤。大部分毛竹林是自然扩展形成的,其间混杂少量马尾松(Pinusmassoniana)、杉木,竹杉混交林由毛竹经过近15年向杉木林扩展形成,杉木林林龄约40年,其中伴生有极少量木荷(Schimasuperba)等阔叶树种。
2 研究方法
2.1试验设计
沿毛竹向杉木林扩展方向设置3个10 m×50 m调查样带,每一样带平均划分为10个5 m×10 m样方,扩展前沿(杉木纯林)编号为样方1,随着毛竹比例增加,依次编号为样方2、样方3、…、样方10。从样方1到样方10毛竹所占比例分别为0、15.8%、32.6%、33.7%、38.0%、52.2%、64.5%、78.7%、78.9%和100%。本研究中毛竹比例为样方内毛竹胸高断面积与林分总胸高断面积的比值。样带的基本情况见表1。
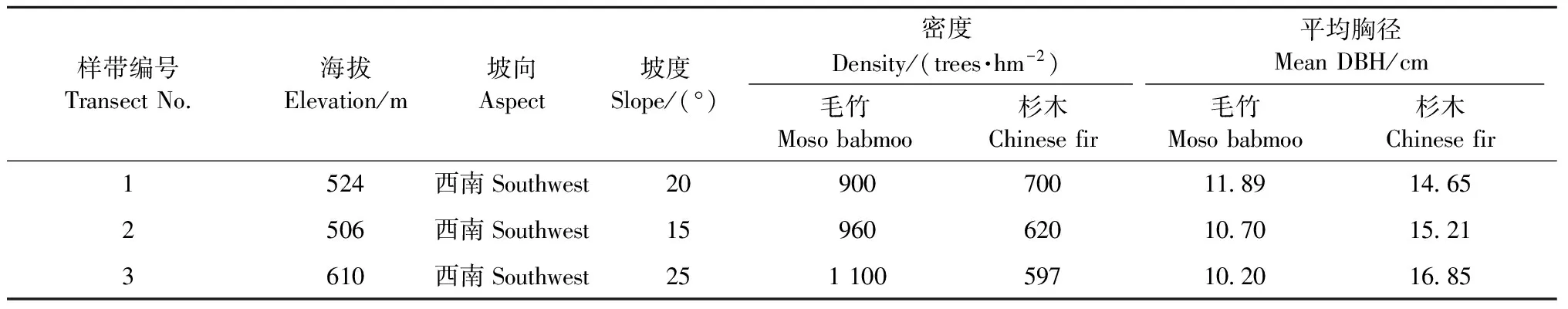
表1 样带基本情况Tab.1 Basic characteristics of the transects
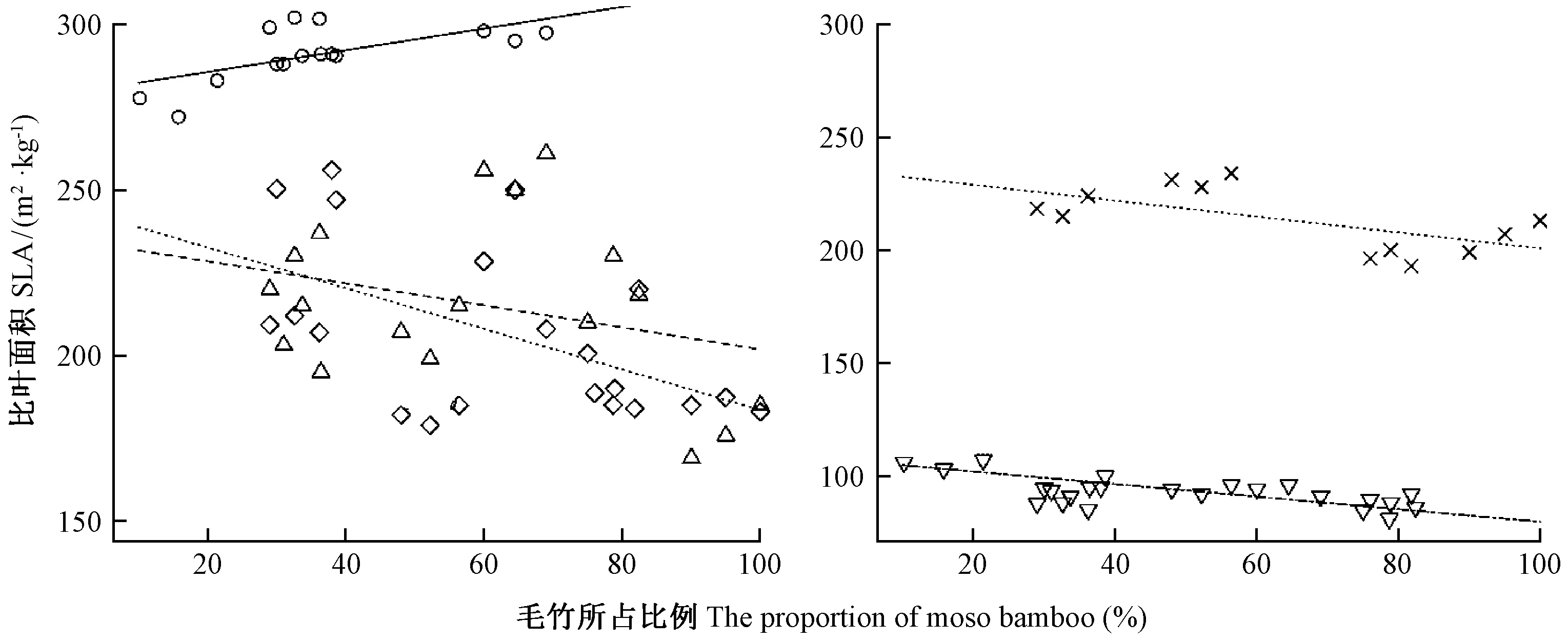
图1 毛竹向杉木林扩展过程中毛竹与杉木比叶面积的变化Fig.1 Changes of SLA of moso bamboo and Chinese fir forests during the expansion from moso bamboo forests to Chinese fir forestsⅠ度竹(R2=0.41,P=0.01),Ⅱ度竹(R2=0.09,P=0.222),Ⅲ度竹(R2=0.29,P=0.012),Ⅳ度竹(R2=0.39,P=0.029),杉木(R2=0.580,P<0.001)。Ⅰ du bamboo(R2=0.41,P=0.01),Ⅱ du bamboo(R2=0.09,P=0.222),Ⅲ du bamboo(R2=0.29,P=0.012),Ⅳ du bamboo(R2=0.39,P=0.029),Chinese fir(R2=0.580,P<0.001).
2.2试验方法
2.2.1 叶片选取 在每个样带1~10号样方内选取平均胸径和树高的标准竹,Ⅰ、Ⅱ、Ⅲ、Ⅳ度竹各选1株,从东南西北4个方向采集竹冠中部健康叶片,每方向至少选10片。然后将所采集叶片立即置于2片湿润的滤纸之间,放入塑封袋,编号、带回实验室。因为毛竹林扩展过程中,不同年龄毛竹分布不均匀,Ⅰ度竹在1~8号样方取得了合适的样品,Ⅱ度竹在4~10号样方取得了合适的样品,Ⅲ度竹在3~10号样方取得了合适的样品,Ⅳ度竹在6~10号样方,取得了合适的样品。杉木叶片为针叶,取50片称重后平铺于白纸上。与毛竹叶片采集类似,在杉木标准木上采集约50片叶片带回实验室分析。
2.2.2 比叶面积(SLA)和叶干物质含量(LDMC)测定 将塑封袋内的叶片去掉叶柄并浸泡于水中,在5 ℃黑暗环境中放置12 h,然后用吸水纸吸去叶表面水分,用天平(德国赛多利斯QUINTIX213-1CN,1 mg)称饱和鲜质量。用扫描仪扫描叶片,并采用Image J软件计算叶面积。将叶片于65 ℃烘箱内烘至恒质量,取出称重即得叶干质量。SLA=叶面积/叶干质量; LDMC=叶干质量/饱和鲜质量(刘希珍等, 2015)。
2.2.3 叶片养分含量测定 将叶片恒温65 ℃下烘至恒质量后研磨,过0.149 mm土壤筛。叶片有机碳(C)采用高温外热重铬酸钾氧化-容量法测定,全氮采用凯式定氮法测定,全磷为钒钼黄比色法测定(张万儒等, 1999)。
2.3数据处理
叶功能性状差异进行one-way ANOVA的差异显著性检验,差异达到显著水平(P< 0.05)时,进行多元比较分析(LSD法); 相关分析采用Correlate-Bivariate法; 数据处理与图表制作在R语言中完成。
3 结果与分析
3.1比叶面积(SLA)变化
随着毛竹的扩展,毛竹和杉木比叶面积发生了规律性变化(图1)。毛竹比叶面积介于169 m2·kg-1(Ⅱ度竹)~302 m2·kg-1之间(Ⅰ度竹),其中Ⅰ度竹的比叶面积显著高于Ⅱ、Ⅲ和Ⅳ度竹(P< 0.001),但Ⅱ、Ⅲ、Ⅳ度竹比叶面积的差异未达到显著水平(P> 0.580)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均比叶面积分别为291、215、207和213 m2·kg-1。与毛竹比面积相比,杉木比叶面积显著低于毛竹林比叶面积(P< 0.001),杉木比叶面积最高为115 m2·kg-1(纯林),最低为87 m2·kg-1(毛竹比例达到70%~80%),平均为95 m2·kg-1。随着毛竹向杉木林扩展,毛竹与杉木林比叶面积呈不同的变化趋势。Ⅰ度竹比叶面积随着毛竹比例增加呈显著增加的趋势(R2=0.41,P= 0.010); Ⅲ、Ⅳ度竹和杉木比叶面积随着毛竹比例增加呈显著降低的趋势(R2> 0.29,P<0.03),但Ⅱ度竹比叶面积随毛竹比例增加并无显著变化(R2=0.09,P= 0.222)。
3.2叶干物质含量(LDMC)变化
不同年龄毛竹林叶干物质含量介于0.33~0.58 g·g-1之间(图2),其差异达到显著水平(P= 0.001)。其中Ⅰ度竹叶干物质含量最小为0.38 g·g-1,显著低于Ⅲ度竹(0.42 g·g-1,P= 0.001)与Ⅳ度竹(0.42 g·g-1,P= 0.005),但Ⅱ度竹(0.40 g·g-1)与Ⅰ、Ⅲ和Ⅳ竹度差异并未达到显著水平(P> 0.128)。杉木林叶干物质含量介于0.25~0.44 g·g-1之间,平均为0.33 g·g-1,显著低于毛竹林(P< 0.001)。随着毛竹向杉木林扩展,毛竹与杉木干物质含量显现出规律性变化,即随着毛竹比例增加,叶干物质含量呈增加的趋势(P≤ 0.045)。

图2 毛竹向杉木林扩展过程中毛竹和杉木叶干物质含量的变化Fig.2 Changes of LDMC of moso bamboo and Chinese fir forests during the expansion from moso bamboo forests to Chinese fir forestsⅠ度竹(R2=0.39,P=0.013),Ⅱ度竹(R2=0.35,P=0.010),Ⅲ度竹(R2=0.40,P=0.002),Ⅳ度竹(R2=0.39,P=0.029),杉木(R2=0.28,P<0.004)。Ⅰ du bamboo(R2=0.39,P=0.013),Ⅱ du bamboo(R2=0.35,P=0.010),Ⅲ du bamboo(R2=0.40,P=0.002),Ⅳ du bamboo(R2=0.39,P=0.029),Chinese fir (R2=0.28,P<0.004).
3.3叶片养分含量及计量比值变化
3.3.1 养分含量的变化 1) 碳含量的变化 随着毛竹向杉木林扩展,杉木和不同年龄毛竹叶片的碳含量差异不大,为41%~53%(图3a,b)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹叶碳含量差异不显著(P> 0.565),平均碳含量分别为46.5%、46.1%、45.5%和46.0%。杉木叶碳含量为49.8%。随着毛竹比例增加,毛竹与杉木叶碳含量均未表现出明显的变化趋势(P≥ 0.05)。
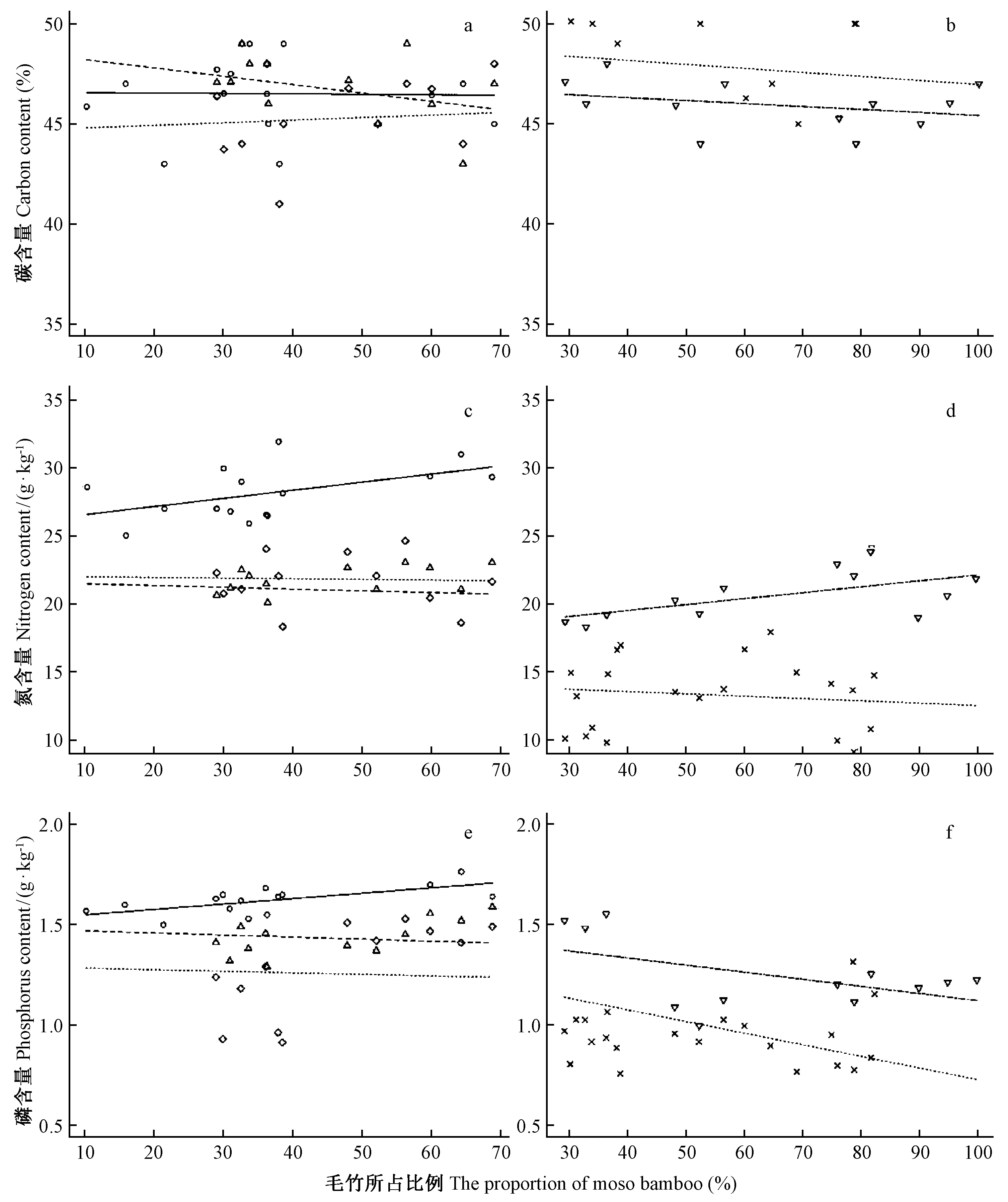
图3 毛竹扩展过程中杉木和不同年龄毛竹叶碳(a,b),氮(c,d)和磷(e,f)含量的变化Fig.3 Changes of leaf carbon (a, b), nitrogen (c, d) and phosphorus (e, f) contents of moso bamboo and Chinese fir forests during the expansion from moso bamboo forests to Chinese fir forests图a和图b中Ⅰ度竹(R2=0.00,P=0.947),Ⅱ度竹(R2=0.22,P=0.050),Ⅲ度竹(R2=0.03,P=0.474),Ⅳ度竹(R2=0.09,P=0.338),杉木(R2=0.05,P=0.278). 图c和图d中Ⅰ度竹(R2=0.25,P=0.056),Ⅱ度竹(R2=0.01,P=0.719),Ⅲ度竹(R2=0.00,P=0.788),Ⅳ度竹(R2=0.38,P=0.033),杉木(R2=0.02,P=0.434). 图e和图f中Ⅰ度竹(R2=0.43,P=0.008),Ⅱ度竹(R2=0.02,P=0.564),Ⅲ度竹(R2=0.01,P=0.729),Ⅳ度竹(R2=0.25,P=0.098),杉木(R2=0.38,P<0.001)。In Fig.3a and Fig.3b,Ⅰ du bamboo(R2=0.00,P=0.947),Ⅱ du bamboo(R2=0.22,P=0.050),Ⅲ du bamboo(R2=0.03,P=0.474),Ⅳ du bamboo(R2=0.09,P=0.338),Chinese fir(R2=0.05,P=0.278). In Fig.3c and Fig.3d,Ⅰ du bamboo(R2=0.25,P=0.056),Ⅱ du bamboo(R2=0.01,P=0.719),Ⅲ du bamboo(R2=0.00,P=0.788),Ⅳ du bamboo(R2=0.38,P=0.033),Chinese fir(R2=0.02,P=0.434). In Fig.3e and fig.3f,Ⅰ du bamboo(R2=0.43,P=0.008),Ⅱ du bamboo(R2=0.02,P=0.564),Ⅲ du bamboo(R2=0.01,P=0.729),Ⅳ du bamboo(R2=0.25,P=0.098),Chinese fir(R2=0.38,P<0.001).
2) 氮含量的变化 毛竹叶氮含量介于17.0-32.0 g·kg-1之间(图3c,d),不同年龄毛竹叶氮含量差异达到显著水平(P< 0.001)。Ⅰ度竹叶氮含量显著高于Ⅱ、Ⅲ和Ⅳ度竹(P< 0.001),但Ⅱ、Ⅲ、Ⅳ度竹叶氮含量差异未达到显著水平(P> 0.522)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均叶氮含量分别为27.6、22.3、21.7和20.8 g·kg-1,随着年龄增加呈降低的趋势。杉木氮含量在8.6~15.6 g·kg-1之间,显著低于毛竹叶氮含量(P< 0.001),平均为13.6 g·kg-1。不同年龄毛竹与杉木叶氮含量随着毛竹比例增加呈不同变化趋势。Ⅰ和Ⅳ度竹叶氮含量随着毛竹比例的增加而呈增加的趋势(P≤ 0.056),但Ⅱ、Ⅲ和杉木叶氮含量随着毛竹比例增加而无显著变化(P> 0.434)。
3) 磷含量的变化 毛竹叶磷含量介于0.9~1.8 g·kg-1之间,并随着毛竹年龄增加呈降低的趋势(图3e,f)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均叶磷含量分别为1.6、1.4、1.2和1.2 g·kg-1。Ⅰ度竹叶磷含量显著高于Ⅱ、Ⅲ和Ⅳ度竹(P< 0.026),且Ⅱ度竹叶磷含量显著高于Ⅲ度竹(P=0.039),但与Ⅳ度竹差异不显著(P=0.106)。Ⅲ、Ⅳ度竹叶磷含量差异未达到显著水平(P=1.000)。杉木磷含量介于0.8~1.5 g·kg-1之间,显著低于毛竹叶磷含量(P≤ 0.036),平均为1.0 g·kg-1。不同年龄毛竹与杉木叶磷含量随着毛竹比例增加呈不同变化趋势。Ⅰ度竹叶磷含量随着毛竹比例增加呈增加的趋势(R2=0.43,P= 0.008),Ⅱ、Ⅲ和Ⅳ度毛竹叶磷含量随着毛竹比例增加无显著变化(P≥ 0.098)。杉木叶磷含量随着毛竹比例增加呈降低的趋势(R2=0.38,P< 0.001)。
3.3.2 C∶N和N∶P的变化 毛竹叶片C∶N介于13.4~26.5之间(图4a,b), Ⅰ度竹叶C∶N显著高于Ⅳ竹(P=0.045),但Ⅱ、Ⅲ和Ⅳ度竹C∶N差异不显著(P≥ 0.080)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均叶C∶N分别为17.1、21.1、21.1和22.2。杉木叶C∶N介于25.9~54.3之间,显著高于毛竹叶C∶N(P< 0.001),平均为38.3。不同年龄毛竹与杉木叶C∶N随着毛竹比例增加呈不同变化趋势。Ⅰ、Ⅱ和Ⅲ度竹叶C∶N随着毛竹比例增加无显著变化趋势(P≥ 0.136),而Ⅳ度竹叶C∶N随着毛竹比例增加呈降低趋势(P=0.017)。杉木叶C∶N随着毛竹比例增加无显著变化趋势(P= 0.385)。
毛竹叶片N∶P介于12.5~22.9之间(图4c,d), 不同年龄毛竹叶N∶P差异不显著(P> 0.152)。Ⅰ、Ⅱ、Ⅲ和Ⅳ度竹平均叶N∶P分别为17.0、15.7、17.8和17.1。杉木叶N∶P介于6.9~22.9之间,平均为13.2,显著低于Ⅰ、 Ⅲ和Ⅳ度竹而高于Ⅱ度竹叶N∶P(P< 0.003)。Ⅰ、Ⅱ、Ⅲ度竹和杉木叶N∶P随着毛竹比例增加无显著变化趋势(P> 0.095), Ⅳ竹叶N∶P随着毛竹比例增加呈增加趋势(P=0.045)。
3.4毛竹叶功能性状间的相关性分析
相关分析结果(表2)表明,SLA与LDMC、N、P、C∶N的相关性达到极显著水平,其中与LDMC和C∶N呈负相关,与叶片N、P含量呈正相关。SLA越大,LDMC含量低,叶片的N、P含量越高。LDMC与叶片P含量呈负相关,达到显著水平,与其他指标的相关性未达到显著水平; 叶片N含量与P含量、C∶N、N∶P的相关性也达到显著水平,其中与P含量和N∶P呈正相关,与C∶N呈负相关; 叶片P含量与叶片C含量呈正相关,与C∶N和N∶P呈负相关,相关性达到显著水平; 叶片C含量与N∶P、C∶N与N∶P显著负相关。

图4 毛竹扩展过程中杉木和毛竹叶片C∶N(a, b)和N∶P(c, d)的变化Fig.4 Changes of ratios of leaf C∶N (a, b) and N∶P (c, d) of moso bamboo and Chinese fir forests during the expansion from moso bamboo forests to Chinese fir forests图a和图b中Ⅰ度竹(R2=0.02,P=0.136),Ⅱ度竹(R2=0.01,P=0.720),Ⅲ度竹(R2=0.00,P=0.541),Ⅳ度竹(R2=0.45,P=0.017),杉木(R2=0.03,P=0.385). 图c和图d中Ⅰ度竹(R2=0.13,P=0.185),Ⅱ度竹(R2=0.16,P=0.095),Ⅲ度竹(R2=0.04,P=0.389),Ⅳ度竹(R2=0.35,P=0.045),杉木(R2=0.10,P=0.115)。In Fig.3a and Fig.3b,Ⅰ du bamboo(R2=0.02,P=0.136),Ⅱ du bamboo(R2=0.01,P=0.720),Ⅲ du bamboo(R2=0.00,P=0.541),Ⅳ du bamboo(R2=0.45,P=0.017),Chinese fir(R2=0.03,P=0.385). In Fig.3c and Fig.3d,Ⅰ du bamboo(R2=0.13,P=0.185),Ⅱ du bamboo(R2=0.16,P=0.095),Ⅲ du bamboo(R2=0.04,P=0.389),Ⅳ du bamboo(R2=0.35,P=0.045),Chinese fir(R2=0.10,P=0.115).
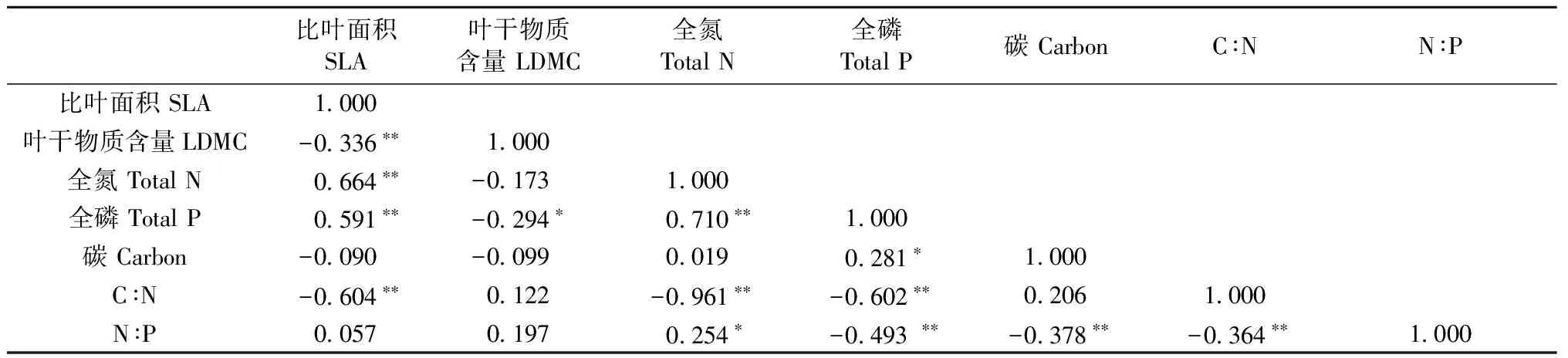
比叶面积SLA叶干物质含量LDMC全氮TotalN全磷TotalP碳CarbonC∶NN∶P比叶面积SLA1 000叶干物质含量LDMC-0 336∗∗1 000全氮TotalN0 664∗∗-0 1731 000全磷TotalP0 591∗∗-0 294∗0 710∗∗1 000碳Carbon-0 090-0 0990 0190 281∗1 000C∶N-0 604∗∗0 122-0 961∗∗-0 602∗∗0 2061 000N∶P0 0570 1970 254∗-0 493∗∗-0 378∗∗-0 364∗∗1 000
① SLA:species leaf area;LDMC:leaf dry mass content.*表示相关性达到显著水平(P<0.05),**表示相关性达到极显著水平(P<0.01)。Treatments with the symbol “*” indicateP< 0.05 based on correlation analysis using SPSS, and treatments with the symbol “**” indicateP< 0.01 based on correlation analysis using SPSS.
4 讨论
随着毛竹向杉木林扩展,毛竹和杉木的比叶面积发生了相应改变(图2)。比叶面积是表征植物对光能捕获能力和同化CO2能力的重要叶功能性状指标(Wilsonetal., 1999),杉木比叶面积随着毛竹比例增加而降低,反映了杉木的光能捕获和碳同化能力减弱,在与毛竹的竞争中逐渐处于不利地位(图2); 不同年龄毛竹比叶面积随着毛竹比例增加呈不同的变化趋势,使毛竹在不同扩展阶段都具有较高光能的捕获能力和同化CO2的能力,使其在竞争中占据有利地位。本研究中杉木的比叶面积显著低于毛竹的比叶面积,反映了2种植物环境适应策略不同,毛竹具有更大的同化能力(Cornelissenetal., 2003)。毛竹较高的比叶面积及其不同年龄毛竹在扩展过程中比叶面积变化趋势的多样性,使其能更有效地利用资源。随着毛竹向杉木林扩展,毛竹和杉木叶干物质含量随着毛竹的扩展呈上升趋势(图3),表明随着毛竹扩展,毛竹和杉木均通过增加某些资源的保有量来维持正常生长 (Qietal., 2008),且毛竹比杉木具有更好的养分保有能力。不同年龄毛竹叶干物质含量随着毛竹比例增加的变化幅度不同,且其通过地下茎互相连接,可以弥补单一年龄毛竹对某些资源获取能力不足的缺陷,获得较好的“集团”优势。
叶片的碳、氮、磷含量是衡量植物光合同化能力和生长发育的重要基础指标,其含量的比值能够较好地反映叶功能性状。毛竹和杉木叶碳含量随着毛竹比例增加变化趋势不明显,杉木叶片的碳含量高于毛竹叶片的碳含量,符合植物碳含量变化的一般规律(李斌等, 2015)。植物叶片氮是光合物质代谢和植物生长的关键性因子,是合成叶绿素和有关光合蛋白的重要成分(Evans, 1989),磷是核酸、磷脂的组分,对植物的光合能力具有重要影响。本研究中随着毛竹比例增加,Ⅰ和Ⅳ度竹叶氮含量增加,杉木叶氮含量无显著变化(图4c,d); Ⅰ度竹叶磷含量增加,杉木叶磷含量降低,可能预示着杉木的同化能力在降低,而毛竹的同化能力在增强。杉木叶N∶P为6.9~22.9,毛竹叶N∶P为12.5~22.9。叶片N∶P比在大约15时被认为是临界状况,当N∶P比大于16时,植物生长对P的响应强烈; 小于16时植物生长对N添加的响应强烈(Fuetal., 2007),反映了试验区毛竹和杉木的生长在不同扩展阶段受到N或者P的制约情况不同,杉木可能受N的制约更为明显(平均值为13.5),毛竹叶片N∶P(平均为16.99)更接近16的临近状态。
比叶面积与叶片干物质含量呈显著的负相关关系,与大部分研究结果(Wilsonetal., 1999;Wrightetal., 2001; Reichetal.,1991;朱媛君等, 2015)一致。叶干物质含量增加,比叶面积减小,使叶片内部水分向叶片表面扩散的距离或阻力增大,降低植物内部水分散失(Qietal., 2008)。比叶面积与全氮、全磷含量呈显著正相关关系,这种正相关关系基本上存在于所有植物中(Wrightetal., 2001; 郭茹等, 2015)。叶干物质含量和叶磷含量呈负相关,可能暗示着干物质含量高的植物叶片的光合同化能力相对较弱,与天山云杉(Piceaschrenkianavar.tianschanica)叶功能性状的相关性结果(张慧文等, 2010)一致。毛竹和杉木叶不同叶功能性状相关关系的差异,反映了其通过对不同性状的权衡来实现对资源的充分利用,进一步证明植物在不同的环境胁迫下可以通过不同性状的权衡来充分利用环境资源(李西良等, 2014)。
5 结论
随着毛竹向杉木林扩展,杉木比叶面积降低,叶干物质含量升高,磷含量降低,杉木的光捕获和碳同化能力逐渐降低,在与毛竹的竞争中逐渐处于弱势。毛竹叶片具有比杉木更高的比叶面积和叶N、叶P含量,毛竹具有更强的光捕获和碳同化能力。同时,不同年龄毛竹叶功能性状随毛竹林比例增加的变化趋势不同,使其能够更有效地利用资源,在与杉木的竞争中逐步处于有利地位。
安海龙, 谢乾瑾, 刘 超, 等. 2015.水分胁迫和种源对黄柳叶功能性状的影响. 林业科学, 51(10):75-84.
(An H L, Xie Q J, Liu C,etal. 2015.Effects of water stress and provenance on leaf functional traits ofSalixgordejevii. Scientia Silvae Sinicae, 51(10):75-84. [in Chinese])
白尚斌, 周国模, 王懿祥, 等.2013. 天目山保护区森林群落植物多样性对毛竹入侵的响应及动态变化.生物多样性, 21(3):288-295.
(Bai S B, Zhou G M, Wang Y X,etal. 2013.Plant species diversity and dynamics in forests invaded by moso bamboo (Phyllostachysedulis) in Tianmu Mountain Nature Reserve. Biodiversity Science, 21(3):288-295. [in Chinese])
陈莹婷,许振柱.2014.植物叶经济谱的研究进展.植物生态学报,38(10): 1135-1153.
(Chen Y T, Xu Z Z. 2014. Review on research of leaf economics spectrum. Chinese Journal of Plant Ecology,38(10): 1135-1153. [in Chinese])
郭 茹, 温仲明, 王红霞, 等. 2015.延河流域植物叶性状间关系及其在不同植被带的表达. 应用生态学报, 26(12):3627-3633.
(Guo R,Wen Z M,Wang H X,etal. 2015. Relationships among leaf traits and their expression in different vegetation zones in Yanhe River basin,northwest China.Chinese Journal of Applied Ecology, 26(12):3627-3633. [in Chinese])
李 斌, 方 晰, 田大伦, 等. 2015.湖南省现有森林植被主要树种的碳含量. 中南林业科技大学学报, 35(1):71-78.
(Li B, Fang X, Tian D L,etal. 2015. Studies on carbon concentration of main forest vegetation tree species in Hunan province. Journal of Central South University of Forestry & Technology, 5(1):71-78. [in Chinese])
李西良, 侯向阳, 吴新宏. 2014. 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应. 植物生态学报, 38(5):440-451.
(Li X L, Hou X Y, Wu X H,etal. 2014. Plastic responses of stem and leaf functional traits inLeymuschinensisto long-term grazing in a meadow steppe. Chinese Journal of Plant Ecology, 38(5):440-451. [in Chinese])
林倩倩, 王 彬, 马元丹, 等. 2014.天目山国家级自然保护区毛竹林扩张对生物多样性的影响.东北林业大学学报, 42(9): 43-47.
(Lin Q Q, Wang B, Ma Y D,etal. 2014. Effect ofPhyllostachyspubescensforest expansion on biodiversity in Tianmu Mountain National Nature Reserve. Journal of Northeast Forestry University, 42(9): 43-47. [in Chinese])
刘 骏, 杨清培, 宋庆妮, 等. 2013.毛竹种群向常绿阔叶林扩张的细根策略. 植物生态学报, 37(3):230-238.
(Liu J, Yang Q P, Song Q N,etal. 2013. Strategy of fine root expansion ofPhyllostachyspubescenspopulation into evergreen broadleaved forest. Chinese Journal of Plant Ecology,37(3):230-238. [in Chinese])
刘希珍, 封焕英, 蔡春菊, 等. 2015.毛竹向阔叶林扩展过程中的叶功能性状研究. 北京林业大学学报, 37(8):8-17.
(Liu X Z,Feng H Y,Cai C J,etal. 2015. Response of leaf functional traits of Moso bamboo during the invading process into the broad-leaved forest. Journal of Beijing Forestry University, 37(8): 8-17. [in Chinese])
刘 烁, 周国模, 白尚斌. 2011. 基于光照强度变化的毛竹扩张对杉木影响的探讨. 浙江农林大学学报, 28(4):550-554.
(Liu S, Zhou G M, Bai S B. 2011.Light intensity changes onCunninghamialanceolatain mixed stands with different concentrations ofPhyllostachyspubescens. Journal of Zhejiang A&F University,28(4):550-554. [in Chinese])
毛 伟, 李玉霖, 张铜会, 等.2012.不同尺度生态学中植物叶性状研究概述. 中国沙漠,32(1): 33-41.
(Mao W, Li Y L, Zhang T H,etal. 2012.Research advances of plant leaf traits at different ecology scale. Journal of Desert Research, 32(1): 33-41.[in Chinese])
任书杰,于贵瑞,姜春明,等.2012.中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征.应用生态学报, 23(3):581-586.
(Ren S J,Yu G R,Jiang C M,etal. 2012. Stoichiometric characteristics of leaf carbon,nitrogen,and phosphorus of 102 dominant species in forest ecosystems along the north-south transect of east China.Chinese Journal of Applied Ecology, 23(3):581-586. [in Chinese])
施建敏,叶学华,陈伏生,等. 2014.竹类植物对异质生境的适应——表型可塑性. 生态学报, 34(20):5687-5695.
(Shi J M,Ye X H,Chen F S,etal.2014.Adaptation of bamboo to heterogeneous habitat: phenotypic plasticity.Acta Ecologica Sinica, 34(20): 5687-5695.[in Chinese])
史纪明, 张纪林, 教忠意, 等. 2013.毛竹对杉木林入侵效应初步调查研究. 江苏林业科技, 40(1):7-9,41.
(Shi J M,Zhang J L, Jiao Z Y,etal.2013. Investigation ofPhyllostachyspubescensexpansion toCunninghamialanceolataforest. Journal of Jiangsu Forestry Science & Technology, 40(1):7-9,41. [in Chinese])
宋庆妮, 杨清培, 刘 骏, 等. 2013.毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响. 应用生态学报, 24(2):338-344.
(Song Q N,Yang Q P, Liu J,etal. 2013.Effects ofPhyllostachysedulisexpansion on soil nitrogen mineralization and its availability in evergreen broadleaf forest.Chinese Journal of Applied Ecology,24(2):338-344. [in Chinese])
王晶苑,王绍强,李纫兰,等.2011.中国四种森林类型主要优势植物的C∶N∶P化学计量学特征.植物生态学报, 35 (6): 587-595.
(Wang J Y, Wang S Q, Li R L,etal. 2011.C∶N∶P stoichiometric characteristics of four forest types’ dominant tree species in China. Chinese Journal of Plant Ecology, 35 (6): 587-595. [in Chinese])
吴家森, 姜培坤, 王祖良. 2008.天目山国家级自然保护区毛竹扩张对林地土壤肥力的影响. 江西农业大学学报, 30(4):689-692.
(Wu J S, Jiang P K, Wang Z L. 2008.The effects ofPhyllostachyspubescensexpansion on soil fertility in National Nature Reserve of Mount Tianmu. Acta Agriculturae Universitatis Jiangxiensis, 30(4):689-692. [in Chinese])
杨冬梅, 章佳佳, 周 丹, 等. 2012.木本植物茎叶功能性状及其关系随环境变化的研究进展. 生态学杂志, 31(3):702-713.
(Yang D M,Zhang J J,Zhou D,etal. 2012. Leaf and twig functional traits of woody plants and their relationships with environmental change: a review. Chinese Journal of Ecology, 31(3):702-713. [in Chinese])
杨清培, 杨光耀, 宋庆妮, 等. 2015.竹子扩张生态学研究:过程、后效与机制. 植物生态学报, 39(1):110-124.
(Yang Q P, Yang G Y, Song Q N,etal. 2015.Ecological studies on bamboo expansion: process, consequence and mechanism. Chinese Journal of Plant Ecology,39(1):110-124. [in Chinese])
张慧文, 马剑英, 孙 伟, 等. 2010.不同海拔天山云杉叶功能性状及其与土壤因子的关系. 生态学报, 30(21):5747-5758.
(Zhang H W, Ma J Y, Sun W,etal. 2010. Altitudinal variation in functional traits ofPiceaschrenkianavar.tianschanicaand their relationship to soil factors in Tianshan Mountains,northwest China. Acta Ecologica Sinica, 30(21):5747-5758. [in Chinese])
朱媛君, 杨 劼, 万俊华, 等. 2015.毛乌素沙地丘间低地主要植物叶片性状及其相互关系. 中国沙漠, 35(6):1496-1504.
(Zhu Y J, Yang J, Wan J H,etal. 2015. Leaf traits and their interrelationships of main plant species in inter-dune lowland in the Mu Us Sandy Land.Journal of Desert Research, 35(6):1496-1504. [in Chinese])
张万儒, 杨光滢,屠星南,等.1999. 森林土壤分析方法. 北京: 中国标准出版社.
(Zhang W R, Yang G Y, Tu X N,etal. 1999.Soil analysis method of forest.Beijing:Standards Press of China.[in Chinese])
Cornelissen J H C, Lavorel S, Garnier E,etal. 2003. A handbook of protocols for standardized and easy measurement of plant funcitional traits worldwide. Australian Journal of Botany, 51(4): 335-380.
Evans J R. 1989. Photosynthesis is and nitrogen relationships in leaves of C3 plants. Oecologia, 78:9-19.
Fu D G, Duan C Q. 2007. Advances in plant functional traits in plant ecology∥Duan C Q. Advances in ecological sciences. Beijing: Higher Education Press, 97-121.
Han W, Fang J, Guo D,etal. 2005.Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist,168(2):377-385.
Liu C, Wang X P, Wu X,etal. 2013. Relative effects of phylogeny, biological characters and environments on leaf traits in shrub biomes across central Inner Mongolia, China. Journal of Plant Ecology, 6(3): 220 -231.
Qi J, Ma K M, Zhang Y X. 2008.Comparisons on leaf traits ofQuercusliaotungensisKoidz. on different slope positions in Dongling Moutain of Beijing. Acta Ecologica Sinica, 28(1):122-128.
Reich P B, Uhl C, Walters M B,etal. 1991.Leaf life span as a determinant of leaf structure and function among 23 Amazonian tree species. Oecologia, 86(1):16-24.
Roche P D, Natalia Díaz-Burlinson N, Gachet S. 2004.Congruency analysis of species ranking based on leaf traits: which traits are the more reliable.Plant Ecology,174(1):37-48.
Wilson P J, Thompson K, Hodgson J G.1999. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.New Phytologist, 143(1):155-162.
Wright I J, Reich P B, Westoby M. 2001. Strategy shifts in leaf physiology, structure and nutrient content between species of high-and low-rainfall and high-and low-nutrient habitats. Functional Ecology, 15(4):423-434.
Wright I J, Reich P B, Cornelissen J H C,etal.2005.Modulation of leaf economic traits and trait relationships by climate. Global Ecology and Biogeography, 14(5): 411-421.
Wright I J, Reich P B, Westoby M,etal.2004. The worldwide leaf economics spectrum.Nature, 428(6985): 821-827.
Xia C, Yu D, Wang Z,etal. 2014.Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China. Ecological Engineering, 70(5):406-413.
(责任编辑 王艳娜)
AdaptiveStrategiesofLeafFunctionalTraitsofMosoBambooduringItsExpansiontoChinesefirforests
Liu Guanglu Fan ShaohuiTang XiaoluLiu Xizhen
(KeyLaboratoryforBambooandRattan,InternationalCenterforBambooandRattanBeijing100102)
S718.54
A
1001-7488(2017)08-0017-09
10.11707/j.1001-7488.20170803
2016-05-05;
2016-06-15。
林业公益性行业科研专项“毛竹林扩展模式及其调控技术研究”(201404408)。
*范少辉为通讯作者。
