Netrin蛋白家族的进化分析
2017-10-13陈丹丹刘阿克邹央云
陈丹丹,张 朝,刘阿克,邹央云,谷 迅
(复旦大学 生命科学学院,上海 200438)
Netrin蛋白家族的进化分析
陈丹丹,张 朝,刘阿克,邹央云,谷 迅
(复旦大学 生命科学学院,上海 200438)
神经突起生长导向因子(Netrin)蛋白家族是分泌型蛋白质,其通过与不同的受体相互作用,在神经发育过程中起轴突导向和细胞迁移中发挥着双重导向功能.本文分别对涉及到细胞相互作用的Netrin基因家族的起源与进化进行研究.以进化史不同阶段的代表物种作为研究对象,通过使用MEGA的邻接法和似释然法分别对Netrin基因家族构建系统发育树;使用PAML 4.7工具以核苷酸序列作为分析材料进行选择压分析,筛选出发生正选择作用的位点;使用DIVERGE3.0软件对Netrin氨基酸序列进行功能分化分析,筛选出Ⅰ型和Ⅱ型功能分化位点.
Netrin; 基因家族; 功能分化; 选择压力; 细胞黏连
癌症是一个进化过程的产物,突变在细胞内不断积累到一定程度会使细胞转变成恶性肿瘤.转移特性是恶性肿瘤特有的特性之一,这个特性为肿瘤细胞转移到身体其他组织提供条件,使得肿瘤细胞可以扩散到临近的组织或者通过血液、淋巴系统等到达身体其他部位,浸润目的组织,形成新的病灶.细胞通过解除与其他细胞之间的吸附作用,这个从原发病灶解吸附的过程是转移过程的前提条件,这个过程涉及到与细胞吸附有关的蛋白质的分泌,因此对这些基因的进化机制的研究具有一定的意义.
神经突起生长导向因子(Netrin)是一组具有药物导向作用的蛋白质.在结构方面,Netrin的二级结构高度保守,是一种多结构域(domain)蛋白质.1992年Hedgecock第一次提出Netrin和层黏连蛋白(Laminin)的关系,Netrin蛋白的N末端结构域和Laminin亚基的N末端结构域同源[1].Netrin的C末端在各Netrin蛋白质间具有多态性,且含有与细胞外基质或者细胞表面相互作用的物种特异性氨基酸[2].Netrin具有药物导向的作用,生长中的轴突会移向或者远离更高浓度Netrin.在多种人类癌症中发现,Netrin过表达的同时会有某些相关受体的表达下调.Netrin也会导致细胞凋亡,Netrin- 1的存在可以抑制凋亡通路.在人类直肠上皮细胞中观察到绒毛顶部高水平的细胞自然死亡和Netrin- 1浓度梯度降低有关.肿瘤抑制基因和Netrin- 1的表达有关,暗示Netrin可能涉及到p53调控细胞周期的通路.Netrin对细胞死亡的调控影响很大,因此编码Netrin- 1的基因被认为是原癌基因[3].同时研究发现Netrin对神经系统以外的其他组织的表达和发育有重要作用,如肺、胎盘、脉管系统、胰腺、肌肉和乳腺组织.Netrin通过控制位于分化中的器官内的发育中的细胞的迁移和黏附作用来影响组织的形态形成.在乳腺癌的发育过程中,腔细胞分泌的Netrin- 1和帽状细胞表面受体neogenin结合,使腔细胞层和帽状细胞层具有黏附作用,因此除了对神经系统中的导向作用,Netrin- 1也在哺乳动物中作为黏附因子起作用[4- 5].在胰腺发育过程中,上皮导管细胞表达的Netrin- 1定位于基底膜,和细胞外基质的一些元件(胶原蛋白Ⅵ、纤维连接蛋白和整合蛋白)相互作用.而这些细胞外基质元件与上皮细胞的黏附和迁移有关.Netrin- 1在血管形成、维持肿瘤微环境和转和抑制直肠癌细胞等过程中都起到重要作用[6].
目前对Netrin基因家族的系统的功能分化分析仍旧很少,本文尝试对Netrin基因家族的进化先后顺序、进化选择压和蛋白质功能加以分析,以期对Netrin基因家族的深入研究有所裨益.
1 材料和方法
1.1基因序列的获取
通过HGNC网站(http:∥www.genenames.org)获得已经人工校对的人类Netrin基因家族成员的基本信息,这些信息包含基因名字、基因在染色体的位置、Ensembl ID等(表1).
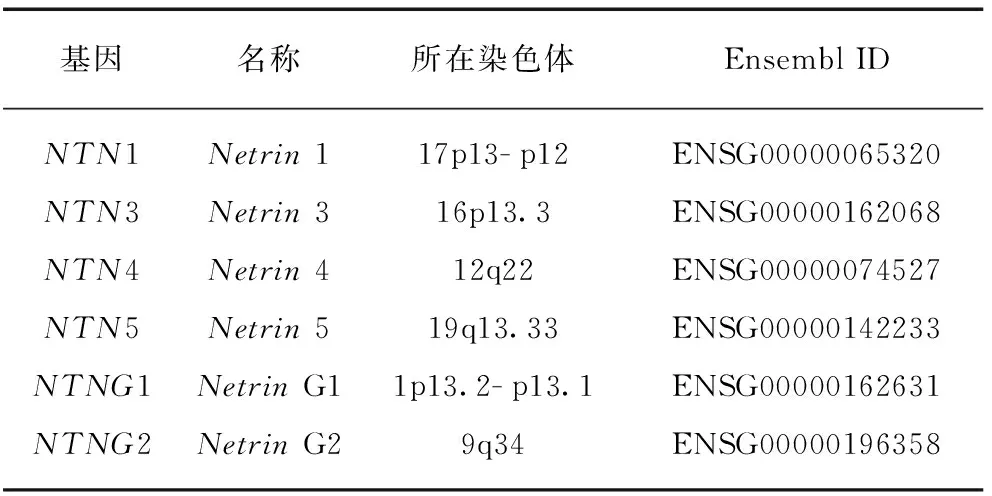
表1 人类Netrin基因家族的基本信息
将人类Netrin基因的Ensembl ID导入Ensembl数据库(http:∥www.ensembl.org/index.html)中的BioMart工具,分别在目标物种数据库中搜索同源基因.将收集到的候选基因作为参考序列,使用BLAST工具在GenBank database中搜索来验证序列真实性.如在某个物种基因组数据库中未找到同源基因,可将人的相应基因作为参考序列,在该物种的基因组数据库中通过BlastN和TblastN工具进行搜索,以进一步核实是否存在该基因.人类与其他物种的同源编码序列(Coding Sequence, CDS)及其蛋白质序列(Protein Sequence)同样通过Ensembl数据库的BioMart工具下载得到[7].用perl脚本作去冗余处理,即针对具有选择性剪切的基因,选取序列最长的选择性剪切形式做序列比对等处理.
1.1.1 氨基酸序列比对
使用MUSCLE(Multiple Sequence Comparison by Log- Expectation)方法对Netrin蛋白的氨基酸序列进行比对.然后通过BioEdit序列比对编辑器(BioEdit Sequence Alignment Editor)对结果进行调整,删除差异大的序列来提高比对的准确度.为保证序列比对结果的准确性,对保留的序列重新进行多序列比对,再通过BioEdit编辑器对比对结果进行修饰.
1.1.2 核苷酸序列比对
将比对后的蛋白质序列及编码序列作为原始数据导入网页PAL2NAL(http:∥www.bork.embl.de/pal2nal/)获得核苷酸序列比对结果.所得比对结果用来计算同义密码子替换率(dS)和非同义密码子替换率(dN).
用WebLogo对氨基酸序列中的功能结构域LamNT区域进行处理,得到序列标识(Sequence Logos),即对氨基酸或核苷酸的进行序列比对结果图形化处理.使用默认参数设置,标识范围为79到598位点.
1.2系统发育分析
本文分别采用邻接法(Neighbor- Joining, NJ)、最小进化法(Minimum Evolution method, ME)以及速度较慢的最大似然法(Maximum Likelihood, ML)3种方法构建Netrin基因家族系统发育树.bootstrap值设为1000,即采用1000次抽样来定量每个进化枝的可信度,一般bootstrap值大于70可认为比较有意义,认为构建的系统发育树比较可靠,高于90%认为显著.
1.3选择压力分析
Netrin直系同源基因和人Netrin基因家族成员进化上的选择压力用PAML4.8分析.PAML(Phylogenetic Analysis by Maximum Likelihood)是一个用极大似然法(Maximum Likelihood, ML)对DNA或蛋白质序列进行分析的程序包.本文主要使用Codeml程序包,本程序包对选择压力的分析模型有位点模型、枝模型和枝位点模型,选择压力使用压力系数ω来表示,ω为非同义替换率dN和同义替换率dS的比值.
位点模型(Site Model)假设密码子或者蛋白质的氨基酸的各位点间有不同的ω值,即不同的dN/dS值[8].Codeml程序包含多个位点模型,通过设置参数NSsites(model=0)实现.如,NSsites=0,1,2,3,7,8时,代表在同一组数据中使用6个模型(3组模型).位点模型采用3对模型检验位点的选择压力,分别为M0(one- ratio model)/M3(discrete)、M1a(nearly neutral)/M2a(positive selection)和M7(β分布模型)/M8(β分布+ω分布模型),通过似然比检验(Likelihood Ratio Tests)来检验最佳模型.其中M0模型假设各个位点具有相同进化速率,称作单一速率模型;M3假设不同位点具有不同的进化速率,参数中的P2、P1和P0分别表示受到正选择、中性选择和纯化选择的位点的比例,相对应的ω值分别为ω0、ω1和ω2;M1a模型(中性选择模型)假设部分位点为保守位点也就是纯化选择位点(ω0<1),位点所占比例为P0,部分位点是中性选择位点(ω1=1),中性位点所占比例为P1=1-P0;M2a模型(正选择模型)中P2=1-P0-P1是正选择位点比例,ω2通过估算得到;M7模型(β分布模型)假设0<ω<1;M8模型(β分布+ω分布模型)假设部分位点是正选择位点(ω>1).M0和M3被用来检验不同位点的不同的ω值,不用来做正选择检测,M1a/M2a和M7/M8两对比较用来检测正选择位点.
枝模型(branch model)假设系统发育树上的不同枝之间有不同的选择压(即不同的ω值),来检测不同种分枝的正选择压[9].枝模型有M0、M1和M2三个模型.其中M0(one- ratio model)模型假设系统发育树各种系之间有单一的选择压,M1(free- ratios model)模型假设系统发育树各种系有各自独立的选择压,M2(two- ratios model)模型假设选定的枝(前景枝,fore- ground branch)和其他枝(背景枝,background branch)相比有不同的选择压,这里需用枝标签标记前景枝.
枝位点模型(Branch- Site Model, BSM)同时考虑不同枝和不同位点的经历的选择压情况,旨在测定特定种系(前景枝)上的正选择位点.最初Yang和Nielsen使用ModelA(model=2,NSsites=2)和ModelB(model=2,NSsites=3)来测定前景枝的正选择位点[10],但是经验证明这两个模型的效果不佳[11],因此将ModelA修改为2个测试[12],即现在的检测正选择压的枝位点模型,是作者推荐使用的一个模型.修改后的前景枝的正选择位点的检测分为两部分,比较零假设(model=2,NSsites=2,fix omega=1,omega=1)和备择假设(model=2,NSsites=2,fix_omega=0)[13].
选择压检验的操作流程如下: 首先使用位点模型的M0/M3、M1/M2和M7/M8初步检测整个基因家族的位点选择压情况,而后按照亚家族分组分别使用位点模型进一步做位点模型的分析,旨在检测每个分组的位点经历的选择压的情况;然后分别使用枝模型M0/M1粗略检测每个分枝接受选择压的情况,最后对每个亚家族进行枝位点模型分析,旨在检测出每个亚家族经历正选择的位点,推测对Netrin亚家族功能形成的重要位点.
1.4功能分歧分析(FunctionDivergenceAnalysis)
采用DIVERGE3.0[14]程序(可在http:∥www.xungulab.com/software.html下载)对Netrin蛋白家族各成员进行功能分化检测.功能分化系数(θ)作为功能分化的衡量标准,θ值在0~1波动,如θ显著大于0,意味存在显著功能分化.Ⅰ型功能分化定义为进化速率发生改变引起的功能分化,Ⅱ型功能分化定义为进化速率未发生改变但氨基酸特性(正负带电性等)发生改变从而引起功能分化.DIVERGE3.0通过后验概率预测发生功能分化的显著程度,通过适当阈值来预测发生功能分化的氨基酸位点.不过θ=0不代表不存在功能分歧位点,可能是功能分歧位点数少被整体掩盖.
分析流程如下: 将多序列比对后的Netrin基因家族的氨基酸序列导入DIVERGE3.0程序中,再将已构建的系统发育树导入DIVERGE3.0中,分别选取NTNG1、NTNG2、NTN5&3、NTN4和NTN1等及其包含的其他序列为独立的5组,对任意的两分组进行比对分析.这里采用DVERGE3.0中Gu99算法进行Ⅰ型和II型功能分化分析.
2 结果和分析
2.1序列收集和氨基酸序列比对
共获得12个物种的45条序列,将Netrin基因在上述物种中的情况汇总(表2)更加直观展示Netrin各基因在不同物种中的存在情况.最终选择39个Netrin基因(表2中Y表示的序列加上斑马鱼的7个基因:NTN- 1a,NTN- 1b,NTN- 2,NTN- 4,NTN- 5,NTN-G1a,NTN-G1b(vav3b))对应的氨基酸序列进行比对,比对结果显示Netrin蛋白含有1947个氨基酸残基(aa),其中结构域LamNT位于79~598aa.这个结构域包含7个保守的位点,分别是186(W)、242(S)、246(G)、249(W)、259(C)、286(C)、342(T)(图1).从比对结果可见Netrin基因家族中功能结构域LamNT的绝对保守位点.同时从Netrin氨基酸全序列的比对结果可见,绝对保守位点仅存在于LamNT结构域中,而对于其他结构域,如EGF_Lam、C354C等,并未显示保守位点,推测可能由于EGF_Lam结构域的多次重复引起.

表2 Netrin基因家族各基因在不同物种中的存在情况
注: Y表示存在相关基因;*倍增.
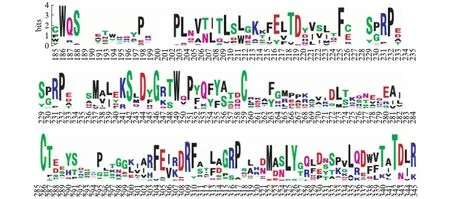
图1 Netrin氨基酸序列比对Fig.1 Multiple alignments of amino acid sequence of Netrin图中字母纵向越高表示该位点越保守.
2.2系统发育分析
构建的系统发育树的结果(图2,见第110页)显示,3种方法构建的系统发育树基本一致,拓扑结构基本吻合且bootstrap值基本一致,基本都在70以上.从拓扑结构看,A节点倍增后形成NTN1和NTN3&5(NTN3&NTN5)两簇,B节点倍增后产生NTN4和NTNG1两簇,其中海鞘(Ciona)的NTNB基因位于NTNG1亚簇的外节点,海鞘的NTNA基因位于NTN4亚簇的外节点,果蝇(fruitfly)的NTN位于NTN1&3&5亚簇的外节点.所以由此推断,NTN1、NTN3和NTN5来自于同个祖先,NTN4和海鞘的NTNA接近,NTNG1&NTNG2海鞘的NTNB接近.从构建的系统发育树的枝长来看,NTN1进化速率最慢,也是最保守,推测NTN1的功能接近稳定,NTNG1s和NTN4的进化速率相对快,推测可能还有很大的进化空间趋向于形成比较稳定的功能.
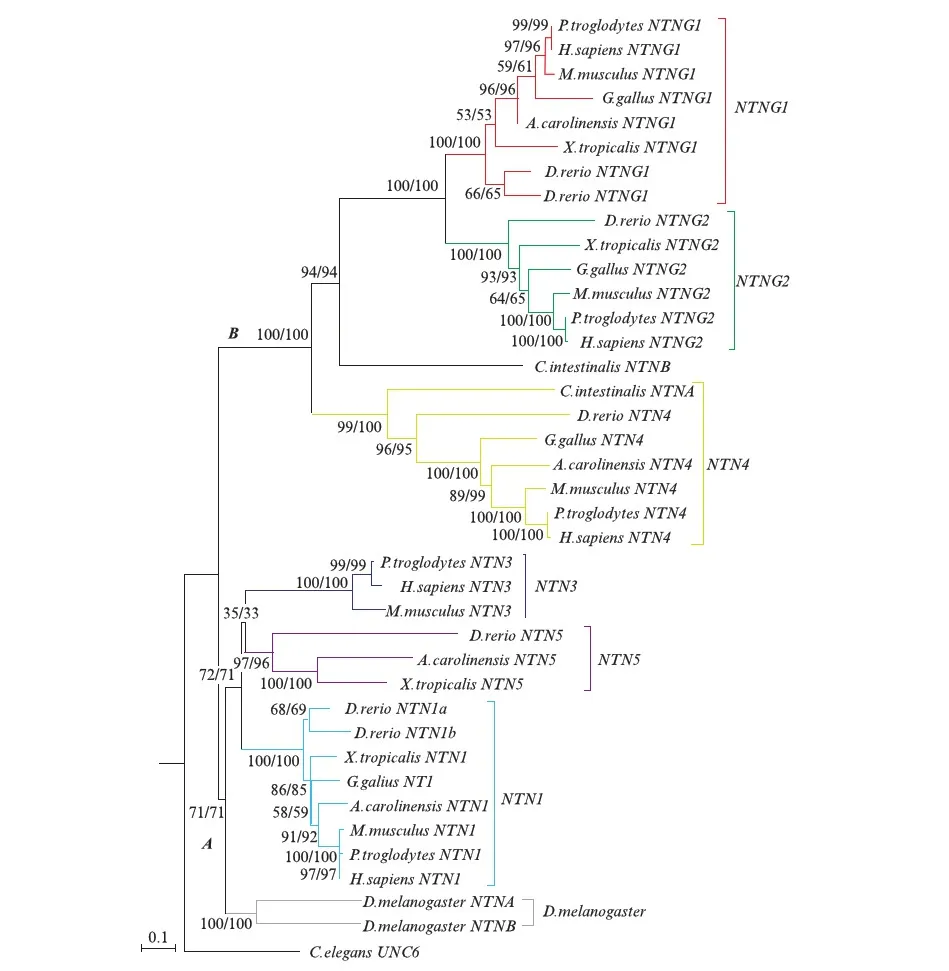
图2 Netrin基因家族系统发育树Fig.2 The phylogenetic tree of Netrin gene family
2.3选择压力分析
首先使用3对位点模型进行选择压分析,结果见表3(第110页).从位点模型的结果中可见,M0vs. M3中P<0.001,存在显著性,可见M3模型优于M0模型,即不同位点有不同的ω值,即经历不同的选择压.随后比对M1a和M2a,M1a是接近中性而M2a是正选择模型,结果不存在显著性,说明从位点模型来看没有明显的正选择现象存在.

表3 位点模型检验结果
这里考虑可能由于不同的基因经历的选择作用差别比较大,湮没了部分基因的选择压力的情况,因此我们使用位点模型将6个Netrin亚家族分别做了分析,结果见表4.
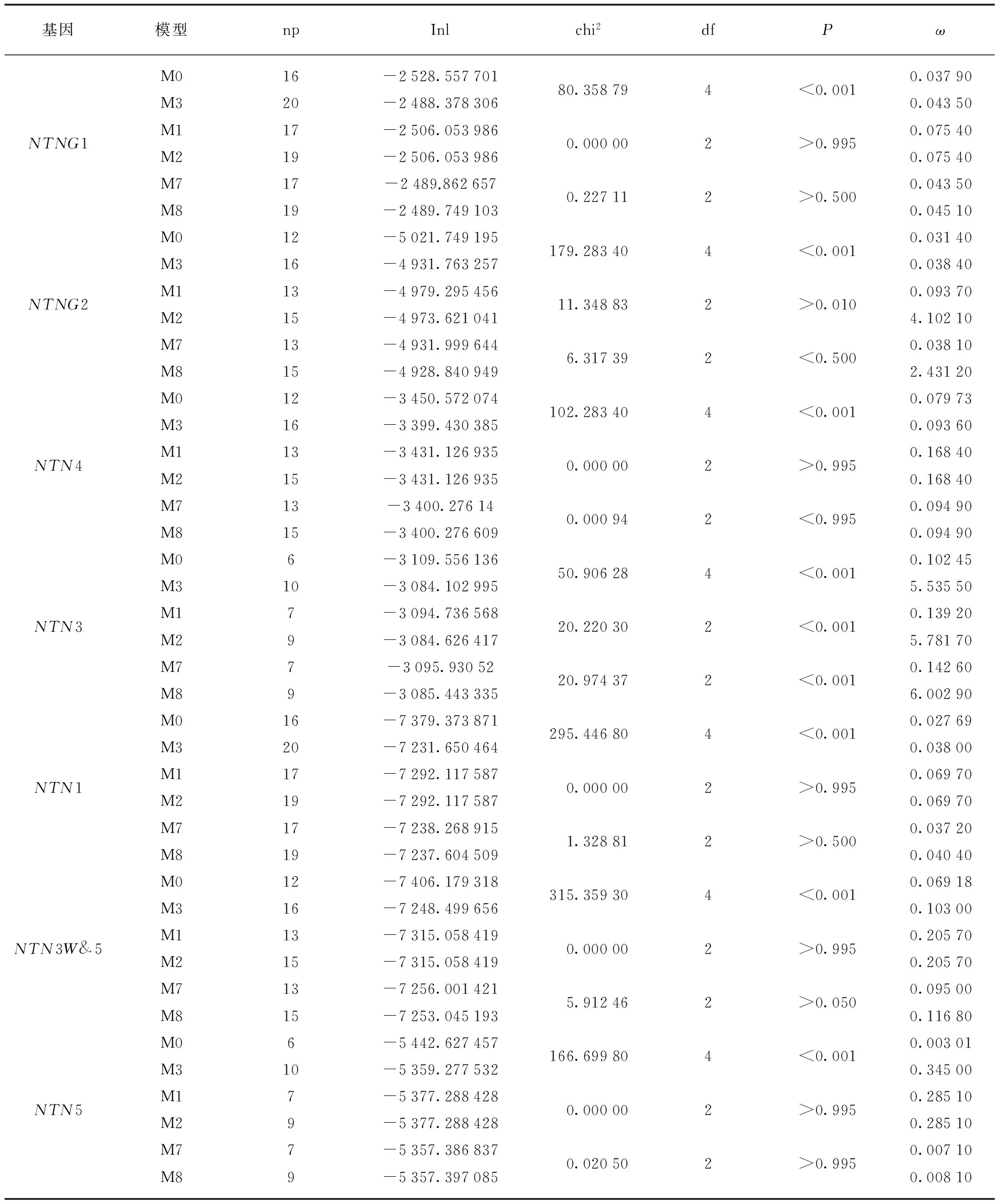
表4 每个Netrin基因位点模型的比对结果以及似然检验值
从结果中显示,在Netrin基因家族的所有基因中M3模型均优于M0模型,表明在所有亚家族中离散模型更合适,即每个基因的每个位点经历的选择压不同,符合大部分功能不同的基因的与预期的结果一致.大部分基因中没有检测出经历正选择的现象,即M1a/M2a和M7/M8均没有显著性.除NTNG2和NTN3,基因的比对结果显示M0/M3、M1a/M2a和M7/M8的检测结果表明NTNG2和NTN3可能均经历了明显的正选择作用.从NTNG2的M2a的参数(表5)可以看出,发生正选择位点的比例为0.00453,大部分位点发生纯化选择作用,6%左右的位点经历中性选择作用.从NTN3的M2a模型的参数(表5,见第112页)可以看出,发生正选择的位点的比例为0.00566,大部分位点(90%)发生纯化选择作用,8.5%的位点发生中性选择作用,正选择ω值为999.00000.枝模型检验结果(表6)显示M0vs. M1有显著性差异,即M1模型优于M0模型,不同的枝有不同的选择压力.

表5 NTNG2和NTN3的M2a模型的参数
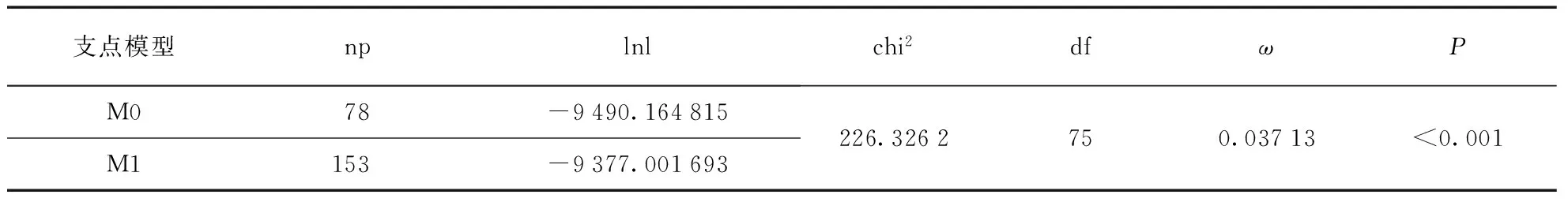
表6 枝模型的参数以及似然比检验结果
检验结果(表7)显示M1模型筛选出经历正选择的位点(ω>1)所在的分枝.
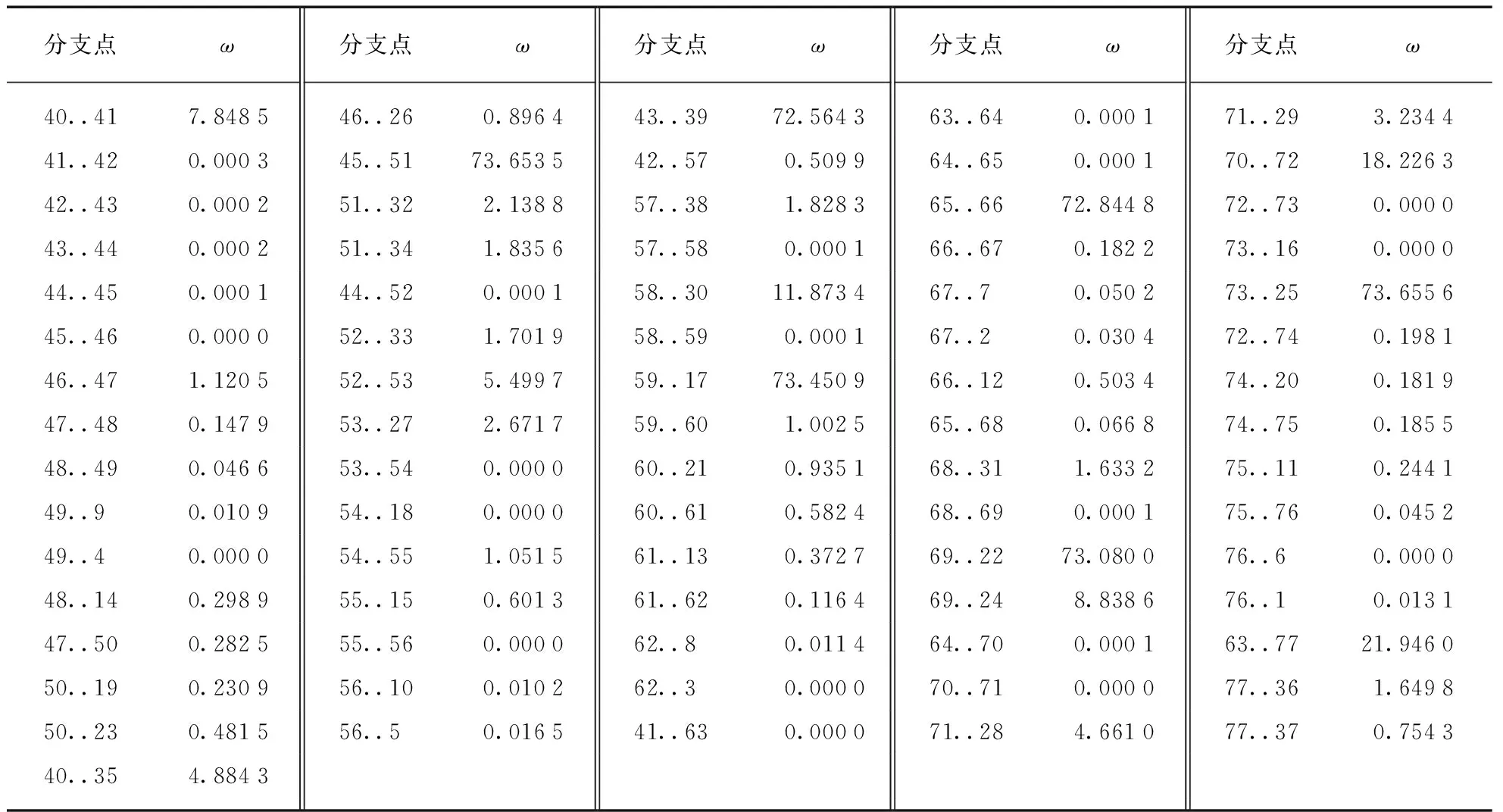
表7 M1枝点模型的结果
注: 灰色阴影标注M1模型筛选出的经历正选择的位点所在枝,即ω>1的位点所在枝的标号及对应的ω的值.
结合系统发育树(图3),发现所涉及到的分枝主要为每个基因亚家族出现前的祖先枝,除果蝇的NTN-a和NTN-b、NTN4的形成,以及海鞘的NTN-b之外.由此推测果蝇可能延续NTN1、NTN3&5的共同祖先的原始功能,而NTN4延续42号节点的祖先基因的功能.同时推测NTNG1、NTNG2、NTN3&5和NTN1在倍增之前可能经历的正选择作用,从而保留下一些非同义突变,增加形成新功能的潜力.除此之外其他的一些枝也存在正选择的现象,已在系统发育树的相应位置标出(图3),这些正选择作用可能对基因功能的形成有一定的潜在作用.
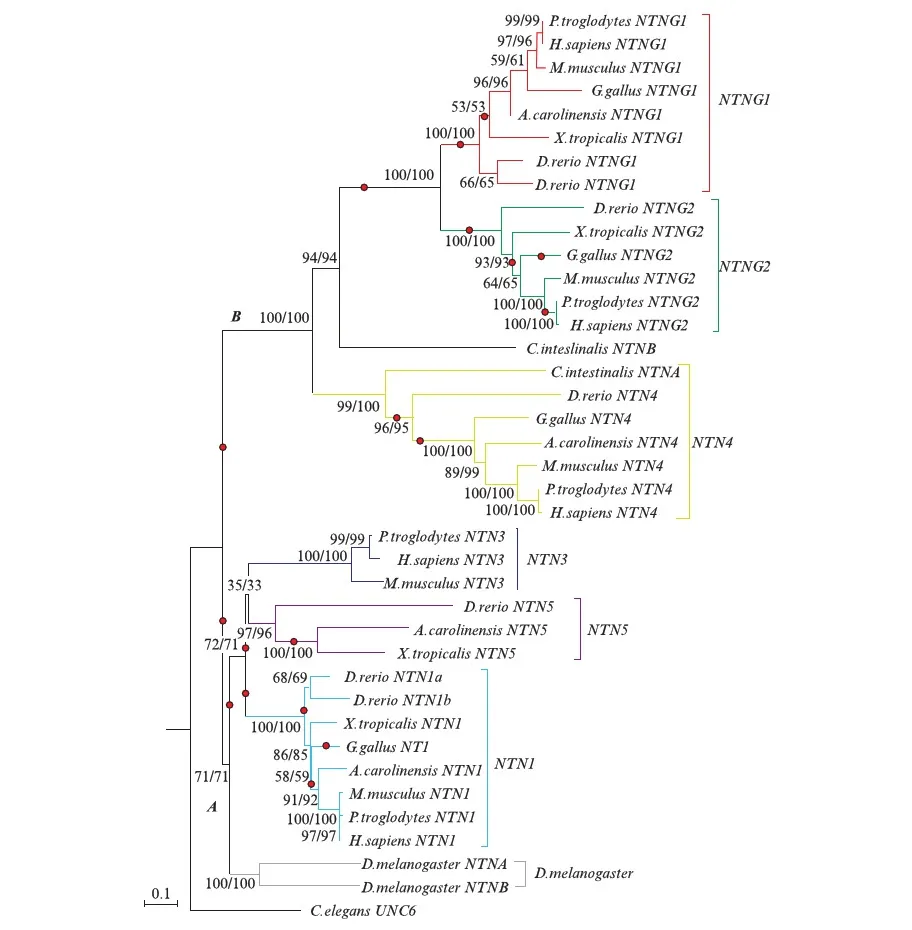
图3 枝模型的M1模型检测出的ω>1的分枝在系统发育树上的位置(红色圆点标记)Fig.3 The position of branches whose ω value is larger than 1 on the phylogenetic tree(red oval dots)
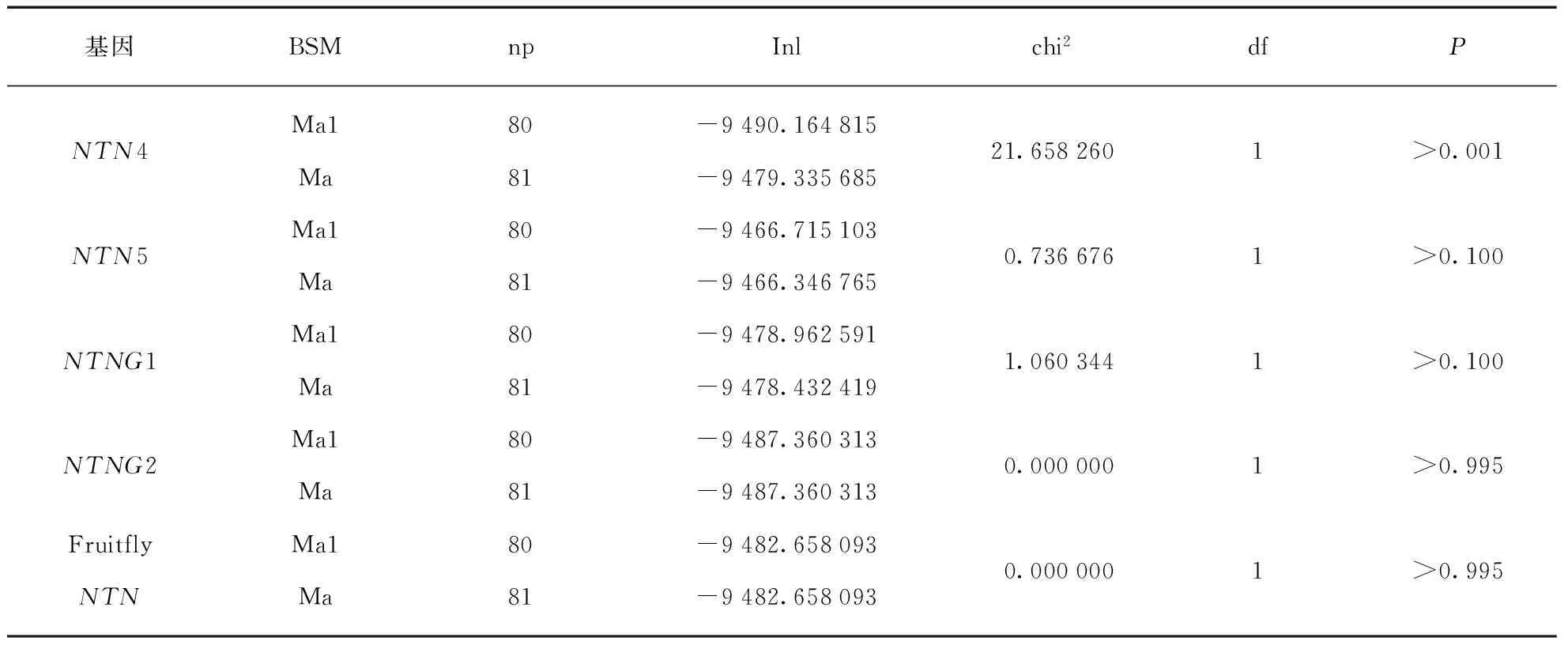
通过枝位点模型Ma(model=2,NSsites=2)和Ma1(model=2,NSsites=2,fix_omega=1,omega=1)分别对Netrin基因家族的不同基因分析,将基因分为7个组(NTN1、NTN3、NTN4、NTN5、NTNG1、NTNG2和fruitfly NTN).结果见表8,使用似然比检验两个模型之间的显著性,得出仅NTN4簇存在显著区别(P<0.001),Ma模型检测发现正选择的位点(13N0.751,115E0.930,73A0.737,116D0.543,87H0.914,121A0.596,88D0.607),后验概率值达到0.9以上的分别有87(H)和115(E),即这两个位点发生正选择,对基因功能的改变或者新功能的形成有潜在的作用.

表8 枝-位点模型检验结果
(续表)
2.4功能分歧分析
结果见表9,Ⅰ型结果中MFEθ显示Netrin亚家族间功能分化程度区别较大,θ值从0.03至1.0不等,也就是各亚组间Ⅰ型功能分化程度波动幅度较大,对应的标准误差SE(Standard Error, SE)为0.15附近.NTNG1/NTNG2和NTN1/NTN3&5两个分组的P值,表示这两个分组内部基因没有显著性,说明NTNG1/NTNG2没有检测到显著的进化速率变化位点;而其余分组的比对结果说明各组间具有显著性,说明存在显著的Ⅰ型功能分化(P<0.01).从Ⅱ型功能分化分析结果(表9)可看出θ值从0.2至0.60,θ的误差在0.1附近浮动,从结果显示NTNG1/NTN4、NTNG1/NTN1、NTNG2/NTN4、NTNG2/NTN1和NTN4/NTN1存在显著Ⅱ型功能分化,即基因组group之间存在进化速率未发生改变而相应的氨基酸特性发生改变的位点.

表9 Ⅰ型和Ⅱ型功能分化分析结果
*MFE: model- free method.E-x表示×10-x.
结合Ⅰ型功能分化的结果发现NTNG1/NTN3&5,NTNG2/NTN3&5及NTN4/NTN3&5均未检测到Ⅱ型功能分化,而NTNG1/NTNG2和NTN1/NTN3&5既无Ⅰ型功能分化也无Ⅱ型功能分化,所以认为这两组的基因功能差别不明显.
由于得到的Ⅰ型功能分化和Ⅱ型功能分化位点的数目比较多,因此我们对位点做了统计结果见表10.分别统计阈值大于0.9和大于0.8的位点数以及所占比例.

表10 Ⅰ型功能分化和Ⅱ型功能分化位点统计分析
1)P>0.9;2)P>0.8.
3 讨 论
通过构建的系统发育树的拓扑结构来展现Netrin基因家族各成员之间在进化上的关系,显示Netrin可分为7个亚家族,分别为果蝇的Netrin和NTN1、NTN3、NTN5、NTN4、NTNG1和NTNG2.其中NTN1、NTN3、NTN5来自于共同祖先,NTNG1、NTNG2和NTN4来自于共同的祖先.从多序列比对结果可以清晰地看出序列中的保守位点多为LamNT结构域的保守位点,可见Netrin基因家族的LamNT结构域相对稳定,猜测为承载Netrin基因原始功能的结构域,而EGF结构域中检测到的保守位点不明显,可能是由于EGF多次重复且重复次数差异大引起.
从对整个基因家族的位点模型的检测结果可以看出,离散模型M1更加适合,即每个位点有不同的选择压力,不过从M1a和M2a的比对结果看,未检测到正选择现象,猜测可能是由于不同的亚家族的进化方向不同等原因湮没了某些重要的正选择位点.根据每个亚家族分类使用位点模型再次检测,从结果可看出NTNG2和NTN3均经历了显著的正选择作用,其他亚家族中的位点模型结果不存在显著性.枝模型M0和M1的检测结果有显著差异,不同的分枝有不同的ω值即不同的选择压,其中M1模型检测到一些受到正选择作用的分枝,这些分枝几乎包括亚家族形成之前的全部祖先基因的进化枝(除NTN4的祖先枝),因此在除NTN4的各亚家族出现之前经历了不同程度的正选择作用.从枝位点模型检测结果看只有在NTN4亚家族检测到显著性,同时检测到几个正选择位点,其中后验值高于0.9的位点有87(H)和115(E),结合枝模型的检验结果,显示NTN4经历的正选择作用主要在NTN4出现之后,枝位点模型检测到的两个正选择位点可能对NTN4的新功能的形成有重要的作用.
Ⅰ型功能分化结果显示NTNG1/NTNG2、NTN1/NTN3&5组外的其他分组均存在明显的Ⅰ型功能分化;Ⅱ型功能分化结果显示,除不存在Ⅰ型共能分化的组,NTNG1/NTN3&5、NTNG2/NTN3&5和NTN4/NTN3&5不存在显著的Ⅱ型功能分化,可见决定NTN1/NTN3&5、NTNG2/NTN3&5和NTN4/NTN3&5的功能分化的主要是Ⅰ型的功能分化位点,即主要由一个亚家族中保守而在另一个亚家族中不保守的位点决定.基因倍增为新功能的形成提供条件,一个基因的序列保持相对稳定,另一个倍增基因的序列发生改变,从而功能分化便会产生.
需要提到的是,2000年的一篇文章中[15]发现鸡种存在Netrin- 2基因,当时的数据量不多,所以猜测在哺乳动物中应该也有Netrin- 2,不过在ENSEMBL数据库中并未找到Netrin- 2基因的数据,因此推测Netrin- 2可能在哺乳动物中丢失,或从未出现,和大多数的发育相关的基因家族相似,Netrin基因家族的倍增发生在脊椎动物进化的早期.研究发现两栖动物Netrin的表达和人Netrin- 1基因很接近,但是与Netrin- 2和Netrin- 3的表达模式差别很大,猜测Netrin- 1基因可能保持了祖先在发育方面的作用,而Netrin- 2和Netrin- 3则已经进化出了新的功能[16].
[1] ISHII N, WADSWORTH W G, STERN B D,etal. UNC- 6, a laminin- related protein, guides cell and pioneer axon migrations inC.elegans[J].Neuron, 1992,9(5): 873- 881.
[2] CHILTON J K. Molecular mechanisms of axon guidance[J].DevBiol, 2006,292(1): 13- 24.
[3] ARAKAWA H. Netrin- 1 and its receptors in tumorigenesis[J].NatRevCancer, 2004,4(12): 978- 987.
[4] LAI W S K, CORREIA J P, KENNEDY T E. Netrins: Versatile extracellular cues with diverse functions[J].Development, 2011,138(11): 2153- 2169.
[5] SRINIVASAN K, STRICKLAND P, VALDES A,etal. Netrin- 1/neogenin interaction stabilizes multipotent progenitor cap cells during mammary gland morphogenesis[J].DevCell, 2003,4(3): 371- 382.
[6] KO S Y, BLATCH G L, DASS C R. Netrin- 1 as a potential target for metastatic cancer: Focus on colorectal cancer[J].CancerMetastasisRev, 2014,33(1): 101- 113.
[7] KINSELLA R J, KAHARI A, HAIDER S,etal. Ensembl BioMarts: A hub for data retrieval across taxonomic space[J].Database(Oxford), 2011,2011: r30.
[8] NIELSEN R, YANG Z. Likelihood models for detecting positively selected amino acid sites and applications to the HIV- 1 envelope gene[J].Genetics, 1998,148(3): 929- 936.
[9] YANG Z, NIELSEN R, HASEGAWA M. Models of amino acid substitution and applications to mitochondrial protein evolution[J].MolBiolEvol, 1998,15(12): 1600- 1611.
[10] YANG Z, NIELSEN R. Codon- substitution models for detecting molecular adaptation at individual sites along specific lineages[J].MolBiolEvol, 2002,19(6): 908- 917.
[11] ZHANG J. Frequent false detection of positive selection by the likelihood method with branch- site models[J].MolBiolEvol, 2004,21(7): 1332- 1339.
[12] ZHANG J, NIELSEN R, YANG Z. Evaluation of an improved branch- site likelihood method for detecting positive selection at the molecular level[J].MolBiolEvol, 2005,22(12): 2472- 2479.
[13] YANG Z. PAML 4: Phylogenetic analysis by maximum likelihood[J].MolBiolEvol, 2007,24(8): 1586- 1591.
[14] GU X, ZOU Y, SU Z,etal. An update of DIVERGE software for functional divergence analysis of protein family[J].MolBiolEvol, 2013,30(7): 1713- 1719.
[15] KOCH M, MURRELL J R, HUNTER D D,etal. A novel member of the Netrin family, beta- Netrin, shares homology with the beta chain of laminin: Identification, expression, and functional characterization[J].JCellBiol, 2000,151(2): 221- 234.
[16] SHIMELD S. An amphioxusNetringene is expressed in midline structures during embryonic and larval development[J].DevGenesEvol, 2000,210(7): 337- 344.
Abstract: Netrins are a class of secretory proteins involved in axon guidance. Netrins are chemotropic; a growing axon will either move towards or away from a higher concentration of Netrin, which involved in different acceptors. This paper studies the evolutionary history ofNetringene family with the sequences of representative species. Phylogenetic trees were constructed by using Neighbor- Joining and Maximum Likelihood methods. The analysis of selection pressure was performed by PAML4.7 tool for detecting positive selected sites. Except these, the function diversity of Netrins was also analysized by using DIVERGE3.0 software, the functional divergence sites of Type Ⅰ and Type Ⅱ in Netrin were screened out.
Keywords:Netrin; gene family; functional divergence; evolutionary selection; selective pressure
TheEvolutionAnalysisofNetrinFamily
CHEN Dandan, ZHANG Zhao, LIU Ake, ZOU Yangyun, GU Xun
(SchoolofLifeSciences,FudanUniversity,Shanghai200438,China)
Q349+.55
A
0427- 7104(2017)01- 0106- 11
2016- 04- 26
国家自然科学基金(31571355)
陈丹丹(1989—),女,硕士研究生;谷 迅,男,教授,通信联系人,E- mail: xgu@iastate.edu.
