西藏山坡流动沙地物种多样性恢复及影响因素
2017-09-29李海东林乃峰廖承锐
李海东,林乃峰,廖承锐,方 颖
西藏山坡流动沙地物种多样性恢复及影响因素
李海东,林乃峰,廖承锐,方 颖*
(环境保护部南京环境科学研究所,南京210042)
在2008年建立的雅鲁藏布江中游的山坡流动沙地植被恢复试验基地,选取丘间地和迎风坡2个类型,并随山坡海拔增加布设了4条调查样带,调查了160个样方的植被恢复物种组成与生长状况,分析了物种丰富度、α多样性和β多样性随微地形改变的变化特征,以期为西藏沙地植被及物种多样性恢复提供实践依据。结果表明:2011年山坡流动沙地恢复植被共有14个种,隶属9个科,12个属。Shannon-Wiener指数在4个调查样带之间的变化较大,Simpson指数和Pielou均匀度指数的变化较小,α多样性指数总体上表现为随物种丰富度的增大而增加。丘间地的群落相似性大于迎风坡,在群落梯度上丘间地的物种多样性恢复效果较好。2016年山坡流动沙地平均植被盖度达65.6%,最大达94%,物种丰富度显著提升。局部微地形、恢复植物种的选择是影响山坡流动沙地物种多样性恢复的主要因素,建议通过设置沙障改变局部微地形,或采用沿等高线人工脚踩回头撒播的方法等,以增加山坡流动沙地植被恢复的物种多样性。
高寒沙地;植被恢复;物种多样性;地形;西藏高原
截止2014年,西藏自治区有沙化土地面积21.58万km2,仅次于新疆和内蒙古地区,居我国第三位。雅鲁藏布江中部流域是西藏社会经济发展的中心,由于地表沙物质丰富、气候干冷多风、植被稀疏低矮等,是遭受风沙灾害影响最严重的区域[1]。关于西藏高寒风沙化土地植被恢复与重建方面的研究仍然较少[2-4],已有的相关研究涉及沙地植被的区系特征、分类和排序[5-6]、顶极群落和种群格局[7-8]、沙漠化过程中物种多样性变化[9]、物种丰富度分布格局[10]等,这些基础性研究指出了高寒沙地植物固沙的先决条件,以及可选的潜在固沙植物种和配置模式,为西藏高原沙化土地植被及物种多样性恢复奠定了初步的基础。
物种多样性是植物群落演替的重要特征之一[11],物种多样性的增加被作为评价退化生态系统恢复成功与否的关键指标之一[12-13]。自2008年以来,笔者所在的课题组先后开展了西藏高寒沙地植物群落结构、优势种的空间格局和生活史、生境因子观测、植被恢复及其潜力综合评价等方面的工作[4,14-15]。研究表明:我国北方优良沙生植物种的恢复效果优于当地乡土沙生植物种,不同类型沙地(河滩、河岸和山坡流动)之间的植被恢复效果差异较大,较差的植被盖度<10%,较好的>45%[16];前期研究强调植被盖度恢复与重建,尚未涉及深层次的物种多样性恢复,不利于稳定生态系统群落的构建。
山坡流动沙地是形成西藏高寒河谷风沙灾害的二次沙源,开展河谷生态安全屏障的保护与建设工作关键是重建与提升其生态防护功能,包括植被盖度及物种多样性恢复。相对于河滩流动沙地、河岸流动沙地,山坡流动沙地不仅相对海拔高、流动性强、面积大,而且坡度大、水分条件差、微地形条件复杂,是一种特殊的河谷型沙地,其植被恢复与重建难度大,基本上不可能实施人工植树造林。本研究结合2008年以来的人工促进植被恢复试验结果,通过样带调查法,详细调查与分析了西藏山坡流动沙地植被恢复的物种组成和生长状况,以及物种多样性随微地形改变的变化特征,以期为加强青藏高原高寒沙地植被与物种多样性恢复提供实践依据。
1 研究区概况
研究地点位于雅鲁藏布江中游山南市的贡嘎县境内,平均海拔约3600 m,属高原温带季风半干旱气候区,冬春干旱多风,夏季温暖湿润,雨热同季。年平均降水量为300~450 mm,90%以上集中于6—9月,且具暴雨性质。蒸发量约为2688 mm,几乎是降水量的7倍。干旱季长达7~8个月,且大风盛行。热量水平不高,年平均气温6.3~8.7℃,6—7月平均气温14.5~16.6℃,1月平均气温-0.8~4.7℃。每年10月至次年4月有冻土出现,其中除1月和12月外,均为日融冻土。无霜期130~140 d,生长季平均气温仅及喜凉作物适宜生长温度的下限。高寒生境胁迫条件下(季节性干旱、低温以及河岸带丰水期的洪水等)河谷风沙化土地极其发育,土壤粒度组成表现为砂粒含量最多,粘粒和粉粒含量很少;土壤pH值呈中性、碱性和强碱性,土壤有机质和全氮含量均很低,全磷和全钾含量均很高[14,16]。
从2008年开始,在与拉萨贡嘎机场隔江相望、雅鲁藏布江北岸的风沙化土地上,选择河滩、河岸和山坡3种类型的流动沙地,以我国北方优良沙生植物种和西藏乡土沙生植物种为供试植物种,采用人工撒播的方式,连续开展了多年的植被恢复试验;其中2008年和2009年撒播的部分供试植物种(中间锦鸡儿、中国沙棘、变色锦鸡儿、西藏锦鸡儿、西藏沙棘、江孜沙棘)在山坡流动沙地物种多样性调查时未见存活苗[4]。河滩、山坡流动沙地的植被恢复效果较好,2~3年生的新播植被盖度在35%以上[14]。山坡流动沙地的坡向主要为东坡和东南坡,少部分为南坡,植被恢复时仅在四周进行了围栏封育,无任何地面处理和灌溉措施,完全依靠天然条件进行植被恢复。
2 研究方法
2.1 样地调查
2011年7月,通过样带调查法,分别在丘间地和迎风坡两种土地类型布设调查样带,进行了山坡流动沙地植被恢复的群落学调查,其中每个类型做1个重复。具体方法为以5 m×5 m的样方作为基本格子单位,自山坡底部向顶部,随高程增大而不断设置新的调查样方,组成一个宽5 m、由一系列样方组成的调查样带。布设的4条调查样带包含了160个5 m×5 m的调查样方,获取了7738个植株的调查数据(表1)。丘间地的坡度范围为2°~31°,迎风坡的坡度范围为1°~23°,有些丘间地调查样方的坡度大于迎风坡,因为其均是发育在河谷山坡流动沙地上,是由河谷特殊的地形条件造成的。
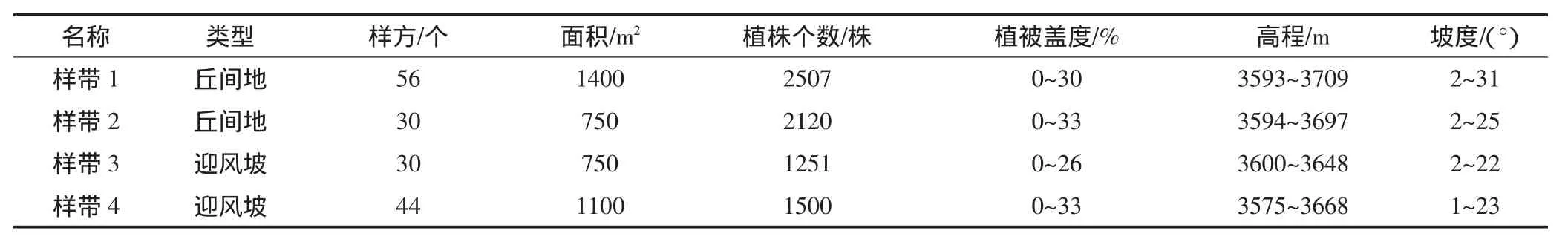
表1 山坡流动沙地恢复植被4条调查样带的基本情况Table 1 Basic situations of the four survey belts of recovered vegetation on valley-slope sand dunes
2016年7月,在上述调查样带中选择10个调查样方进行抽样调查,调查内容主要包括样方的物种组成、植被盖度以及调查植物种的个体数目、冠幅、高度、分盖度等。同时用GPS记录下样方中心点的经纬度、高程,用罗盘测定其坡度、坡向等。
2.2 分析方法
2.2.1 植被生长指标
植株体积是反映其形体大小的重要指标,植株较大的物种能很好地庇护流动沙地植被恢复幼苗的存活和生长。本研究以沙生植物植株长短两个冠幅和地上部分的高度三者的乘积计算植株体积,以其立方根代表植株形体大小[8,17](不考虑量纲),并用植株形体大小和植株高度作为表征植被生长状况的重要指标。
2.2.2 物种多样性指标
(1)物种丰富度指数
丰富度指数S=样方中的物种数。
(2)α多样性
α多样性指某个群落或生境内部的物种多样性,考虑了群落中不同物种的重要值,从而比直接的物种丰富度指标更为准确地反映了植物群落的多样性特征[18-20]。本研究采用3个指标(Shannon-Wiener指数,Simpson指数和Pielou均匀度指数)来测度山坡流动沙地植被恢复群落的α多样性。

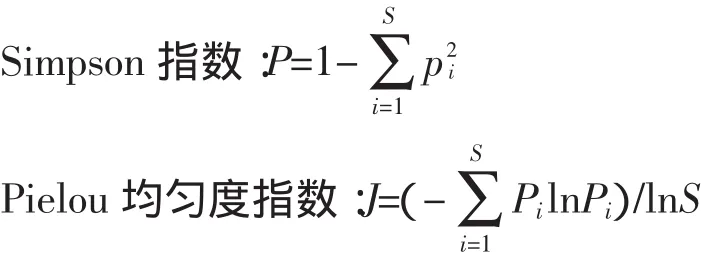
式中:Pi为物种i的个体数占所有物种个体总数的比例;N为所有物种个体总数;S为物种i所在样方的物种总数。
(3)β多样性
β多样性是指沿着某一环境梯度物种替代的程度或速率[12],用以分析不同生境间的梯度变化,可以较直观地反映不同群落间物种组成的差异[21-23]。本研究采用3个指标(Jaccard指数,Cody指数和SÖrenson指数)来测度山坡流动沙地植被恢复的β多样性。

式中:c为相邻两个群落的相同物种数;a和b分别为群落A和群落B的物种数。
3 结果和分析
3.1 植被恢复的物种记载和生长状况
在160个调查样方中共记载了14个植物种,隶属9个科12个属(表2)。个体数较多的为固沙草(Orinus thoroldii)(3424个)、沙米(Agriophyllum squarrosum)(2167个)、花棒(Hedysarum scoparium)(1030个),其他依次为毛瓣棘豆(Oxytropis sericopetala)(291个)、拉萨狗娃花(Heteropappus gouldii)(163个)、杨柴(Hedysarum fruticosum var.mongolicum)(145个)、藏沙蒿(Artemisia wellbyi)(75个)、籽蒿(A.sphaerocephala)(22个)、砂生槐(Sophora moorcroftiana)(22个)、沙拐枣(Calligonum mongolicum)(19个)、荆包紫金标(Ceratostigma ulicinum)(10个)、藏布三芒草(Atistida tsangpoensis)(7个)、香柏(Sabina pingii)(2个)、戟叶醉鱼草(Buddleja hastata)(1个)。在上述7378个调查植物个体中,花棒、杨柴、籽蒿、沙拐枣为引进的我国北方优良沙生植物种,其中花棒、杨柴在山坡流动沙地的适应性较好;其余为西藏当地乡土沙生植物种(藏沙蒿、砂生槐),且大部分为原生沙地植被(如固沙草、藏布三芒草、香柏、荆包紫金标、戟叶醉鱼草)或植被恢复形成的适宜生境条件由试验基地外侵入而新定居的(如拉萨狗娃花、沙米、毛瓣棘豆)物种。
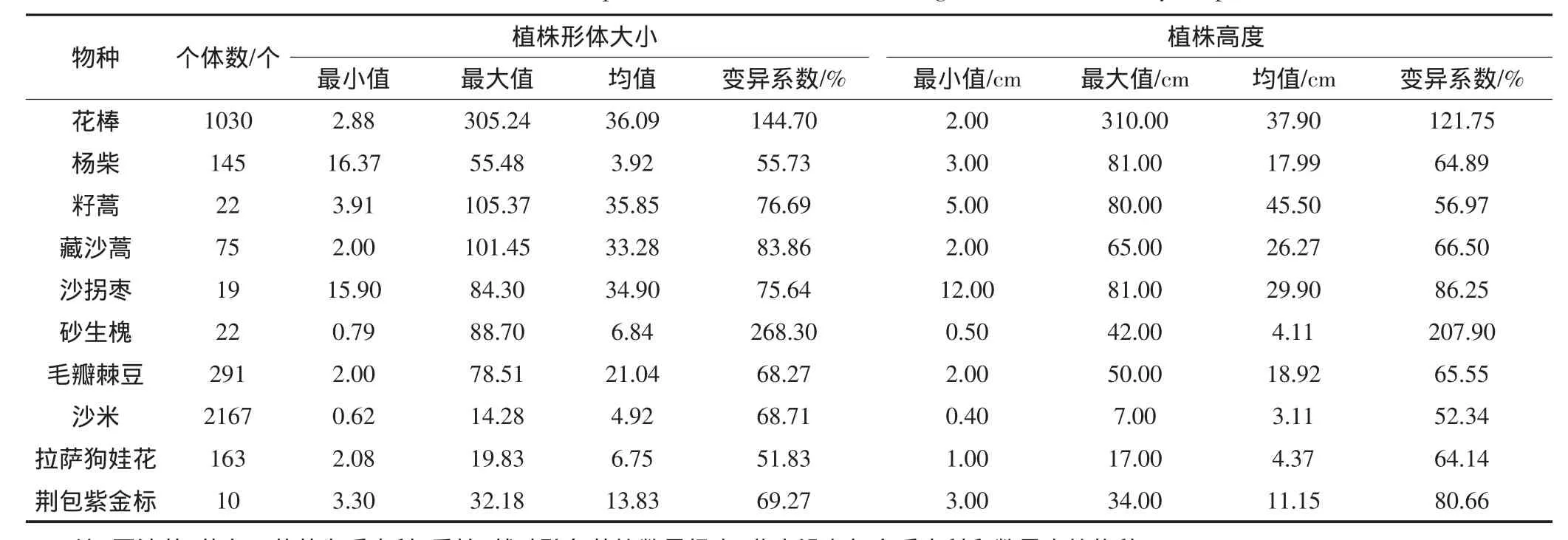
表2 主要调查物种植株形体大小和高度的统计特征Table 2 Statistical values of plant volume cube root and height of the main surveyed species
从植株形体大小的均值来看,花棒最大(36.09),其次为籽蒿(35.85)、沙拐枣(34.90)、藏沙蒿(33.28)、毛瓣棘豆(21.04)和荆包紫金标(13.83),再次为杨柴、砂生槐、沙米和拉萨狗娃花(统计均值<10)。
从植株高度的均值来看,籽蒿最高(45.50 cm),其次为花棒(37.9 cm)、沙拐枣(29.90 cm)、藏沙蒿(26.27 cm)、毛瓣棘豆(18.92 cm)、杨柴(17.99 cm)、荆包紫金标(11.15 cm),再次为砂生槐、沙米和拉萨狗娃花(统计均值<10)。就最大值而言,花棒的最高(310 cm),并远大于杨柴(81 cm)、沙拐枣(81 cm)、籽蒿(80 cm)、藏沙蒿(65 cm)、毛瓣棘豆(50 cm)、砂生槐(42 cm)等。
可以看出,山坡流动沙地植被恢复适应较好的植物种有花棒、杨柴、籽蒿和沙拐枣等,大都是我国北方引进的优良沙生植物种。乡土植物固沙草和沙米的个体数最多,能在植被恢复初期起到阻沙作用,有利于其他物种的定居,可作为流动沙地自然恢复的先锋植物。
3.2 物种丰富度和α多样性的空间差异
由图1可见,4条调查样带的物种丰富度(S)均较低(<7)。样带1、样带2的物种丰富度(平均值分别为2.53和4.3)总体高于样带3、样带4(平均值分别为2.0和2.52),这与调查样带所在的山坡流动沙地微地形有关,丘间地的生境条件较好,有利于新播植物种子的出苗和定居,物种丰富度和植被盖度优于迎风坡。物种丰富度较高的样方主要由于具有灌木-草本的双层结构,既包含从我国北方引进的优良沙生植物灌丛(花棒、杨柴等),又有西藏乡土沙生草本植物(固沙草、沙米等)。样带3、样带4的物种丰富度分别在样方11至样方24、样方13至样方21有两个明显的低值区,这与所在的样方坡度较大,沙地流动性强,加之风蚀剧烈,新播及原有植物无法存活或物种单一、植被盖度较低有关(图1),调查样方内仅发现有零星生长的花棒或固沙草。
可以看出,Shannon-Wiener指数和Simpson指数均以样带2数值最大、样带1最小,Pielou均匀度指数以样带4数值最大、样带3最小,这说明丘间地所在的样带2包含的植物种类较多而样带1较少,迎风坡所在的样带4包含的植物种类较多而样带3较少。结合物种丰富度和植株个体数(图1和表1)可以发现,Shannon-Wiener指数在4个调查样带之间的变化较大,Simpson指数和Pielou均匀度指数的变化均较小,α多样性指数总体上表现为随物种丰富度的增大而增加的趋势,山坡流动沙地的沙丘部位和类型的变化对α多样性指数的影响较小。
就同一个样带内不同α多样性指数之间的大小而言,除样带3外,样带1、样带2和样带4总体上均表现为Shannon-Wiener指数>Simpson指数>Pielou均匀度指数,且随着海拔增加(样方编号增大),同一个样带内3个α多样性指数的波动性变化趋势相似,反映了基本一致的结果(图1)。
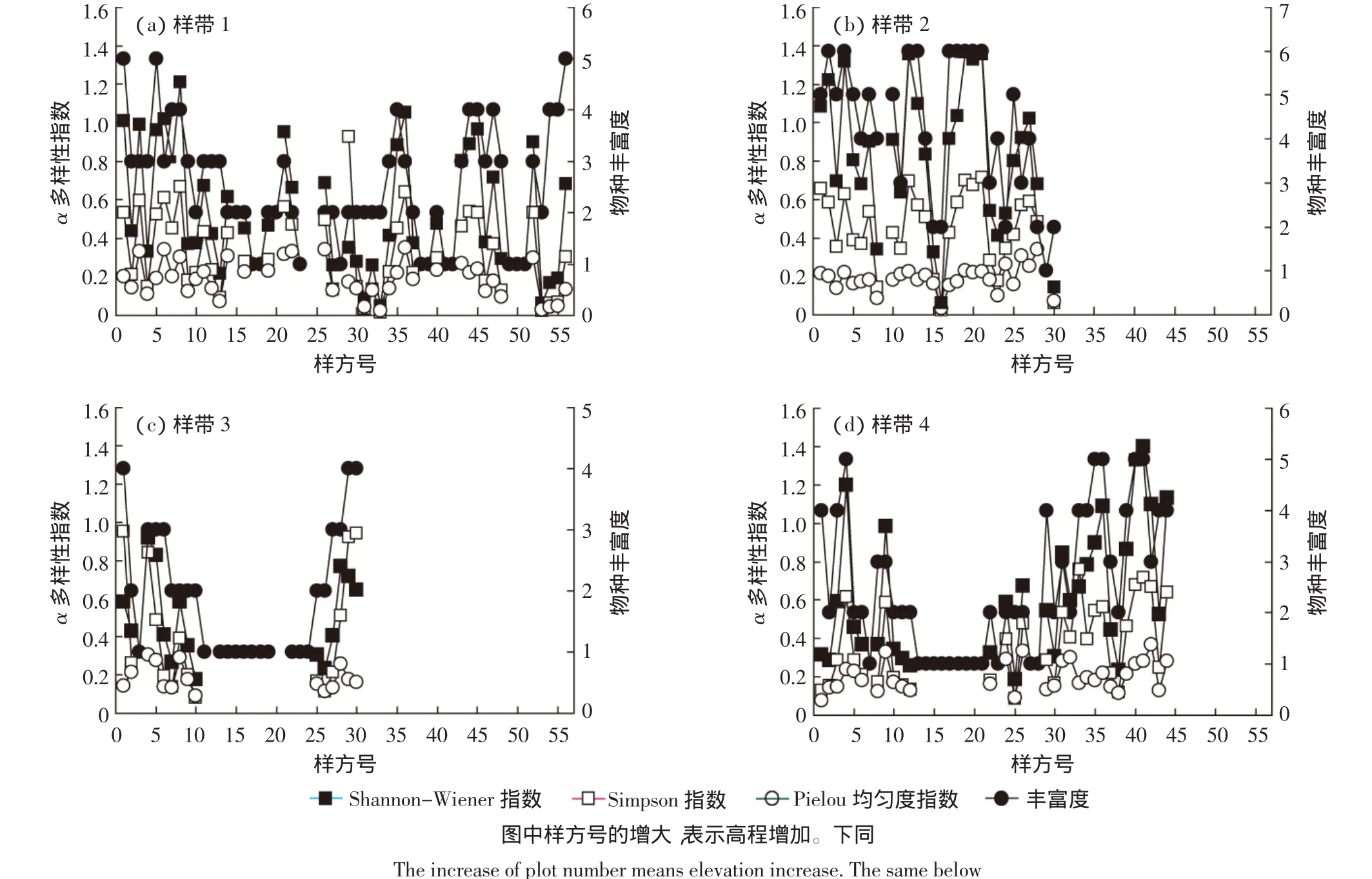
图1 山坡流动沙地恢复植被4条调查样带的α多样性变化Figure 1 Changes of α species diversity in the four survey belts of recovered vegetation on valley-slope sand dunes
3.3 β多样性的空间差异
由图2可见,表示群落相似性的Jaccard指数和SÖrenson指数在不同调查样带之间的变化趋势一致,均表现为迎风坡(样带3、样带4)大于丘间地(样带1、样带2),表示群落相异性的Cody指数的变化趋势与Jaccard指数和SÖrenson指数相反,表现为丘间地(样带1、样带2)大于迎风坡(样带3、样带4)。就同一个样带内不同β多样性指数之间的大小而言,4个调查样带内Jaccard指数和SÖrenson指数随海拔增加(样方编号增大)的变化趋势基本一致,但与Cody指数的变化趋势相反。山坡流动沙地沙丘部位和类型的变化对3个β多样性指数的影响均较大。
Jaccard指数和SÖrenson指数随着海拔增加,表现为无规律的波动趋势。由于山坡流动沙地的坡度大、流动性强,有些调查样方没有新播或原生植物生长,比如样带1在样方22至样方25、样带2在样方8至样方9、样带3在样方11至样方12等,存在β多样性指数的空值区,直接影响了该样带的β多样性。总体而言,丘间地的群落相似性大于迎风坡,这表明在群落梯度上丘间地的物种多样性恢复效果较好。
4 讨论
植被盖度是各种环境因子与植物综合相互作用的结果,能够较好地反映局地生境质量的好坏[24]。Pearson相关分析表明,物种丰富度、除Simpson指数外其他α多样性指数和β多样性指数均与植被盖度呈不显著的正相关(表3)。这说明物种多样性恢复主要受山坡流动沙地微地形的间接影响,表现为植被盖度与高程95%显著性水平的负相关,高程与坡度99%显著性水平的正相关。4个调查样带的最大坡度介于22°~31°,不同样带之间坡度的差异较小。物种丰富度、α多样性和β多样性与坡度之间的相关关系均不显著(表3)。与物种丰富度不同的是,α多样性指数较高的样方(如样带1的样方8和样方36),植被盖度低、坡度较大,没有物种能占据优势;而坡度较小的样方,物种多样性指数却较低(图2)。坡度主要影响水文和落种稳定性,坡度越大,种子越易滑落,种子位移对山坡流动沙地人工撒播治沙不利[25],造成植被盖度随坡度变化较大。由于山坡流动沙地基本无原生植被,植被恢复形成了以花棒、杨柴等占优势的灌木层,且植株形体较大,导致有些调查样方虽然植被盖度较大,但α多样性较低(图2)。
一般认为,物种多样性随海拔的上升而降低,但也有研究认为物种多样性随海拔的升高先增加后减少[26-28]。本研究的调查样带海拔介于3575~3709 m,相对高程较小,仅为134 m。山坡流动沙地α多样性和β多样性指数沿海拔增加的变化规律不明显(图2和图3),区别在于α多样性和β多样性指数在群落水平上的变异程度均低于它们的物种丰富度指数,说明人工促进植被恢复对增加群落内物种个体数影响较大[13]。Pearson相关分析表明,物种丰富度、α多样性指数和除Cody指数外其他β多样性指数均与高程呈不显著的负相关性(表3),这可能是由于α多样性不仅仅反映了物种数量多少(丰富度)的信息,还反映了物种数量的相对多度分布的信息。与自然植被不同,人工促进植被恢复条件下山坡流动沙地的物种多样性沿海拔的变化不明显,由于相对高程较小,随着海拔的上升,在当前尺度下群落的热量条件变化不显著[29],海拔不是影响山坡流动沙地物种多样性恢复的主要因素。

图2 山坡流动沙地恢复植被4条调查样带的β多样性变化Figure 2 Changes of β species diversity in the four survey belts of recovered vegetation on valley-slope sand dunes
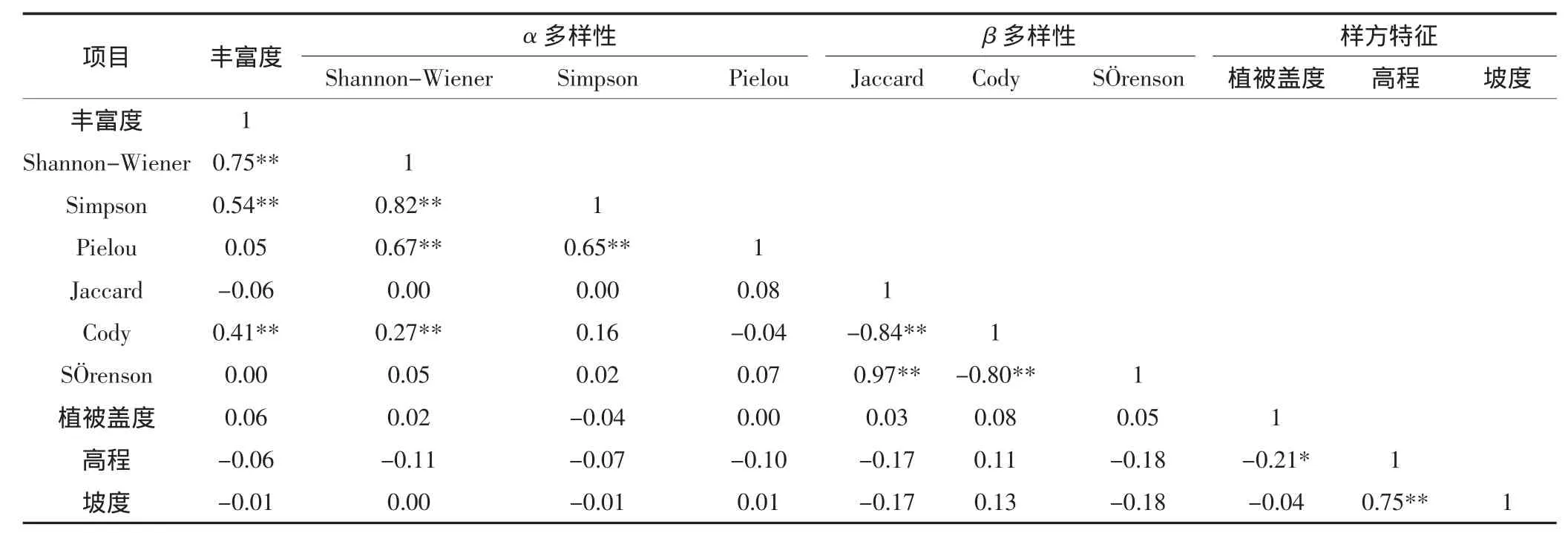
表3 山坡流动沙地物种多样性恢复与生境因子的相关系数Table 3 Pearson coefficients between different species diversity indexes and habitat factors on valley-slope sand dunes
2016年8月,在《西藏航空》报道的“骑着单车拍飞机”,以西藏山坡流动沙地植被恢复试验基地为背景,刻画出一架西藏航空B-6473飞机正在起飞的照片,反映了雅鲁藏布江中游沙化土地植被恢复的现状。2016年7月下旬,笔者对2011年布设的160个调查样方随机抽取10个进行了再次调查(图3和表4),发现山坡流动沙地植被平均盖度已达65.6%,最大可达94%。花棒、杨柴、籽蒿生长较好。花棒高度可达410 cm,冠幅可达560 cm×520 cm,所形成的巨大灌丛,已见小鸟在其中筑巢,野兔在根部打洞;杨柴高度达261 cm,冠幅可达370 cm×410 cm;籽蒿的高度达122 cm,冠幅仅为80 cm×110 cm。物种丰富度介于3~6,平均为4.5,明显高于2011年的平均值(图3a)。这表明2016年的植被盖度和物种多样性较2011年得到显著提高,山坡流动沙地的植被恢复效果得到明显改善。然而,本文关于物种多样性变化研究尚存在不足之处,仅从空间差异性方面分析了物种多样性变化,重在查找影响物种多样性恢复的地形因素,没能从时间变化和样地全面调查的角度,动态分析山坡流动沙地2011—2016年物种多样性变化规律,这是下一步需要重点加强研究之处。
5 结论
(1)西藏山坡流动沙地植被恢复适生植物种主要有花棒、杨柴、籽蒿、沙拐枣、藏沙蒿、固沙草和沙米,丘间地由于坡度较小、生境条件较好,有利于新播植物种子的出苗和定居,物种丰富度和植被盖度优于迎风坡。

图3 西藏山坡流动沙地植被恢复效果Figure 3 Vegetation recovery on the valley-slope sand dunes of Tibet

表4 2016年抽查样方的植被盖度和物种丰富度Table 4 Vegetation coverage and species richness of sampling quadrats surveyed in 2016
(2)物种丰富度的高低取决于山坡流动沙地是否具有灌木-草本的双层结构,综合表现在既有从我国北方引进的优良沙生植物灌丛(花棒、杨柴等),又有西藏乡土沙生草本植物(固沙草、沙米等)。α多样性指数总体上表现为随物种丰富度的增大而增加的趋势,山坡流动沙地的沙丘部位和类型变化对α多样性指数的影响较小。
(3)山坡流动沙地恢复植被的物种多样性变化主要受植物种选择和局部微地形影响,建议通过设置沙障等地面处理措施改变微地形,或采用沿等高线人工脚踩回头撒播的方法,以提高山坡流动沙地植被和物种多样性恢复效果。
[1]Shen W S,Li H D,Sun M,et al.Dynamics of aeolian sandy land in the Yarlung Zangbo River basin of Tibet,China from 1975 to 2008[J].Global and Planetary Change,2012,86/87:37-44.
[2]刘志民.西藏日喀则固沙植物引种的比较研究[J].中国沙漠,1996,16(3):326-330.
LIU Zhi-min.A comparative study of shigatse sand-fixation plant introductioninXigaze,Tibet[J].JournalofDesertResearch,1996,16(3):326-330.
[3]刘志民,高红瑛,蒋德明.西藏日喀则流沙固定的几个问题[J].中国沙漠,2003,23(6):665-669.
LIU Zhi-min,GAO Hong-ying,JIANG De-ming.Several questions of quicksand in Xigaze,Tibet[J].Journal of Desert Research,2003,23(6):665-669.
[4]沈渭寿,李海东,林乃峰,等.雅鲁藏布江高寒河谷流动沙地适生植物种筛选和恢复效果[J].生态学报,2012,32(17):5609-5618.
SHEN Wei-shou,LI Hai-dong,LIN Nai-feng,et al.Screening trial for the suitable plant species growing on sand dunes in the alpine valley and its recovery status in the Yarlung Zangbo River basin of Tibet,China[J].Acta Ecologica Sinica,2012,32(17):5609-5618.
[5]沈渭寿.雅鲁藏布江中部流域沙地植被的分类和排序[J].中国沙漠,1997,17(3):269-273.
SHEN Wei-shou.Classification and sorting of the psammophytic vegetation in the middle Yarlung Zangbo River basin,Tibet[J].Journal of Desert Research,1997,17(3):269-273.
[6]沈渭寿.雅鲁藏布江中游沙地植物区系特征[J].植物分类学报,1996,34(3):276-281.
SHEN Wei-shou.Sand floristic features in middle reaches of the Yarlung Zangbo River[J].Journal of Plant Classification,1996,34(3):276-281.
[7]赵文智,李 森,刘玉璋,等.西藏雅鲁藏布江中游下段沙地植被研究[J].中国沙漠,1994,14(1):68-74.
ZHAO Wen-zhi,LI Sen,LIU Yu-zhang,et al.Study on sand vegetation in bottom of middle reaches of Yaluzangbu River in Tibet[J].Journal of Desert Research,1994,14(1):68-74.
[8]李海东,沈渭寿,方 颖,等.雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析[J].植物生态学报,2011,35(8):834-843.
LI Hai-dong,SHEN Wei-shou,FANG Ying,et al.Point pattern analysis of several psammophyte population in the riparian ecotone in the middle reaches of Yarlung Zangbo River of Tibet,China[J].Chinese Journal of Plant Ecology,2011,35(8):834-843.
[9]魏兴琥,杨 萍,李 森,等.西藏沙漠化典型分布区沙漠化过程中的生物生产力和物种多样性变化[J].中国沙漠,2005,25(5):663-667.
WEI Xing-hu,YANG Ping,LI Sen,et al.Changes of bio-productivity and species diversity in process of desertification in typical desertified land,Tibet[J].Journal of Desert Research,2005,25(5):663-667.
[10]拉 琼,扎西次仁,朱卫东,等.雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释[J].生物多样性,2014,22(3):337-347.
LA Qiong,ZHAXI Ci-ren,ZHU Wei-dong,et al.Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet,China[J].Biodiversity Science,2014,22(3):337-347.
[11]Van der Putten W H,Mortimer S R,Headland K,et al.Plant species diversity as a driver of early succession in abandoned fields:A multi site approach[J].Oecologia,2000,124(1):91-99.
[12]彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1996:84-93.
PENG Shao-lin.South Asian tropical forest community dynamics[M].Beijing:Science Press,1996:84-93.
[13]李全发,刘文耀,沈有信,等.南涧干热退化山地不同恢复群落土壤种子库储量及其分布[J].北京林业大学学报,2005,27(5):26-31.
LI Quan-fa,LIU Wen-yao,SHEN You-xin,et al.Change of plant diversity of different rehabilitated communities on degraded mountains areaofdry-hotvalleyinNanjian[J].JournalofBeijingForestry University,2005,27(5):26-31.
[14]李海东.雅鲁藏布江流域风沙化土地遥感监测与植被恢复研究[D].南京:南京林业大学,2012.
LI Hai-dong.Remote sensing monitoring and vegetation restoration experiments on aeolian sandy land in the Yarlung Zangbo River basin on the Tibet Plateau[D].Nanjing:Nanjing Forestry University,2012.
[15]杨兆平,沈渭寿,孙 明,等.雅鲁藏布江中游河谷风沙化土地砂生槐群落结构特征[J].应用生态学报,2011,22(5):1121-1126.
YANG Zhao-ping,SHEN Wei-shou,SUN Ming,et al.Structural characteristics of Sophora moorcroftiana community on wind-sandy land in middle reaches of Yaluzangbu River[J].Chinese Journal of Applied E-cology,2011,22(5):1121-1126.
[16]李海东,秦卫华,张 涛,等.西藏高寒河谷流动沙地植被恢复潜力综合评价模型[J].农业工程学报,2015,31(16):220-228.
LI Hai-dong,QIN Wei-hua,ZHANG Tao,et al.Comprehensive evaluation model for revegetation potentiality on moving sandy land in alpine valley of Tibet,China[J].Transactions of the Chinese Society of Agricultural Engineering,2015,31(16):220-228.
[17]杨洪晓,卢 琦,吴 波,等.高寒沙区植被人工修复与种子植物物种多样性的变化[J].林业科学,2004,40(5):45-49.
YANG Hong-xiao,LU Qi,WU Bo,et al.Artificial restoration in alpine sandy plantation and the change of the seed plant species diversity[J].Forestry Science,2004,40(5):45-49.
[18]Magurran A E.Ecological diversity and its measurement[M].Princeton:Princeton University Press,1988.
[19]马克平,刘灿然,刘玉明.生物群落多样性的测度方法Ⅱβ多样性的测度方法[J].生物多样性,1995,3(1):38-43.
MA Ke-ping,LIU Can-ran,LIU Yu-ming.Biological diversity measure methodⅡβ biological diversity[J].Biodiversity Science,1995,3(1):38-43.
[20]石胜友,杨季冬,王周平,等.缙云山风灾迹地人工混交林生态恢复过程中物种多样性研究[J].生物多样性,2002,10(3):274-279.
SHI Sheng-you,YANG Ji-dong,WANG Zhou-ping,et al.Species diversity in the progression of ecological restoration of artificial mixed forest after wind-damage in Jinyun Mountain,Sichun[J].Biodiversity Science,2002,10(3):274-279.
[21]Whittaker R H,Marks P L.Method of terrestrial productivity[M]//Leith H,Whittaker R H.Primary Productivity of Biosphere.Berlin:Springer-Verlag,1975.
[22]刘增力,郑成洋,方精云.河北小五台山北坡植物物种多样性的垂直梯度变化[J].生物多样性,2004,12(1):137-145.
LIU Zeng-li,ZHENG Cheng-yang,FANG Jing-yun.Changes in plant species diversity along an gradient on Mt.Xiaowutai,Hebei,China[J].Biological Diversity,2004,12(1):137-145.
[23]陈圣宾,欧阳志云,徐卫华,等.Beta多样性研究进展[J].生物多样性,2010,18(4):323-335.
CHEN Sheng-bin,OUYANG Zhi-yun,XU Wei-hua,et al.A review of beta diversity studies[J].Biodiversity Science,2010,18(4):323-335.
[24]方精云,神崎护,王襄平,等.西藏珠峰-卓奥友峰普士拉地区高山稀疏植被的群落特征及小地形的影响[J].生物多样性,2004,12(1):190-199.
FANG Jing-yun,Kanzaki M,WANG Xiang-ping,et al.Community structure of alpine sparse vegetation and effects of micro-topography in Pushila,Everest-Choyu region,Tibet,China[J].Biodiversity Science,2004,12(1):190-199.
[25]沈渭寿,李海东,张 涛,等.西藏高寒风沙化土地飞播可行性分析[J].生态与农村环境学报,2009,25(1):106-111.
SHEN Wei-shou,LI Hai-dong,ZHANG Tao,et al.Feasibility study of air-seeding on alpine desertified land in Tibet,China[J].Journal of E-cology and Rural Environment,2009,25(1):106-111.
[26]Rahbek C.The relationship among area,elevation,and regional species richness in neotropical birds[J].American Naturalist,1997,149:875-902.
[27]王国宏.祁连山北坡中段植物群落多样性的垂直分布格局[J].生物多样性,2002,10(1):7-14.
WANG Guo-hong.Species diversity of plant community along an altitudinal gradients in the middle section of northern slopes of Qilian Mountain,Zhangye,Gansu,China[J].BiodiversityScience,2002,10(1):7-14.
[28]冯建孟,王襄平,徐成东,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110-116.
FENG Jian-meng,WANG Xiang-ping,XU Cheng-dong,et al.Altitudinal patterns of plant species diversity and community structure on Yulong Mountains,Yunnan,China[J].Journal of Mountain Research,2006,24(1):110-116.
[29]唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
TANG Zhi-yao,FANG Jing-yun.A review on the elevational patterns of plant species diversity[J].Biodiversity Science,2004,12(1):20-28.
Restoration of plant species diversity on valley slope sand dunes and the influencing factors in Tibet,China
LI Hai-dong,LIN Nai-feng,LIAO Cheng-rui,FANG Ying*
(Nanjing Institute of Environmental Sciences,Ministry of Environmental Protection,Nanjing 210042,China)
Desertification has been described as one of the most critically important global problems.By 2014,the sandification area of Tibet Autonomous Region,China was 21.58×104km2,and caused immense damages to regional ecological security and local socioeconomic development.In this paper,the plant species composition and vegetation recovery situation were investigated on sand dunes in the alpine valley slope of the middle reaches of Yarlung Zangbo River of Tibet,China,based on field surveys of four belt transects and 160 quadrats established on two types of interdune and windward sides of sand dunes,respectively.Firstly,the changes in species richness,α diversity,and β diversity were analyzed,and then the influences of elevation,slope,and vegetation coverage on species diversity were explored.The purpose of this study was to provide a practical basis for vegetation and biodiversity restoration on sand dunes in the Tibetan Plateau.The results showed that there were 14 species in the 160 quadrats of vegetation recovery of valley slope sand dunes in 2011,belonging to 12 genera in nine families.In terms of α diversity,the change in the Shannon-Wiener index between the four belt transects was relatively large,while the changes in the Simpson index and Pielou evenness index were small.Generally speaking,the α diversity index presented a similar increasing trend with that of species richness.In terms of β diversity,the community similarity of interdunes was greater than that of windward dunes,and from the viewpoint of community gradient,the result of species diversity restoration on interdunes was also better than that ofwindward dunes.The filed survey in 2016 showed that the average vegetation coverage had reached 65.6%,with the highest 94%.The main factors that influenced the restoration of species diversity were microtopography on valley slope sand dunes and screening for suitable plant species.To improve the species diversity of vegetation restoration,several measures could be used,such as setting a sand barrier to alter the microtopography,using an approach which sows seeds into footprints trampled on sand dunes by feet following the contours of the mountain,or increasing the number of plant species.
alpine sandy land;vegetation recovery;plant diversity;topographic influence;Tibetan plateau
Q948
A
1672-2043(2017)09-1762-09
10.11654/jaes.2016-1605
李海东,林乃峰,廖承锐,等.西藏山坡流动沙地物种多样性恢复及影响因素[J].农业环境科学学报,2017,36(9):1762-1770.
LI Hai-dong,LIN Nai-feng,LIAO Cheng-rui,et al.Restoration of plant species diversity on valley slope sand dunes and the influencing factors in Tibet,China[J].Journal of Agro-Environment Science,2017,36(9):1762-1770.
2016-12-14
李海东(1984—),男,博士,副研究员,主要研究方向为生态保护与修复。E-mail:lihd2020@163.com
*通信作者:方 颖E-mail:fangying@nies.org
环境保护部南京环境科学研究所中央级公益性科研院所基本科研业务专项;国家自然科学基金项目(41301611);江苏省自然科学基金项目(BK20141091)
Project supported:The Basic Special Business Fund for Research and Development for the Central Level Scientific Research Institutes,Nanjing Institute of Environmental Sciences,Ministry of Environmental Protection;The National Natural Science Foundation of China(41301611);The Natural Science Foundation of Jiangsu Province(BK20141091)
