臭氧胁迫对不同敏感型水稻茎秆抗倒性状的影响
2017-09-29邵在胜穆海蓉赵轶鹏贾一磊王云霞杨连新王余龙
邵在胜,穆海蓉,赵轶鹏,2,贾一磊,王云霞,杨连新*,王余龙*
臭氧胁迫对不同敏感型水稻茎秆抗倒性状的影响
邵在胜1,穆海蓉1,赵轶鹏1,2,贾一磊1,王云霞3,杨连新1*,王余龙1*
(1.扬州大学,江苏省作物遗传生理国家重点实验室培育点/粮食作物现代产业技术协同创新中心,江苏 扬州225009;2.江苏徐淮地区徐州农业科学研究所,江苏 徐州221121;3.扬州大学环境科学与工程学院,江苏 扬州225009)
为了研究臭氧胁迫对不同敏感类型水稻成熟期茎秆抗倒性状的影响,利用自然光气体熏蒸平台,以23个水稻品种或株系为供试材料,设置室内对照(10 nL·L-1)和臭氧浓度增高(100 nL·L-1)处理,利用组内最小平方和的动态聚类方法,将所有供试材料按地上部成熟期生物量对臭氧胁迫的响应从小到大依次分为A类(-19%)、B类(-39%)和C类(-52%)3个类别。臭氧胁迫环境下A、B和C类水稻倒3节间抗折力降幅差异不显著,均为19%~20%;但倒4节间抗折力分别下降28%、36%和44%,降幅因类型不同而异,与原位测定的植株田间抗折力的变化趋势一致。臭氧胁迫使水稻基部节间的长度、干重、单位长度干重、横截面积、长直径、短直径和茎壁厚均显著或极显著下降,节间干重降幅最大(平均58%),长直径、短直径以及茎壁厚的降幅最小(平均13%~15%);不同节间比较结果显示,臭氧胁迫对这7个参数的影响多表现为第4节间>第3节间,其中臭氧处理与节间互作对节间长度、单位长度干重、横截面积、长直径和短直径的影响均达极显著水平;从不同类型看,臭氧胁迫下上述指标的降幅多表现为C类>B类>A类水稻,其中臭氧处理与水稻类型的互作对节间横截面积、长直径和短直径的影响均达极显著水平。相关分析表明,臭氧胁迫下基部两节间抗折力的响应与对应节间横截面积、长直径、短直径和节间单位长度干重的响应多呈明显的线性正相关。以上结果说明,100 nL·L-1臭氧浓度使不同类型水稻茎秆质量明显变劣,臭氧胁迫导致的茎秆抗折力显著下降主要与节间变细相关,其次与节间单位长度干重下降亦有关联。
水稻;臭氧;倒伏;茎秆抗折力;生长
近年来,随着工业化与城市化的快速发展,化石燃料消耗持续增加、氮肥大量使用以及汽车尾气排放急剧增加,导致近地层臭氧浓度随氮氧化合物及挥发性有机化合物(VOCs)等含量增加而升高[1-3]。联合国政府间气候变化专门委员会(IPCC)预测,2015—2050年近地层臭氧浓度将会继续上升20%~25%[4],本世纪末将上升到80 nL·L-1[5]。作为最重要的近地层大气污染物之一,不断上升的臭氧浓度已经成为全球科学家和公众密切关注的重要问题[6]。
水稻是人类重要的粮食来源,种植面积非常广泛,是全球50%以上人口的主食[7]。近年来,随着水稻生产方式的改革,直播、抛秧等轻型水稻栽培技术的推广及化肥施用量的不断增加,倒伏对水稻生产的负面影响越来越大[8]。同时,近年来极端天气条件进一步加重了水稻倒伏的严重程度,如洪涝灾害、台风等[9-10]。水稻发生倒伏后因相互遮蔽冠层,光合作用降低[11],影响光合产物的积累和向籽粒的运输[12],导致品质变劣,产量降低,严重制约水稻生产力的提高。研究表明,臭氧胁迫使水稻产量下降[13-14],稻米品质总体变劣[15-16],如果臭氧胁迫增加水稻倒伏风险,这种损失将变得更为严重,因此准确评估臭氧浓度升高对水稻茎秆抗倒能力的影响具有重要意义。
臭氧胁迫对水稻影响的报道多集中在生长发育、产量形成[13,17]和稻米品质[18-19]等方面,聚焦茎秆抗倒性状的报道甚少。这可能与研究手段的局限性有关:前期的气室研究试验空间狭小,加上主要利用盆钵培育试验材料,难以对水稻倒伏这一群体现象进行研究[20-21]。前期针对单一品种的大田研究发现,臭氧胁迫使水稻基部节间抗折力下降,这主要与节间充实程度变差有关[20-21],但这种影响是否因不同品种而异,迄今为止未见报道。水稻茎秆性状还包括节间横截面积(长直径和短直径)和茎壁厚,这些指标亦可能是导致节间抗折力或机械强度变化的重要原因[22-23],但臭氧胁迫下水稻这些茎秆性状的变化及其品种间差异未见报道。2010年本课题组在借鉴传统气室优点的基础上新建了自然光气体熏蒸平台,该平台增大了试验空间,采用自然采光特别是土培方式(而非盆栽)培育水稻植株,并实现了对室外温度等要素的动态模拟,使控制区域的微环境更接近于自然条件[24-25],从而为开展倒伏研究提供了条件。本试验依托这一平台,以23个水稻品种或株系为供试材料,根据成熟期地上部生物量对臭氧胁迫的响应分为3类不同敏感类型水稻,研究100 nL·L-1臭氧浓度下不同类型水稻基部节间抗折力响应的差异及其可能原因,为近地层臭氧浓度不断增加的背景下科学评价水稻茎秆抗倒性提供科学参考。
1 材料与方法
1.1 试验平台与处理
试验于2013年在扬州大学农学院(119.42°E,32.39°N)自然光气体熏蒸平台上进行。试验平台的结构和控制详见赵轶鹏等[24]前期报道,简要说明如下:该平台分为上下两层,下层为设备间,上层共有4间相同并独立的气室,每个气室面积为9 m2,培育面积约为4 m2,气室中间为过道,两侧为气体流通的进风口和出风口,采用自然采光特别是土培方式培育植株,避免盆栽方式对根系生长的限制。在结构上采用分布式拓扑结构,通过实时监测由平台附属气象站观测采集到的温度变化,利用温度调控系统实现对外界环境的动态模拟,使气室内的温度与外界环境的差异维持在最小水平。
本试验共设置2个处理,即对照(C-O3,浓度为10 nL·L-1)和高臭氧浓度(E-O3,浓度设定为100 nL·L-1)处理,每处理两个气室。气体先在下层混气箱中混匀再由风机输送至上层试验区内,由于与室外空气无直接接触,受外界环境干扰较小,臭氧浓度本底值较低,即室内对照臭氧浓度为10 nL·L-1左右,高浓度臭氧是以纯氧为气源,由佳环臭氧发生器(QD-001-3A)产生,通过Model 49i臭氧分析仪对臭氧浓度进行即时监测(间隔时间为1 min),并通过气体熏蒸控制系统(S7-200,Siemens)实现对目标气体浓度的设定。臭氧熏气时间设定为每天上午9:00至下午5:00。温度、光照和大气压力动态模拟外界环境。相对湿度在7月22日之前设定为78%,其后根据室外平均湿度改为65%,9月3日至9月10日又调整为50%。水稻返青后,即6月13日开始进行臭氧熏蒸处理,9月10日停止熏气,共90 d。平台运行期间,除因设备故障、雷雨天气以及臭氧分析仪校准等原因暂停布气外(6月17日、7月10日、7月17—18日和7月20—21日,共6 d),其余时间系统均正常运行。室内对照和高浓度臭氧处理熏蒸期间臭氧浓度分别为10.4 nL·L-1和100.1 nL·L-1,整个生长季臭氧浓度的动态变化以及其他环境参数的平均值详见邵在胜等[26]前期报道。
1.2 材料培育与类型划分
以23个水稻品种或株系为供试材料,分别为L2、L9、L12、L17、L26、L35、L36、L48、L54、L56、L71、L81、L82、L92、L95、L105、L147、L152、L154、L156、Nipponbare、SL41和SL46,其中SL41和SL46为Nipponbare和野生稻品种Kasalath杂交得到的两个株系,其余均为SL41和SL46杂交再自交得到的株系[27]。大田旱育秧,5月14日播种,6月5日移栽,9月10日开始收获。密度为27株·m-2,每室三个重复,均随机分布。试验土壤类型为清泥土,土壤理化性质:有机质24.3 g·kg-1,全N 1.5 g·kg-1,碱解N 126.1 mg·kg-1,全P 0.66 g·kg-1,速效P 13.4 mg·kg-1,速效K 35.2 mg·kg-1,电导率0.17 mS·cm-1,pH 7.1。肥料运筹:总施氮量为15 g·m-2,其中6月4日施基肥(占总施氮量的60%),7月20日施穗肥(占总施氮量的40%);磷、钾肥总施用量均为7 g·m-2,均作为基肥一次性施用。水分管理:6月5日—7月15日保持水层(约4 cm),7月16日—7月25日控水搁田,以轻搁为主,7月26日—8月10日保持水层(约3 cm),8月10日以后干湿交替,8月25日后断水。适时进行病虫草害防治,保证水稻正常生长发育。
最小组内平方和动态聚类方法是以组内平方和之和最小为标准的新的动态聚类方法,能有效地调整初始分组中的个体,使其达到最优的分类,并具有良好的稳健性[28]。地上部最终生物量被广泛地用于衡量水稻逆境条件下生长响应的重要指标[29]。本研究采用组内最小平方和的动态聚类分析方法,根据23个水稻品种或株系成熟期地上部生物量对臭氧胁迫的响应大小,从小到大依次分为A、B、C 3个类别。臭氧胁迫使A、B、C类地上部生物量平均分别下降19.2%、38.5%和51.8%,对A类无显著影响,B和C类均达极显著水平[26,30]。因此,本试验中将A类水稻定义为低度敏感类型水稻,包括L147、L12和L92;B类为中度敏感型,包括L136、L17、L48、L26、L82、L81、L35、L56、L54、L156、L71、SL46和L154;C类为高度敏感型,包括L95、L152、SL41、L2、L9、Nipponbare和L105。
1.3 测定内容和方法
生物量测定:收获时将试验植株以穴为单位分割为叶片、茎秆和稻穗三个部分,其中叶片和茎秆部位装于纸袋中105℃杀青30 min,80℃烘干72 h后称重,稻穗部分自然风干1个月后称重,并计算单株地上部总干重。
田间抗折力测定:用植物倒伏仪(型号YYD-1A)原位测定田间单株抗折力,测定时水稻被推45°角(用十字架构造等腰直角三角形确定45°角),被推点位于距地面20 cm处,此时,植株倒伏测试仪自动记录最大的压力值,即水稻单株承受最大推压阻力。根据田间单株抗折力计算田间单茎抗折力,即:田间单茎抗折力=田间单株抗折力/单株分蘖数。
基部节间性状:于成熟期以穴为单位,在每株水稻中随机挑选长势一致的4根单茎,即:每品种每室共12根单茎,用水洗净植株基部并用吸水纸擦干,然后从节部位剪断,测量各节间长度,将基部倒3和倒4节间剥鞘后用游标卡尺测量节间长直径和短直径,然后采用茎秆强度检测仪(型号YYD-1)测定节间的抗折力(测定时慢慢用力往下压,不可用力过猛,当茎鞘变折时仪器上读数即为抗折力,倒3节间测定时支撑点距离是5 cm,倒4节间为3 cm,计算时将抗折力统一转换成支撑点为3 cm),然后沿各节间中间部位剪断用游标卡尺测定节间茎壁厚,将12根单茎的倒3和倒4节间分别合并后装入纸袋中,105℃杀青30 min,80℃烘干72 h后,测定各节间干重。其中节间横截面积(mm2)=长直径(mm)×短直径(mm)×π/4,节间单位长度干重(mg·cm-1)=节间干重(g)/节间长度(cm)×1000。
1.4 统计分析方法
本试验所有数据采用Microsoft Excel 2013软件进行基础统计和作图,应用SPSS 19.0软件进行数据标准化处理和方差分析。处理间的比较采用最小显著差法(LSD),显著水平设P<0.01、P<0.05、P<0.1、P>0.1,分别用**、*、+和ns表示。
2 结果与分析
2.1 臭氧胁迫对不同敏感型水稻田间抗折力的影响
利用田间茎秆倒伏仪测定水稻成熟期田间单株抗折力,结果见图1。臭氧胁迫使所有水稻田间单株抗折力平均下降30.6%,其中A、B和C类平均分别下降13.6%、33.6%和44.6%,B、C两类水稻均达极显著水平(图1a)。根据单株茎蘖数计算田间单茎抗折力,臭氧胁迫使田间单茎抗折力平均下降11.1%,其中对A类水稻没有影响,而使B和C类水稻分别下降17.2%和16.3%,均达极显著水平(图1b)。方差分析表明,臭氧处理与水稻类型间互作对田间单株抗折力的影响达极显著水平,但对田间单茎抗折力无显著影响(表1)。
2.2 臭氧胁迫对不同敏感型水稻单茎基部节间抗折力的影响
臭氧处理对不同类型水稻基部节间抗折力的影响见图2。结果表明,臭氧胁迫使倒3和倒4节间抗折力平均分别下降19.3%和35.9%。从不同类型看,臭氧处理使A、B和C类水稻倒3节间抗折力分别下降19.7%、18.9%和19.2%,使倒4节间分别下降27.6%、36.1%和44.0%,均达极显著水平。综合方差分析表明,臭氧处理、水稻类型、节间这3个主效应对节间抗折力的影响均达极显著水平;从互作效应看,臭氧与节间的互作对节间抗折力的影响达极显著水平,臭氧×类型、臭氧×类型×节间的互作达到或接近0.1显著水平(表1)。
2.3 臭氧胁迫对不同敏感型水稻茎秆基部节间长度的影响
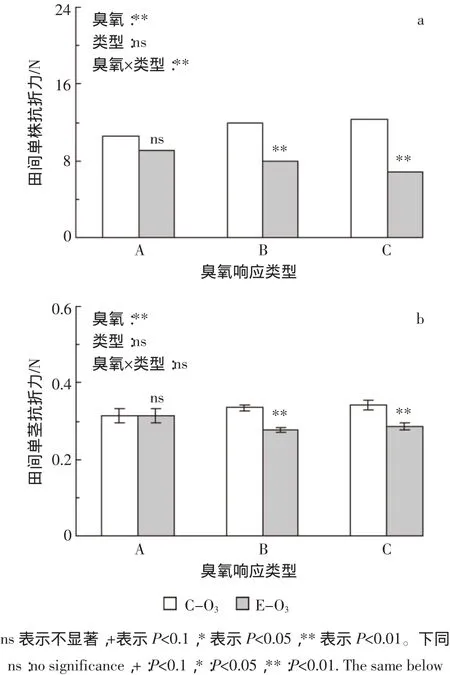
图1 臭氧胁迫对不同类型水稻成熟期田间单株抗折力(a)和单茎抗折力(b)的影响Figure 1 Effects of ozone stress on pushing resistance of the lower part of a single plant(a)and pushing resistance of the lower part of a single stem(b)of the different types of rice at maturity
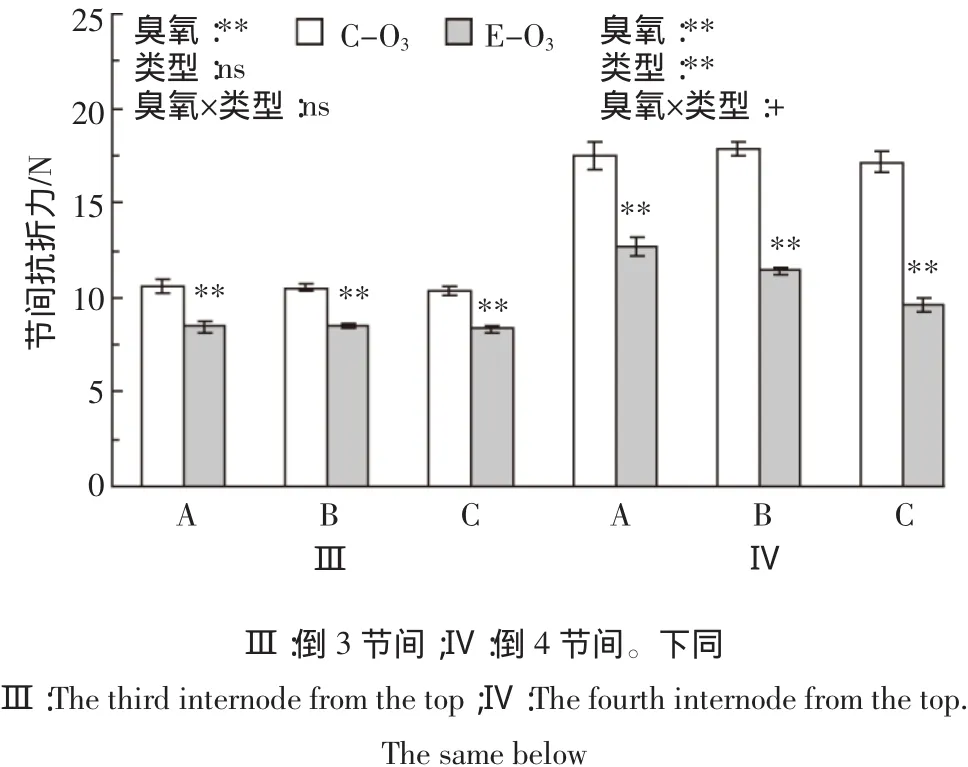
图2 臭氧胁迫对不同类型水稻茎秆基部节间抗折力的影响Figure 2 Effects of ozone stress on breaking resistance of the basal internodes of the different types of rice
臭氧浓度升高对不同类型水稻节间长度的影响见图3。臭氧胁迫使倒3和倒4节间长度平均分别下降33.8%和35.7%。从不同类型看,臭氧胁迫下A、B和C类水稻倒3节间长度分别下降33.4%、31.8%和36.2%,倒4节间分别下降39.2%、38.0%和29.9%,均达极显著水平。方差分析表明,臭氧处理、水稻类型和不同节间对这一参数的影响均达极显著水平,臭氧×节间和臭氧×类型×节间的互作对节间长度的影响分别达极显著和显著水平(表1)。
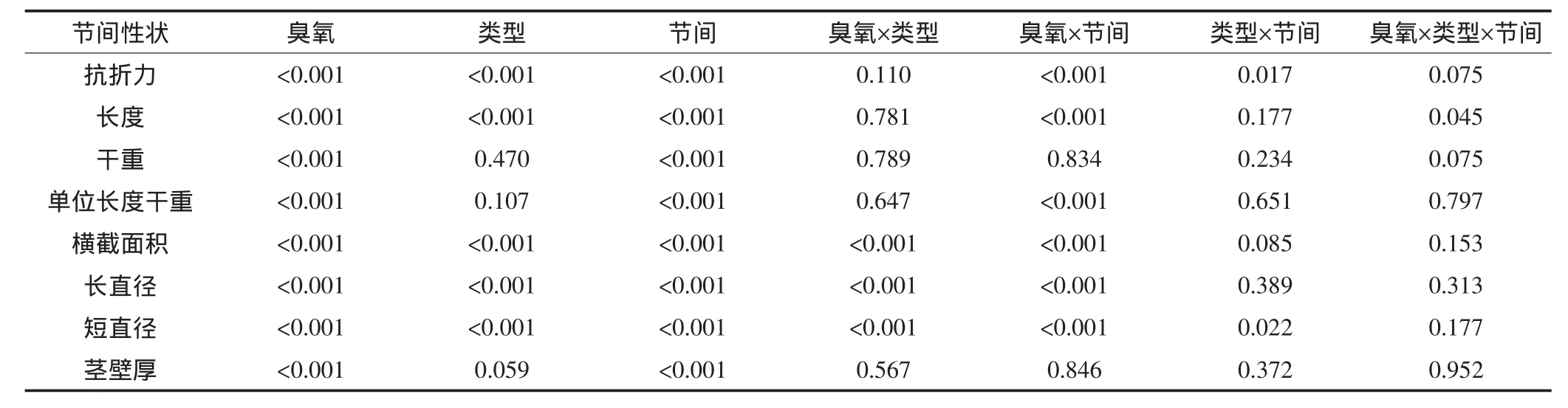
表1 臭氧胁迫对水稻茎秆基部节间性状影响的显著性检验(P值)Table 1 Significance test for the characters of different basal internodes of rice to ozone stress(P values)
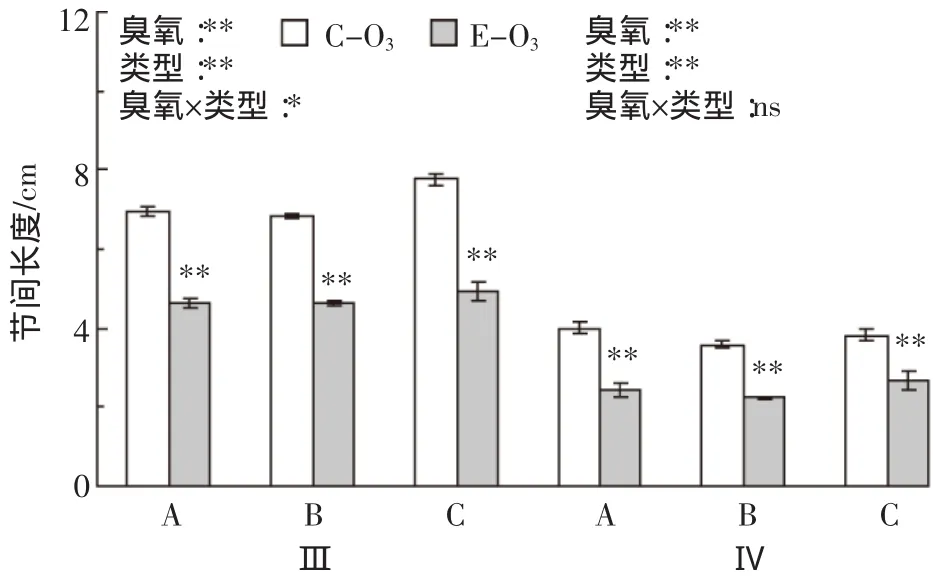
图3 臭氧胁迫对不同类型水稻基部节间长度的影响Figure 3 Effects of ozone stress on the basal internodes length of the different types of rice
2.4 臭氧胁迫对不同敏感型水稻茎秆基部节间干重的影响
水稻基部节间总干重和单位长度干重的结果见图4。臭氧胁迫使水稻倒3和倒4节间干重平均分别下降55.1%和60.8%;从不同类型看,臭氧胁迫下A 、B和C类水稻倒3节间分别下降51.3%、54.5%和59.6%,倒4节间分别下降58.8%、62.9%和60.7%,均达极显著水平(图4a)。相似地,臭氧胁迫使倒3和倒4节间单位长度干重平均分别下降31.1%和38.4%;从不同类型看,臭氧胁迫下A、B和C类水稻倒3节间分别下降28.0%、33.0%和32.3%,倒4节间分别下降38.5%、39.1%和37.7%,均达极显著水平(图4b)。方差分析表明,臭氧处理、节间两主效应对上述两参数的影响均达极显著水平,但从交互作用看只有臭氧×节间对单位长度干重有极显著的互作效应(表1)。
2.5 臭氧胁迫对不同敏感型水稻茎秆基部节间横截面积和茎壁厚度的影响

图4 臭氧胁迫对不同类型水稻基部节间干重(a)和节间单位长度干重(b)的影响Figure 4 Effects of ozone stress on dry weight(a)and dry weight per unit length(b)of the basal internodes of the different types of rice
臭氧浓度升高对不同类型水稻基部节间横截面积的影响见图5a。臭氧胁迫使水稻倒3节间和倒4节间横截面积平均分别下降21.8%和30.4%;从不同类型看,臭氧胁迫下A、B和C类水稻倒3节间分别下降18.9%、20.0%和26.6%,倒4节间分别下降23.3%、31.6%和36.4%,均达极显著水平。方差分析表明,臭氧处理、水稻类型、节间、臭氧×类型以及臭氧×节间对该参数的影响均达极显著水平(表1)。
水稻单茎基部节间横截面积取决于长直径和短直径的大小。从节间长直径看,臭氧胁迫使水稻倒3和倒4节间平均分别下降10.5%和15.3%;从不同类型看,臭氧胁迫下A、B和C类水稻倒3节间长直径分别下降8.6%、9.5%和13.4%,倒4节间分别下降11.4%、15.6%和18.9%,均达极显著水平(图5b)。基部节间短直径对臭氧的响应与长直径趋势一致,但幅度略增(图5c):臭氧胁迫下水稻倒3和倒4节间平均分别下降12.7%和16.1%;从不同类型看,臭氧胁迫下两节间短直径的降幅均表现为C类>B类>A类水稻。与横截面积相同,臭氧处理、水稻类型、节间、臭氧×类型以及臭氧×节间对这两个参数的影响均达极显著水平(表1)。
水稻基部节间茎壁厚的测定结果见图5d。臭氧胁迫使倒3和倒4节间茎壁厚平均分别下降14.0%和13.8%;从不同类型看,臭氧处理使A、B和C类水稻倒3节间分别下降12.9%、15.0%和14.0%,使倒4节间分别下降9.7%、16.8%和15.0%,均达显著或极显著水平。方差分析表明,臭氧处理对节间茎壁厚的影响达极显著水平,两节间茎壁厚亦存在极显著差异,但不同主效应间均无显著的互作效应(表1)。
2.6 水稻基部节间抗折力对臭氧胁迫的响应与茎秆其他性状响应的关系
相关分析表明,臭氧胁迫下倒3节间抗折力的响应与节间横截面积、长直径和短直径的响应呈显著或极显著线性正相关,相关系数分别为0.567、0.442和0.656(表2)。倒4节间抗折力对臭氧胁迫的响应除了与这三个参数的变化密切相关外,还与节间干重和单位长度干重的响应呈显著正相关,相关系数的变幅为0.352~0.531(表2)。以上结果说明,臭氧胁迫下水稻单茎基部节间抗折力的下降主要与节间变细有关,其次亦与节间单位长度干重下降有关。
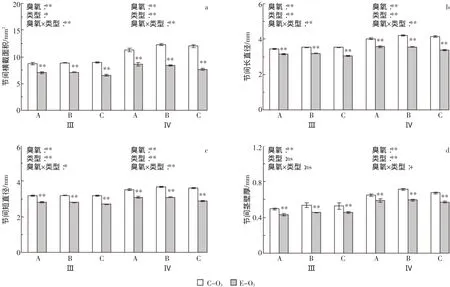
图5 臭氧胁迫对不同类型水稻基部节间横截面积(a)、长直径(b)、短直径(c)和茎壁厚(d)的影响Figure 5 Effects of ozone stress on cross-sectional area(a),maxium stem diameter(b),minium stem diameter(c)and culm wall thickness(d)of the basal internodes of the different types of rice

表2 水稻基部节间抗折力对臭氧胁迫的响应与其他节间相关性状响应的相关分析(n=23)Table 2 Correlation analysis of the ozone-induced changes in breaking resistance of the basal internodes and other internodes relevant characters
3 讨论
为了准确预测地表臭氧浓度升高对未来稻作生产的影响,必须定量研究臭氧胁迫对水稻茎秆抗倒性状的影响,但目前这方面的报道甚少[20-21]。本研究利用空间相对较大的自然光气体熏蒸平台[24],在群体土培条件下,首次对臭氧胁迫下不同敏感类型水稻基部节间的抗折力及其相关形态性状进行了研究。
田间单株抗折力将植株作为一个整体来考虑,综合了单株鲜重、茎秆弹性等影响因素,反映田间植株倒伏需受到的外力大小,较能反映生产实际[8]。臭氧胁迫对水稻原位田间抗折力的影响未见报道。本研究结果表明,与干净空气相比,100 nL·L-1臭氧浓度使供试材料田间单株抗折力大幅下降,降幅因不同水稻类型而异:B和C两类水稻的降幅分别是A类水稻的2倍和3倍(图1a)。这一结果说明臭氧胁迫使水稻整株的机械强度明显下降,且越敏感的水稻降幅越大。相似地,臭氧胁迫亦使水稻田间单茎抗折力显著下降,且B和C两类水稻对臭氧的响应显著大于A类水稻(图1b)。与田间单株抗折力不同,臭氧胁迫对田间单茎抗折力的影响(平均下降11%)明显小于单株抗折力的响应(平均下降31%),且不同水稻类型间差异较小,可能与分蘖数的变化有关。本试验同期试验发现,臭氧胁迫明显抑制水稻分蘖的发生,全生育期A、B和C类水稻平均降幅分别为4%、17%和23%[26]。分蘖的这种差异在一定程度上使不同类型水稻单茎抗折力的响应差异变小。
水稻自身的抗倒伏能力受茎秆特性决定,其中单茎基部节间的抗折力反映茎秆的硬性和韧性,是表示茎秆强度的主要指标,抗折力越大,茎秆抗倒伏的能力就越强[31]。前人研究结果表明[20],臭氧胁迫使两优培九抽穗后35 d倒3和倒4节间抗折力分别下降10%和17%,均达显著水平,前者降幅明显小于后者。本试验对23个供试材料的观察结果表明,臭氧胁迫使水稻成熟期基部倒3和倒4两节间的抗折力均大幅下降,但倒4节间的降幅(-36%)几乎是倒3节间的两倍(-19%),臭氧与节间存在极显著的互作效应(表1)。臭氧胁迫对作物生长的影响存在累积效应[14,30],抗折力响应的这种差异可能与臭氧胁迫时间有关:水稻倒4节间先于倒3节间形成,在臭氧中熏蒸的时间更长,因此造成的累积伤害也更大。本文根据最终生物量对臭氧胁迫的响应,将23个供试材料从小到大分为A、B和C类水稻。结果发现臭氧×类型×节间对水稻茎秆抗折力有互作效应(表1),主要与臭氧处理对倒4节间抗折力的影响因水稻敏感类型而异有关:A、B和C类水稻平均分别下降28%、36%和44%(图2)。倒4节间的这种变化与原位测定的植株田间抗折力的变化趋势一致(图1a)。
水稻茎秆的节间抗折力与节间长度、粗度、茎壁厚、干重、单位长度干重等性状有关[32-34]。水稻基部节间是近似规则的椭圆形结构,为了全面度量水稻基部节间的粗细程度,我们同时测定了这些节间的长直径、短直径、横截面积和茎壁厚,这些参数对臭氧胁迫的响应及其品种间差异未见报道。同时分析这些参数以及基部节间长度、干重和单位长度干重的变化,结果发现,臭氧胁迫使这7个参数均显著或极显著下降,其中与节间重量相关的参数降幅最大,而与茎秆粗细相关的参数如长直径、短直径以及茎壁厚的降幅较小。本研究观察到的茎秆性状对臭氧处理的响应(最大达58%)明显大于前期大田研究[20-21],除了与本试验臭氧处理浓度较高有关外,可能亦与供试材料较为敏感有关。从不同节间看,臭氧胁迫下倒3节间这些性状的响应明显小于倒4节间(茎壁厚除外),进一步说明臭氧胁迫对水稻节间生长亦存在明显的累积伤害。不同类型间比较显示,多数情况下臭氧胁迫对上述指标的影响表现为C类>B类>A类水稻,其中臭氧×类型对节间横截面积、长直径和短直径有极显著的互作效应(表1),后者与田间原位单株抗折力和倒4节间抗折力的变化趋势一致(图1a和图2)。
前人报道臭氧胁迫下水稻基部节间抗折力减弱主要与节间单位长度干重下降有关,但该研究没有测定节间横截面积[20-21]。本研究相关分析表明,水稻基部倒3和倒4节间抗折力对臭氧胁迫的响应与对应节间横截面积、长直径和短直径的响应均存在明显的线性正相关,与节间单位长度干重的响应亦存在一定的正相关关系(表2)。除此之外,臭氧胁迫下田间原位单株抗折力亦与基部节间横截面积、长直径、短直径和充实度密切相关(数据未列出)。可见,臭氧浓度增加使不同敏感类型水稻田间抗折力和基部节间抗折力下降,降幅主要与节间变细密切相关,其次与节间单位长度干重下降亦有一定关联。另外,水稻的抗倒能力除了与茎秆形态性状有关外,亦与茎秆的化学组成以及植株的生理活性有关联[35-36]。因此,臭氧胁迫环境下这些性状的变化及其与茎秆强度的关系需要进一步研究。
4 结论
本文以23个水稻品种或株系为供试材料,研究臭氧浓度增加对不同敏感类型水稻茎秆抗倒性状的影响。与干净空气相比,100 nL·L-1臭氧浓度使所有供试材料的茎秆质量明显变差,这种变化不同程度上与节间位置和水稻的敏感类型有关;相关分析表明,臭氧胁迫导致的茎秆抗折力明显下降主要与基部节间变细有关,与节间单位长度干重下降亦存在一定关联。
[1]Feng Z Z,Hu E Z,Wang X K,et al.Ground-level O3pollution and its impacts on food crops in China:A review[J].Environmental Pollution,2015,199:42-48.
[2]刘 峰,朱永官,王效科.我国地面臭氧污染及其生态环境效应[J].生态环境,2008,17(4):1674-1679.
LIU Feng,ZHU Yong-guan,WANG Xiao-ke.Surface ozone pollution and its eco-environmental impacts in China[J].Ecology and Environment,2008,17(4):1674-1679.
[3]Shao M,Tang X Y,Zhang Y H,et al.City clusters in China:Air and surface water pollution[J].Frontiers in Ecology and the Environment,2006,4(7):353-361.
[4]Intergovernmental Panel on Climate Change(IPCC).Fifth Assessment Report[EB/OL].Geneva:ICPP,2013.http://www.ipcc.ch/report/ar5/index.shtml.
[5]Fiscus E L,Booker F L,Burdey K O.Crop responses to ozone:Uptake,modes of action,carbon assimilation and partitioning[J].Plant,Cell and Environment,2005,28(8):997-1011.
[6]Ashmore M R.Assessing the future global impacts of ozone on vegetation[J].Plant,Cell&Environment,2005,28(8):949-964.
[7]Maclean J L,Dawe D C,Hardy B,et al.Rice almanac:Source book for the most important economic activity on earth[M].3rd edn.Wallingford:CABI Publishing,2002.
[8]陈桂华,邓化冰,张桂莲,等.水稻茎秆性状与抗倒性的关系及配合力分析[J].中国农业科学,2016,49(3):407-417.
CHEN Gui-hua,DENG Hua-bing,ZHANG Gui-lian,et al.The correlation of stem characters and lodging resistance and combining ability analysis in rice[J].Scientia Agricultura Sinica,2016,49(3):407-417.
[9]Ishimaru K,Togawa E,Ookawa T,et al.New target forrice lodging resistance and its effect in a typhoon[J].Planta,2008,227(3):601-609.
[10]Phelan L.Managing climate risk:Extreme weather events and the future of insurance in a climate-changed world[J].Australasian Journal of Environmental Management,2011,18(4):223-232.
[11]Setter T L,Laureles E V,Mazaredo A M.Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis[J].Field Crops Research,1997,49(2/3):95-106.
[12]Hitaka N,Kobayashi H.Studies on the lodging of rice plants(2)Source of decreasing yield due to lodging[J].Japanese Journal of Crop Science,1961,32(4):270-276.
[13]Ainsworth E A.Rice production in a changing climate:A meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration[J].Global Change Biology,2008,14(7):1642-1650.
[14]Rai R,Agrawal M.Impact of tropospheric ozone on crop plants[J].Proceedings of the National Academy of Sciences,India Section B:Biological Sciences,2012,82(2):241-257.
[15]宋琪玲,齐义涛,赵轶鹏,等.自由空气中臭氧浓度升高对“武运粳21”稻米物性及食味品质的影响[J].中国生态农业学报,2013,21(5):566-571.
SONG Qi-ling,QI Yi-tao,ZHAO Yi-peng,et al.Impact of free air ozone concentration enrichment on cooked rice(Wuyunjing 21)texture and palatability[J].Chinese Journal of Eco-Agriculture,2013,21(5):566-571.
[16]Wang Y X,Song Q L,Frei M,et al.Effects of elevated ozone,carbondioxide,and the combination of both on the grain quality of Chinese hybrid rice[J].Environmental Pollution,2014,189(12):9-17.
[17]杨连新,王余龙,石广跃,等.近地层高臭氧浓度对水稻生长发育影响研究进展[J].应用生态学报,2008,19(4):901-910.
YANG Lian-xin,WANG Yu-long,SHI Guang-yue,et al.Responses of rice growth and development to elevated near-surface layer ozone(O3)concentration:A review[J].Chinese Journal of Applied Ecology,2008,19(4):901-910.
[18]Wang Y X,Frei M.Stressed food:The impact of abiotic environmental stresses on crop quality[J].Agriculture,Ecosystems and Environment,2011,141(3):271-286.
[19]Jing L Q,Dombinov V,Shen S B,et al.Physiological and genotypespecific factors associated with grain quality changes in rice exposed to high ozone[J].Environmental Pollution,2016,210:397-408.
[20]王云霞,王晓莹,杨连新,等.臭氧胁迫使两优培九倒伏风险增加:FACE研究[J].生态学报,2011,31(20):6098-6107.
WANG Yun-xia,WANG Xiao-ying,YANG Lian-xin,et al.Ozone stress increases lodging risk of rice cultivar Liangyoupeijiu:A FACE study[J].Acta Ecologica Sinica,2011,31(20):6098-6107.
[21]王云霞,王晓莹,周晓冬,等.近地层臭氧(O3)浓度升高对水稻武运粳21抗倒性的影响[J].江苏农业学报,2011,27(6):1167-1173.
WANG Yun-xia,WANG Xiao-ying,ZHOU Xiao-dong,et al.Impactsof tropospheric ozone concentration enrichment on lodging resistance of rice Wuyunjing 21[J].Jiangsu Journal of Agriculture Science,2011,27(6):1167-1173.
[22]Shimono H,Okada M,Yamakawa Y,et al.Lodging in rice can be alleviated by atmospheric CO2enrichment[J].Agriculture,Ecosystems and Environment,2007,118(1):223-230.
[23]何永美,湛方栋,祖艳群,等.大田增强UV-B辐射对元阳梯田地方水稻茎秆性状和倒伏指数的影响[J].应用生态学报,2015,26(1):39-45.
HE Yong-mei,ZHAN Fang-dong,ZU Yan-qun,et al.Effects of enhanced UV-B radiation on culm charateristics and lodging index of two local rice varieties in Yuanyang terraces under field condition[J].Chinese Journal of Applied Ecology,2015,26(1):39-45.
[24]赵轶鹏,邵在胜,宋琪玲,等.一种新型自然光气体熏蒸平台:系统结构与控制精度[J].农业环境科学学报,2012,31(11):2082-2093.
ZHAO Yi-peng,SHAO Zai-sheng,SONG Qi-ling,et al.System structure and control accuracy of a solar-illuminated gas fumigation platform[J].Journal of Agro-Environment Science,2012,31(11):2082-2093.
[25]穆海蓉,邵在胜,沈士博,等.臭氧浓度增加对超级稻南粳9108稻穗不同部位籽粒氨基酸含量的影响[J].农业环境科学学报,2017,36(3):420-427.
MU Hai-rong,SHAO Zai-sheng,SHEN Shi-bo,et al.Impacts of ozone stress on grain amino acids of super rice cultivar Nanjing 9108 differ with grain positions on apanicle[J].Journal of Agro-Environment Science,2017,36(3):420-427.
[26]邵在胜,沈士博,贾一磊,等.臭氧胁迫对不同敏感型水稻生长和产量形成的影响[J].中国农业科学,2016,49(17):3319-3331.
SHAO Zai-sheng,SHEN Shi-bo,JIA Yi-lei,et al.Impact of ozone stress on growth and yield formation of rice genotypes with different ozone sensitivity[J].Scientia Agricultura Sinica,2016,49(17):3319-3331.
[27]Wang Y X,Yang L X,Meike H,et al.Pyramiding of ozone tolerance QTLs OzT8 and OzT9 confers improved tolerance to season-long ozone exposure in rice[J].Environmental and Experimental Botany,2014,104:26-33.
[28]顾世梁,莫惠栋.动态聚类的一种新方法:最小组内平方和法[J].江苏农学院学报,1989,10(4):l-8.
GU Shi-liang,MO Hui-dong.A new dynamic clustering method:MinSSwmethod[J].Journal of Jiangsu Agriculture College,1989,10(4):l-8.
[29]Frei M,Tanaka J P,Wissuwa M.Genotypic variation in tolerance to elevated ozone in rice:Dissection of distinct genetic factors linked to tolerance mechanisms[J].Journal of Experimental Botany,2008,59(13):3741-3752.
[30]邵在胜,穆海蓉,赵轶鹏,等.臭氧胁迫对不同敏感型水稻叶片伤害的比较研究[J].中国水稻科学,2017,31(2):175-184.
SHAO Zai-sheng,MU Hai-rong,ZHAO Yi-peng,et al.A comparative study of ozone-induced leaf injury of rice with different ozone sensitivity[J].Chinese Journal of Rice Science,2017,31(2):175-184.
[31]申广勒,石英尧,黄艳玲,等.水稻抗倒伏特性及其与茎秆性状的相关性研究[J].中国农学通报,2007,23(12):58-62.
SHEN Guang-le,SHI Ying-yao,HUANG Yan-ling,et al.Study on rice lodging resistance character and correlation between the culm traits and lodging resistance traits[J].Chinese Agricultural Science Bulletin,2007,23(12):58-62.
[32]马 均,马文波,田彦华,等.重穗型水稻植株抗倒伏能力的研究[J].作物学报,2004,30(2):143-148.
MA Jun,MA Wen-bo,TIAN Yan-hua,et al.The culm lodging resistance of heavy panicle type of rice[J].Acta Agronomica Sinica,2004,30(2):143-148.
[33]张喜娟,李红娇,李伟娟,等.北方直立穗型粳稻抗倒性的研究[J].中国农业科学,2009,42(7):2305-2313.
ZHANG Xi-juan,LI Hong-jiao,LI Wei-juan,et al.The lodging resistance of erect panicle Japonica rice in Northern China[J].Scientia A-gricultura Sinica,2009,42(7):2305-2313.
[34]Kashiwagi T,Ishimaru K.Identification and functional analysis of a locus for improvement of lodging resistance in rice[J].Plant Physiology,2004,134(2):676-683.
[35]刘立军,王康君,葛立立,等.旱种水稻基部节间性状与倒伏的关系及其生理机制[J].作物学报,2012,38(5):848-856.
LIU Li-jun,WANG Kang-jun,GE Li-li,et al.Relationship between characteristics of basal internodes and lodging and its physiological mechanism in dry-cultivated rice[J].Acta Agronomica Sinica,2012,38(5):848-856.
[36]张丰转,金正勋,马国辉,等.灌浆成熟期粳稻抗倒伏性和茎鞘化学成分含量的动态变化[J].中国水稻科学,2010,24(3):264-270.
ZHANG Feng-zhuan,JIN Zheng-xun,MA Guo-hui,et al.Dynamic changes of lodging resistance and chemical component contents in culm and sheaths of japonica rice during grain filling[J].Chinese Journal of Rice Science,2010,24(3):264-270.
Impacts of ozone stress on stem lodging resistance of rice genotypes with varying ozone sensitivities
SHAO Zai-sheng1,MU Hai-rong1,ZHAO Yi-peng1,2,JIA Yi-lei1,WANG Yun-xia3,YANG Lian-xin1*,WANG Yu-long1*
(1.Jiangsu Key Laboratory of Crop Genetics and Physiology/Co-Innovation Center for Modern Production Technology of Grain Crops,Yangzhou University,Yangzhou 225009,China;2.Xuzhou Institute of Agricultural Sciences of the Xuhuai District of Jiangsu Province,Xuzhou 221121,China;3.College of Environmental Science and Engineering,Yangzhou University,Yangzhou 225009,China)
From a week after transplanting until maturity,23 rice cultivars or lines were grown in glasshouse-type fumigation chambers,with a control group receiving low concentrations of ozone(C-O3,10 nL·L-1)and an elevated O3treatment group receiving high concentrations of ozone(E-O3,100 nL·L-1).Based on ozone-induced changes in the aboveground biomass,these rice genotypes were clustered,using the MinSSw method(a dynamic clustering method using minimum sum of squares within groups)into three types,namely A(-19%),B(-39%),and C(-52%)in ascending order of ozone sensitivity.Ozone stress decreased the breaking resistance of the third internode from the top of rice types A,B,and C by 19%to 20%,but decreased breaking resistance in the fourth internode from the top of the three rice types by 28%,36%,and 44%respectively.Ozone-induced changes in pushing resistance of the lower part of a single plant showed a trend similarto that of the fourth internode from the top.Ozone stress significantly decreased the length,dry weight,dry weight per unit length,cross-sectional area,maximum stem diameter,minimum stem diameter,and culm wall thickness of the basal internodes,with the dry weight of the internode showing the greatest decrease(on average 58%),whereas the maximum stem diameter,minimum stem diameter,and culm wall thickness showed the least decline(on average 13%to 15%).In most cases,the ozone-induced changes in the seven parameters of the fourth internode from the top were greater than those of the third internode from the top,and ozone-internode interactions were significant for the length,dry weight per unit length,cross-sectional area,maximum stem diameter,and minimum stem diameter of the basal internodes.Among the different rice types,the ozone-induced reduction in each measured parameter followed the pattern C>B>A,and ozonerice type interactions were significant for the cross-sectional area,maximum stem diameter,and minimum stem diameter of the basal internodes.Correlation analysis showed that the response of basal internode breaking resistance to ozone stress was positively correlated with cross-sectional area,maximum stem diameter,minimum stem diameter,or dry weight per unit length of the corresponding internodes.These results indicated that ozone fumigation at concentrations of 100 nL·L-1deteriorated the culm quality of the different types of rice.Ozone stress significantly decreased the breaking resistance of the rice stem,which was mainly associated with basal internode thinning,and also with the decline of dry weight per unit length of the basal internodes.
rice;ozone;lodging resistance;plant internode breaking resistance;paddy growth
S511
A
1672-2043(2017)09-1703-09
10.11654/jaes.2017-0465
邵在胜,穆海蓉,赵轶鹏,等.臭氧胁迫对不同敏感型水稻茎秆抗倒性状的影响[J].农业环境科学学报,2017,36(9):1703-1711.
SHAO Zai-sheng,MU Hai-rong,ZHAO Yi-peng,et al.Impacts of ozone stress on stem lodging resistance of rice genotypes with varying ozone sensitivities[J].Journal of Agro-Environment Science,2017,36(9):1703-1711.
2017-03-30
邵在胜(1989—),男,江苏扬州人,博士研究生,主要从事大气变化与作物响应方面的研究。E-mail:460180389@qq.com
*通信作者:杨连新E-mail:lxyang@yzu.edu.cn;王余龙E-mail:ylwang@yzu.edu.cn
国家自然科学基金面上项目(31371563,31471437,31171460);江苏高校优势学科建设工程项目
Project supported:The National Natural Science Foundation of China(31371563,31471437,31171460);The Priority Academic Program Development of Jiangsu Higher Education Institutions
