黄芩苷与人源RANKL胞外结构域相互作用的研究
2017-09-16陈相如汪劲松王卫东张新潮潘继承
陈相如,汪劲松,王卫东,张新潮,潘继承
(1.湖北师范大学 生命科学学院,湖北 黄石 435002; 2.食用野生植物保育与利用湖北省重点实验室,湖北 黄石 435002)
黄芩苷与人源RANKL胞外结构域相互作用的研究
陈相如1,2,汪劲松1,2,王卫东1,2,张新潮1,2,潘继承1,2
(1.湖北师范大学 生命科学学院,湖北 黄石 435002; 2.食用野生植物保育与利用湖北省重点实验室,湖北 黄石 435002)
细胞核因子κB受体活化因子配体(receptor activator of NF-kappa B ligand,RANKL)源于成骨细胞,对于破骨细胞分化和激活具有重要作用。RANKL能与破骨细胞表面受体RANK(receptor activator of NF-kappa B)结合,骨保护素(osteoprotegerin,OPG)作为诱骗受体竞争性地与RANKL结合,两者共同调节骨平衡。两者调节异常会引起骨形成与骨吸收失衡,导致骨代谢性疾病发生,如骨质疏松症、关节炎等。黄芩苷,是从传统中药黄芩中提取的黄酮类化合物,在医药行业中被广泛应用。荧光光谱法实验表明:在体外黄芩苷对RANKL胞外结构域的内源荧光有明显的猝灭作用,黄芩苷使RANKL内部生色基团所处微环境的极性增大;蛋白ANS荧光峰强度随着黄芩苷的浓度先增后降,在8×10-5mol/L时达到峰值,说明RANKL疏水面暴露程度可能先增大后减小,在8×10-5mol/L时,疏水面暴露程度最大;分子模拟结果显示:黄芩苷与人源RANKL/OPG-CRD复合物之间有较强的相互作用,且位于该复合物中心内部空腔,说明两者结合紧密,预测了两者相互作用的氨基酸位点。但黄芩苷是否抑制RANKL-RANK结合需进一步研究。研究黄芩苷与RANKL相互作用对阐明调节骨代谢紊乱及抗癌机理具有重要意义。
RANKL/RANK/OPG;黄芩苷;荧光光谱法;分子模拟
骨平衡由成骨细胞的骨形成功能和破骨细胞的骨吸收功能共同调节。细胞核因子κB受体活化因子配体(receptor activator for nuclear factor κB ligand,RANKL)是一种Ⅱ型跨膜蛋白,由成骨细胞产生,对于破骨细胞分化和激活具有重要作用[1]。细胞核因子κB活化因子受体(receptor activator for nuclear factor κB,RANK)是一种Ⅰ型跨膜蛋白,在破骨细胞表面表达,与其配体RANKL都属于肿瘤坏死因子家族(tumor necrosis factor receptor-ligand family),介导破骨细胞生成[2]。骨保护素(osteoprotegerin,OPG)作为诱骗受体竞争性地与RANKL结合,能有效地抑制破骨细胞的成熟与激活[3-4]。RANKL与OPG的结合比能很好地调节骨代谢的方向,即骨形成或骨吸收。因此,RANKL与OPG的结合比调节异常会引起骨形成与骨吸收失衡,导致骨代谢紊乱性疾病发生,如骨质疏松症、类风湿性关节炎等[5-8]。在发展治疗骨吸收过量引起的骨代谢紊乱性疾病时,RANKL/RANK/OPG系统是调控破骨细胞生成的关键点,同时也成为公认的靶标[9]。 RANKL/RANK/OPG系统还参与了癌细胞的增殖与转移,乳腺癌是女性患病和死亡人数最高的恶性肿瘤,远处转移是乳腺癌致死的主要原因,阻断RANKL与RANK的结合可以达到间接抗癌的疗效。
黄芩苷(Baicalin)是黄芩主要活性成分之一,属于黄酮类化合物,具有降压、镇静、抗菌、消炎、解热、抗病原体、抗癌、修复脑损伤、保肝利胆、改善糖尿病肾病等多种药理作用[10]。近年来,黄芩苷在口腔医学领域研究较多,尤其是治疗牙周炎方面有明显效果,黄芩苷可以有效的抑制 LPS 诱导的大鼠牙周炎牙槽骨的吸收,可以降低人牙周膜细胞表面RANKL/OPG比值等[11]。Guo A J等人报道,黄芩苷通过Wnt/β-catenin诱导成骨细胞的分化,增强OPG mRNA表达水平,直接参与破骨细胞的活性的调控[12]。黄芩苷在骨质疏松症预防与治疗领域研究较少,由于黄芩苷可以调控RANKL/OPG表达,从理论上可以推测黄芩苷可以成为骨质疏松症的潜在药物。最近,桂林源等人(2016年5月)发现,用黄芩提取物(以总黄酮含量计算)对雌性骨质疏松模型SD大鼠进行腹腔注射后,骨体积和骨小梁数目增加,促进了骨髓间充质干细胞的增殖和成骨分化,可以在一定程度上预防和治疗雌激素缺乏所致的骨质疏松[13]。
在人源RANKL/RANK信号通路中,黄芩苷与RANKL相互作用的研究尚无相关研究报道。为使传统中药黄芩在预防和治疗骨质疏松方面具有更深入的研究,充分发挥其药用价值,达到旧药新用的目的,同时为体外研究人源RANKL的结构提供依据。因此,挖掘黄芩有效成分新功能以及深入研究人源RANKL的结构十分必要。本项研究采用荧光光谱法和分子模拟,研究黄芩苷与人源RANKL胞外结构域相互作用的荧光猝灭,同时预测黄芩苷与RANKL/RANK复合物相互作用的活性位点,尝试解释两者的作用机理,但黄芩苷是否抑制RANKL与RANK结合以及临床效果需进一步研究。
1 材料与方法
1.1 实验材料
载体和菌种:pGEX-6P-1和E.coli Rosseta(DE3),由本实验室保存。
1.2 试剂与仪器
试剂:黄芩苷(黄石市药品检验所提供);氨苄霉素(Amp)、异丙基-β-D-硫代吡喃半乳糖苷(IPTG)等购自Amresco;1-苯胺基萘-8-磺酸(ANS)、苯甲基磺酰氟(PMSF)均购自Sigma;Ni亲和层析柱填料购自GE。
实验中常见溶液的配制参见《分子克隆(第三版)》[14]。
仪器:荧光分光光度计F-4500(Hitachi,Japan)、XO-650超声波细胞粉碎机(南京先欧生物科技有限公司)、HD-3紫外检测仪(上海沪西分析仪器厂有限公司)、YC-1型电脑恒温层析柜(北京博医康实验仪器有限公司)。
1.3 人源RANKL胞外结构域的表达与纯化
本实验室已将人源RANKL(141-317)号位氨基酸的活性区域重组至原核表达载体pGEX-6P-1,并通过表达菌株E.coliRosseta(DE3)进行人源RANKL胞外结构域(RANKL ectodomain,eRANKL)的诱导、表达与纯化。操作过程中的具体条件参照文献[15]。
1.4 黄芩苷与RANKL相互作用的荧光光谱法测定
1.4.1 黄芩苷工作液的配置
用DMSO作溶剂配置浓度为0.01 mol/L的黄芩苷母液,用0.01 mol/L PBS逐级稀释得到0、2、4、6、8、10、20、40、60、80、100×10-5mol/L的工作液。保证每组中DMSO和PBS量一致,黄芩苷浓度为单一变量,配置体系见表1。
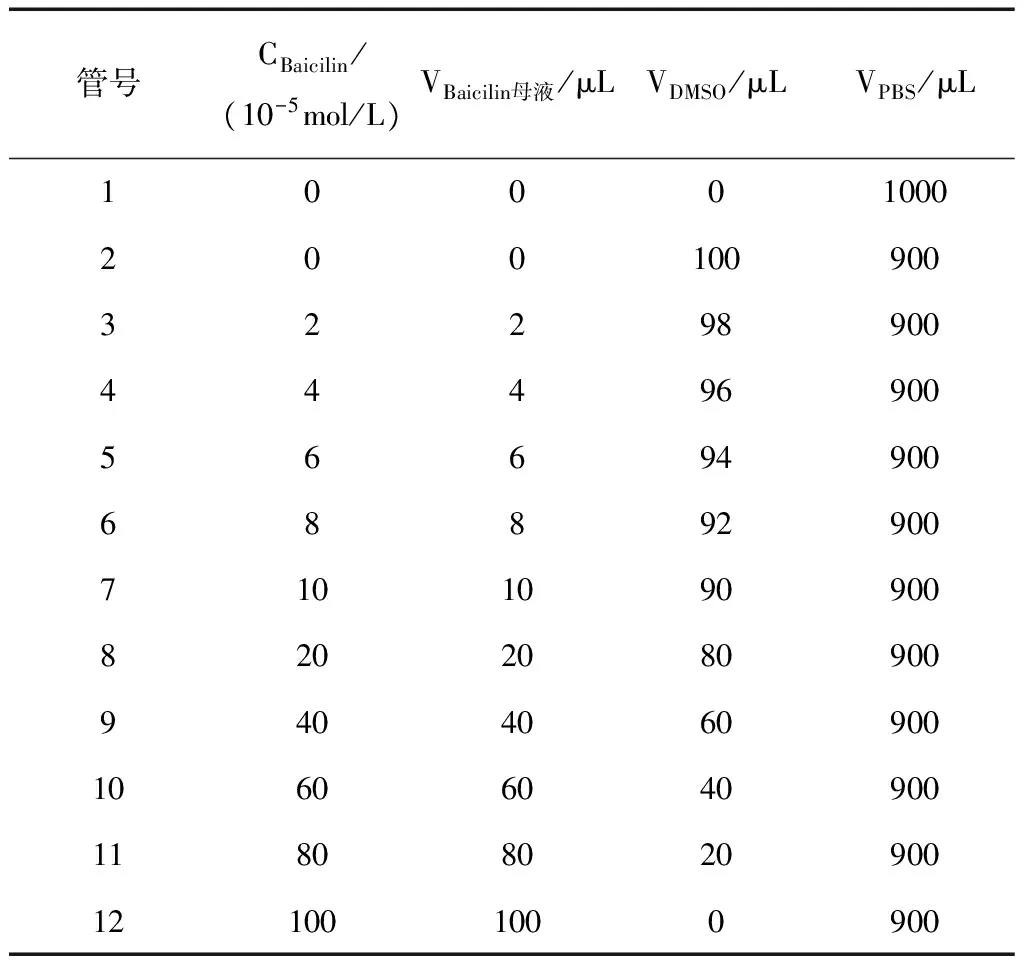
表1 黄芩苷浓度体系(1 mL)
1.4.2 内源荧光的测定
调整RANKL浓度为22.56 μmol/L(A280=0.8),采用表1中黄芩苷浓度梯度。固定反应体系中RANKL蛋白浓度,增加黄芩苷用量,考察RANKL的荧光强度变化。反应体系见表2。

表2 内源荧光反应体系
分别在10、20、30℃水浴30 min后,进行荧光光谱扫描,激发光波长295 nm,激发狭缝2.5 nm,发射狭缝5 nm,电压700V,扫描速度1200 nm/min,记录300~400 nm波段光谱数据。
1.4.3 外源荧光的测定
调整RANKL浓度为22.56 μmol/L(A280=0.8),采用表1中黄芩苷浓度梯度。避光称取ANS,蒸馏水溶解,离心(12000 r/min,5min),避光取上清,立即使用。ANS浓度为蛋白50倍,按1:100加入蛋白溶液。反应体系见表3。
30℃水浴避光反应30min后,进行荧光光谱扫描,激发光波长380 nm,激发和发射狭缝均设为5 nm,电压700V,扫描速度1200 nm/min,记录400~600 nm波段光谱数据。

表3 ANS荧光反应体系
1.5 黄芩苷与人源RANKL/OPG复合物的分子模拟
人源RANKL与人源OPG上4个半胱氨酸富集区(cysteine rich domain,CRD)结合所形成的复合体(RANKL /OPG-CDR)的3D晶体结构是从PDB (http://www.pdb.org/,PDB ID: 3URF)中获取。参照文献[16]方法,在Sybyl-X 2.1.1中对蛋白和黄芩苷优化,柔性对接。
2 结果和分析
2.1 RANKL表达与纯化

M:Marker;0:上清;1:40 mmol/L;2:80 mmol/L;
由图1看出,通过Ni亲和层析在150 mmol/L和200 mmol/L洗脱下来的蛋白经过SDS-PAGE的检测,目标蛋白约为21 KD,且均一性较高。
2.2 内源荧光光谱
2.2.1 黄芩苷对RANKL内源荧光的猝灭作用
在本实验条件下,荧光谱340 nm处有一荧光发射峰,不同温度下的实验结果见图2。
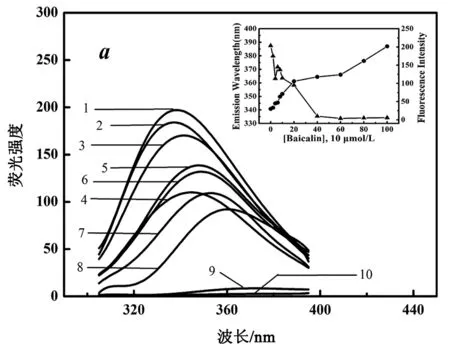

图(a)、(b)、(c)分别表示在10℃、20℃以及30℃温度下,黄芩苷对RANKL作用,RANKL浓度为22.56 μmol/L,1~10分别代表表1中黄芩苷的浓度梯度。内插图中“▲”表示最大荧光发射峰强度,“●”表示最大荧光发射峰强度对应的发射波长
图2 三种温度下黄芩苷与RANKL相互作用的内源荧光图谱
由图2可以看出,随着黄芩苷浓度增加,RANKL荧光发射峰强度减弱,发射波长明显红移,说明黄芩苷使RANKL内部生色基团所处微环境的极性增大,RANKL构象发生变化。
2.3 外源荧光图谱
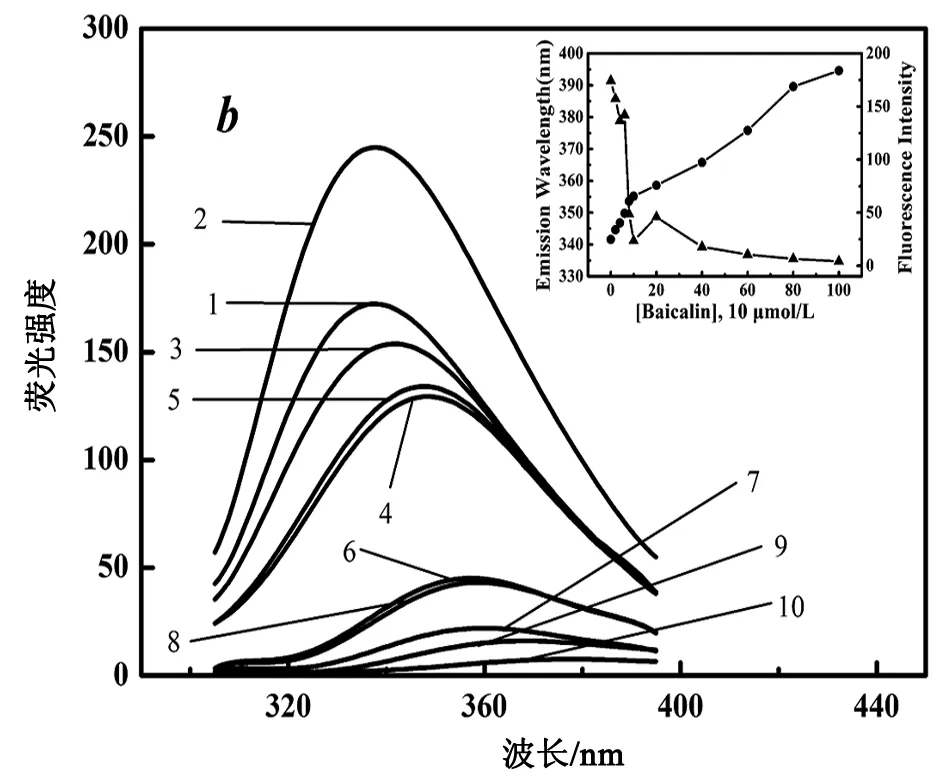
ANS通过非共价结合蛋白分子的非极性区域中,其荧光光谱会随着非极性的增加而发生蓝移,且荧光强度也会随之提高。30℃下,黄芩苷与RANKL相互作用的ANS荧光见图3。

RANKL浓度为22.56 μmol/L,1~12分别代表表1中黄芩苷的
实验结果显示,随着黄芩苷浓度的增加,蛋白ANS荧光峰强度先逐渐增加后逐渐下降,在8×10-5mol/L时达到峰值。说明RANKL疏水面暴露程度先增大后减小,在8×10-5mol/L时,疏水面暴露程度最大。
2.4 黄芩苷与人源RANKL/OPG复合物的分子模拟
Xudong Luan等人发表的人源RANKL结构域与OPG中4个半胱氨酸富集区所形成的晶体结构用于与黄芩苷分子对接,其中OPG中4个半胱氨酸的富集区(OPD-CRD)与人源RANK胞外结构域中4个半胱氨酸的富集区(RANK-CRD)序列相同[17],这也是OPG能作为RANKL诱骗受体的主要原因。分子对接全局和局部情况见图4,作用位点信息见表4。
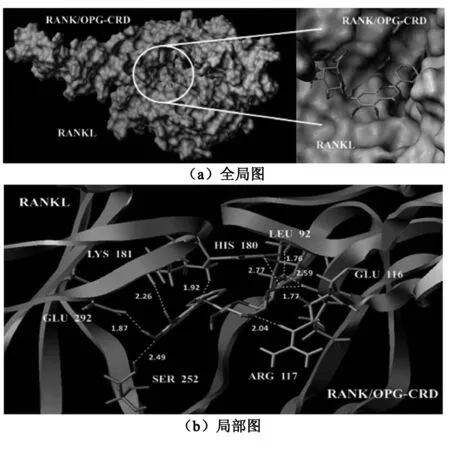
(a)全局图展示了黄芩苷在RANKL/OPG-CRD复合物中的具体位置;(b)局部图显示了黄芩苷分别在RANKL和OPG-CRD分别作用的氨基酸位点,黄色虚线表示氢键,黄色数字表示氢键键长,蓝色β-折叠带来自RANKL,绿色β-折叠带来自OPG-CRD。

图4 黄芩苷与人源RANKL/OPG-CRD复合物分子对接
黄芩苷与人源RANKL/OPG-CRD复合物分子对接的总得分为7.638,属于高分,两者作用的氢键有9个,表明黄芩苷与RANKL/OPG-CRD间氢键相互作用力较强,结合紧密。从对接的全局图来看,黄芩苷恰好被复合物内部中心空腔包裹。从局部图来看具体作用的氨基酸位点,很可能是黄芩苷发挥作用的位点。分子模拟结果为黄芩苷阻止RANKL/RANK复合物形成的机制提供理论依据。
3 结论
我们通过研究黄芩苷影响RANKL胞外结构域的聚集状态,进而影响RANKL/RANK通路的形成。荧光光谱学数据表明:在体外,黄芩苷对RANKL内源荧光强度的减弱均呈剂量依赖关系,黄芩苷对RANKL的内源荧光有明显的猝灭作用,黄芩苷使RANKL内部生色基团所处微环境的极性增大;蛋白ANS荧光峰强度与黄芩苷浓度并无剂量依赖关系,而是先增加后降低,在8×10-5mol/L时达到峰值,说明RANKL疏水面暴露程度可能先增大后减小,在8×10-5mol/L时,疏水面暴露程度最大。分子模拟结果显示,黄芩苷与人源RANKL/OPG-CRD复合物之间有较强的相互作用,且位于该复合物中心内部空腔,说明两者结合紧密,预测了两者相互作用的氨基酸位点。推测黄芩苷可能有利于阻止RANKL-RANK复合物的形成,但在体外还需要通过等温滴定量热法进一步证明。黄芩苷在细胞水平和动物水平是否有效还需进一步深入研究,同时可以结合结构生物学方法准确地找出两者结合的具体位置以及开展动物学实验。
综上所述,在体外黄芩苷能影响RANKL的结构变化,可能影响RANKL与RANK的结合,从而可能阻止RANKL/RANK信号通路的开启,达到间接治疗骨质疏松和抗癌药效。因此,黄芩苷可能成为骨质疏松、乳腺癌等疾病的潜在药物。
[1] Willard D,Chen W J,Barrett G,et al. Expression,purification,and characterization of the human receptor activator of NF-kB ligand(RANKL) extracellular domain[J]. Protein Expr Purif,2000,20:48-57.
[2] Ta H M,Nguyen G T,Nguyen G T,et al. Structure-based development of a receptor activator of nuclear factor-κB ligand (RANKL) inhibitor peptide and molecular basis for osteopetrosis[J]. Proc Natl Acad Sci USA,2010,107:20281-20286.
[3] Simonet W S,Lacey D L,Dunstan C R,et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density[J]. Cell,1997,89:309-319.
[4] Yasuda H,Shima N,Nakagawa N,et al. Identity of osteoclastogenesis inhibitory factor (OCIF) and osteoprotegerin (OPG): A mechanism by which OPG/OCIF inhibits osteoclastogenesis in vitro[J]. Endocrinology,1998,139:1329-1337.
[5] Tanaka S,Nakamura K,Takahasi N,et al. Role of RANKL in physiological and pathological bone resorption and therapeutics targeting the RANKL-RANK signaling system[J]. Immunol Rev,2005,208:30-49.
[6] Vega D,Maalouf N M,Sakhaee K. CLINICAL Review: The role of receptor activator of nuclear factor-kappaB (RANK)/RANK ligand/osteoprotegerin: clinical implications[J].J Clin Endocrinol Metab,2007,92:4514-4521.
[7] Theoleyre S,Wittrant Y,Tat S K,et al. The molecular triad OPG/RANK/RANKL: Involvement in the orchestration of pathophysiological bone remodeling[J]. Cytokine Growth Factor Rev,2004,15:457-475.
[8] Wright H L,McCarthy H S,Middleton J,et al. RANK,RANKL and osteoprotegerin in bone biology and disease[J]. Curr Rev Musculoskelet Med,2009,2:56-64. [9] Hur J,Ghosh A,Kim K,et al. Design of a RANK-mimetic peptide inhibitor of osteoclastogenesis with enhanced RANKL-binding affinity[J]. Mol Cells,2016,39:316-321.
[10] 欧阳俊杰. 黄芩苷及其衍生物抗病原体作用研究新进展[J]. 宜春学院学报,2015,37:28-30.
[11] 陈 悦.黄芩苷抑制牙周炎牙槽骨吸收的作用及机理研究[D].西安:第四军医大学,2008.
[12] Guo A J,Choi R C,Cheung A W,et al. Baicalin,a flavone,induces the differentiation of cultured osteoblasts[J]. J Biol Chem,2011,286:27882-27893.
[13] 桂林源,刘世宇,邱新毓,等. 黄芩小分子化合物对雌激素缺乏骨质疏松的预防和治疗作用[J].牙体牙髓牙周病学杂志,2016,26(5):294-300.
[14] 莎姆布鲁克 J,拉塞尔 D W.分子克隆实验指南[M].3版.黄培堂,译.北京:科学出版社,2002.
[15] 习雪丽. 生物活性多糖及黄酮类化合物与人源RANKL胞外结构域相互作用的研究[D].黄石:湖北师范大学,2014.
[16] Mi R,Li P Q,Hu Y J,et al. Biophysical studies on the interactions of jatrorrhizine with bovine serum albumin by spectroscopic and molecular modeling methods[J]. Mol Biol Rep,2013,40:4397-4404.
[17] Luan X,Lu Q,Jiang Y,et al. Crystal structure of human RANKL complexed with its decoy receptor osteoprotegerin[J]. J Immunol,2012,189:245-252.
(本文文献格式:陈相如,汪劲松,王卫东,等.黄芩苷与人源RANKL胞外结构域相互作用的研究[J].山东化工,2017,46(7):56-60.)
The Study of the Interaction Between Baicalin and Human RANKL Ectodomain
ChenXiangru1,2,WangJinsong1,2,WangWeidong1,2,ZhangXinchao1,2,PanJicheng1,2*
(1.College of Life Science, Hubei Normal University; Huangshi 435002,China; 2.Hubei Key Laboratory of Edible Wild Plants Conservation and Utilization, Huangshi 435002,China)
RANKL (receptor activator of NF-kappa B ligand), steming from osteoblast, play an important role in the osteoclast differentiation and activation. RANKL can bind osteoclast surface receptor RANK(receptor activator of NF-kappa B), and OPG (osteoprotegerin), as decoy receptor,competitively combine with RANKL,which they jointly regulate the bone balance. The RANKL/RANK/OPG dysregulation causes the imbalance between bone formation and resorption,which results in bone metabolic diseases,such as osteoporosis,arthrophlogosis,et al. Baicalin,a kind of flavonoid extracted from Scutellaria baicalensis Georigi―a traditional Chinese medicine,are widely applied in the field of medicine. Fluorescence spectral measurements data showed that with the baicalin’s concentration increasing,accompanied with the fluorescence emission intensity dropping,the emission wavelength red-shift occurred. Further,ANS fluorescence data also showed that baicalin could result emission intensity obvious change. Thus,combined with intrinsic and ANS fluorescence evidences,and an conclusion seemed to be drawn that baicalin could cause RANKL’s conformational change. The results of molecular simulation revealed the existence of combination between baicalin and human RANKL/OPG-CRD (cysteine rich domain) ,which located in the inner cavity of RANKL/OPG-CRD compound,and predict the concrete amino acid residue sites of the stronger binding. The study of the interaction between baicalin and human RANKL carry important implication on the demonstration of the dysregulation of bone metabolism and the anticancer mechanism.
RANKL/RANK/OPG;baicalin;fluorescence spectral measurements;molecular simulation
2017-02-20
陈相如(1990—),男,湖北武穴人,硕士研究生,研究方向:蛋白质结构与功能;通讯作者:潘继承,教授。
TQ461
A
1008-021X(2017)07-0056-05
