丛枝菌根真菌对柳枝稷苗期生长作用机制的研究
2017-09-13程亭亭刘曙东孙风丽奚亚军
徐 鹏, 程亭亭, 张 超, 刘曙东, 孙风丽*, 奚亚军*
(1. 西北农林科技大学农学院, 陕西 杨凌 712100; 2. 西北农林科技大学旱区作物逆境生物学国家重点实验室, 陕西 杨凌 712100)
柳枝稷(PanicumvirgatumL)是禾本科(Gramineae)黍属(Panicum)的多年生C4植物,原产于北美大草原[1]。它具有适应性广、产量高、对环境友好等优点,已成为最理想的能源作物之一[2-3]。但播种后柳枝稷出苗较晚,苗期生长缓慢,导致其与同时期生长的杂草在水分、营养、光照等竞争中处于不利地位[4-5]。如果种植当年不能建成较好的植被群体,会提高柳枝稷的生产成本,降低其经济价值[2]。相关研究发现柳枝稷苗期生长缓慢可能与土壤类型、杂草竞争、种子大小等因素相关[6]。
共生是植物适应环境的一种重要策略。丛枝菌根真菌(Arbuscularmycorrhizalfungi, AMF)可以与大多数陆生植物形成互利共生的内生性菌根,促进植物根系养分的吸收和利用[7]。盆栽试验和大田试验都表明柳枝稷根系也能与真菌共生形成菌根,有助于促进柳枝稷对养分的吸收,提高柳枝稷的生物产量[8-10]。因此菌根共生可能是促进柳枝稷苗期生长的重要手段。
植物苗期的生长需要多方面的协作与调控,其中植物激素起着重要的调控作用。独脚金内酯(strigolactones, SLs)是一类来源于类胡萝卜素的萜内酯,近来被认定为一类新型植物激素,控制植物分枝的发育[11-12]。同时他还能促进丛枝菌根的分枝[13-14],诱导寄生植物如独脚金金(Strigaspp.)和列当(Orobanchespp.)种子的萌发[15]。独脚金内酯途径相关基因的研究取得了一系列进展,至今为止,利用突变体及功能分析法从不同植物中克隆得到SLs合成途径中的4个基因,包括编码类胡萝卜素裂解双加氧酶7的同源基因(MAX3/RMS5/DAD3/D17)、编码类胡萝卜素裂解双加氧酶8的同源基因(MAX4/RMS1/DAD1/D10)、一个编码含铁蛋白的基因(D27/AtD27)、一个编码细胞色素 P450 蛋白的基因(MAX1)。外源施加人工合成的SLs类似物GR24能够恢复这些基因的突变体表型[16]。高粱和玉米与菌根真菌共生后能减轻寄生植物独脚金的侵染[17-18],可能是由菌根形成后独脚金内酯含量的下降所导致[19]。因此独脚金内酯含量的变化很可能是菌根共生过程中的重要调控手段。本文研究了丛枝菌根真菌对柳枝稷苗期生长的影响,并通过测定生理指标及独脚金内酯的含量研究丛枝菌根真菌影响柳枝稷苗期生长的机制。
1 材料与方法
1.1 材料与基质
供试材料为来自于柳枝稷低地型栽培品种‘Alamo’的2个品系Ma和Mg。所用的接种物为Agrauxine公司(英国)商业化的菌根真菌混合孢子,由中国科学院上海生命科学研究院植物生理生态研究所王二涛研究员馈赠。试验于2016年在西北农林科技大学农学院的光照培养箱以及人工气候室进行。将珍珠岩与沙子按1∶2的比例混合,用121℃高温灭菌1 h,平均分成2份。取全新的4×8育苗钵用酒精消毒后晾干,将混合基质装入育苗钵中备用。瓜列当种子由西北农林科技大学水土保持研究所马永清研究员馈赠。
1.2 种子处理及试验设计
将Ma和Mg种子先用10% NaClO处理2 min,再用双蒸水清洗5次后,在灭菌的滤纸上发芽[20]。在30℃培养箱中培养10天后,选取长势一致的幼苗移入育苗钵中,每个育苗钵加入200个左右孢子单位的接种物作为实验组,另一份作为对照组,每种材料种植32株,每个处理设置3个重复。放入光培养12 h,温度28℃,光照强 度20 000 lux;暗培养12 h,温度24℃的循环培养箱中生长,隔天浇1次超纯水。40天后调查菌根侵染及柳枝稷的生长发育状况。
1.3 菌根墨水及WGA染色
参照水稻菌根染色的方法,取柳枝稷根系用墨水染色观察侵染情况[21]。先用10% KOH在95℃水浴锅中处理6 min,然后流水冲洗3遍。随后用墨水染料(100 mL水+5 mL派克标准墨水+5 mL冰醋酸)95℃染色3 min。水冲洗4~5次后静置2~3天,每天换水使其脱色,最后制片并在显微镜下观察。
取柳枝稷根系,用WGA染色的方法观察侵染情况[22]。根系在50%酒精中处理过夜;用水冲洗去除酒精,在20% KOH中室温放置2~3天;用水冲洗去除KOH,在0.1 M HCL中处理1~2小时;用水冲洗去除HCL,用PBS溶液 (每800 mL加入8 g NaCl,0.2 g KCl,1.44 g Na2HPO4,0.24 g KH2PO4)再冲洗2遍;用水冲洗后,加入WGA-Alexafluor 488染液,室温处理6 h左右,制片后在荧光显微镜 (激发波长488 nm)下观察。
1.4 生理指标的测定
将幼苗整株取出,用自来水将根系仔细地冲洗干净,然后用吸水纸将水吸干,分别测定各组的鲜重及干重、根长、株高、叶绿素含量。
根长:测定10株幼苗单株的根长,取其平均值。
鲜重、干重:取10株幼苗测定其鲜重,于105℃下杀青15 min,85℃下烘干至恒重,测定干重。
叶绿素含量:利用叶绿素测定仪(SPAD-502Plus)直接测定叶片中叶绿素含量。
1.5 根系分泌物的提取及瓜列当萌发处理
将幼苗根系清洗干净后进行风干处理,研钵研成粉末。取100 mg粉末加入1.5 mL甲醇,超声30 min后离心取上清作为原液,稀释10倍备用。参照安雨[23]的方法处理瓜列当种子,分别以GR24和去离子水处理为正、负对照。
1.6 RNA提取,cDNA合成及荧光定量PCR
取处理后的柳枝稷根系液氮速冻,用Trizol法抽提RNA,然后参照天根反转试剂盒去除gDNA以及反转录合成cDNA。使用柳枝稷中SLs合成相关基因的定量引物(表1)在伯乐公司的CFX96型实时荧光定量仪中进行扩增。
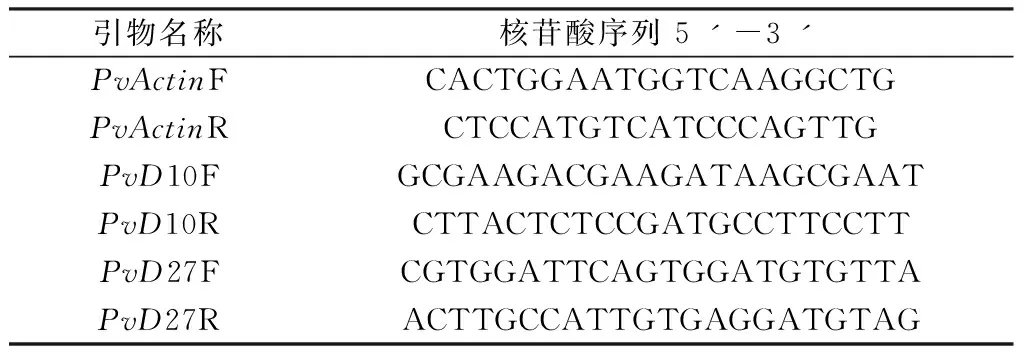
表1 SLs合成相关基因及内参基因定量引物序列Table 1 The primers of gene in SLs synthesis pathway and internal reference gene actin
1.7 数据分析
利用Excel 2010做图表,利用LDS法对各指标进行方差分析。
2 结果与分析
2.1 丛枝菌根真菌侵染的检测
丛枝菌根是内生菌根的主要类型之一,其菌丝没有横膈膜,可在根的内皮层细胞内形成丛枝状结构,其中有些还能在细胞内或皮层细胞间形成泡囊结构。墨水染色(图1)和WGA染色(图2)均能明显观察到含有丛枝状菌丝结构和充满泡囊的细胞,说明菌根真菌侵染成功。对照中则没有观察到菌根真菌侵染的迹象。

图1 柳枝稷菌根侵染墨水染色(A. Ma接菌;B. Mg接菌;C. Ma对照;D. Mg对照)Fig.1 Staining of AMF in switchgrass roots with ink ( A. Ma-AMF; B. Mg-AMF; C. Ma-CK; D. Mg-CK)
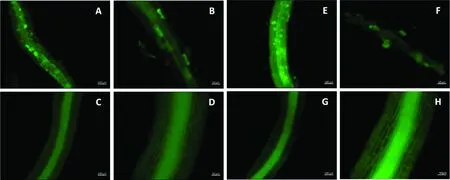
图2 柳枝稷菌根侵染WGA染色(A-B. Ma接菌;C-D. Ma对照;E-F. Mg接菌;G-H. Mg对照)Fig.2 Staining of AMF in switchgrass roots with WGA( A and B. Ma-AMF; C and D. Ma-CK; E and F. Mg-AMF; G and H. Mg-CK)
2.2 丛枝菌根真菌侵染对柳枝稷苗期表型的影响
由表2可知,Ma和Mg 2个品系接种菌根真菌后,除了根长没有变化外,株高、干重、鲜重、叶绿素含量和主茎叶片数等均极显著高于对照(P<0.01),这说明菌根真菌与苗期柳枝稷共生后,通过促进柳枝稷根部养分吸收,加快柳枝稷地上叶片的生长,最终增加柳枝稷的生物量。菌根共生对柳枝稷苗期的生长具有促进作用。
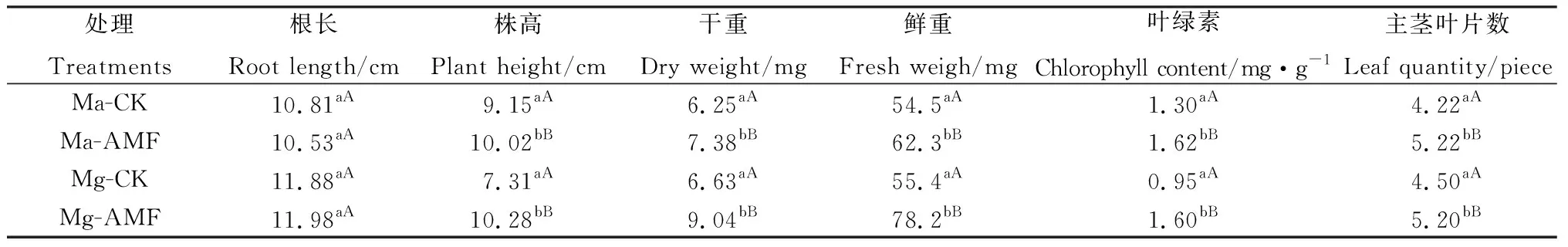
表2 菌根真菌侵染对柳枝稷苗期生长指标的影响Table 2 Effects of Arbuscular Mycorrhizal Fungi on the growth of switchgrass at seedling stage
注:同列不同小写字母表示不同处理间差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)
Note: Different lowercase letters in the same column indicate significant differences at the 0.05 level among different treatments; different capital letters indicate significant differences at the 0.01 level
2.3 丛枝菌根真菌侵染对根系独脚金内酯含量的影响
寄主植物根系分泌的独脚金内酯能够促进独脚金、列当等寄生植物的萌发[15],因此我们利用柳枝稷根系粉末的甲醇提取液处理寄生植物瓜列当的种子,其发芽率作为判断根系独脚金内酯含量的间接指标。作为阳性对照的GR24处理,瓜列当种子萌发率达到了37.7%,而阴性对照H2O处理,瓜列当种子萌发率只有2.8%。比较Ma和Mg 2个品系的处理发现,Ma和Mg未接菌处理的根系提取物处理,瓜列当的萌发率分别达到了22.6%和57.3%,而接菌处理的根系提取物处理后其萌发率分别只有9.6%和19.8%(图3)。由此可以初步推测,接种菌根真菌后,根系中独脚金内酯含量会显著下降,从而导致瓜列当的萌发率只有未接菌的1/3。为进一步验证独脚金内酯的变化,我们选择了Mg接菌和未接菌的根系材料提取RNA,反转cDNA后利用实时荧光定量的方式检测SLs合成相关标记基因的表达情况,利用柳枝稷的Actin基因作为内参基因,结果发现在Mg未接菌材料中,SLs合成相关标记基因D10和D27的表达量显著高于接菌的Mg(图4)。综合以上结果,接种丛枝菌根真菌后柳枝稷根系中独脚金内酯含量会明显减少。
3 讨论
柳枝稷作为新型能源作物而倍受关注,但因其苗期生长缓慢,植株建植时间长,降低了柳枝稷的利用价值,加大了柳枝稷推广的难度。共生是植物适应环境的一种重要策略,丛枝菌根真菌能与大多数陆地上的维管植物形成专性共生体系,促进根部营养吸收以及植物生长发育并增强植物的环境适应能力。因此本试验利用来自于柳枝稷‘Alamo’的Ma和Mg 2个品系为试验材料,接种丛枝菌根真菌的混合孢子,研究菌根共生对柳枝稷苗期生长的影响。结果发现菌根真菌能成功侵染柳枝稷根系。接菌后的柳枝稷株高增加,叶片数增多,生物量显著增长,说明菌根真菌与植株共生后能加快柳枝稷苗期的生长发育。因此,在种植柳枝稷时可通过施撒菌根真菌孢子的方法促进柳枝稷苗期的生长,加快其建植速度。
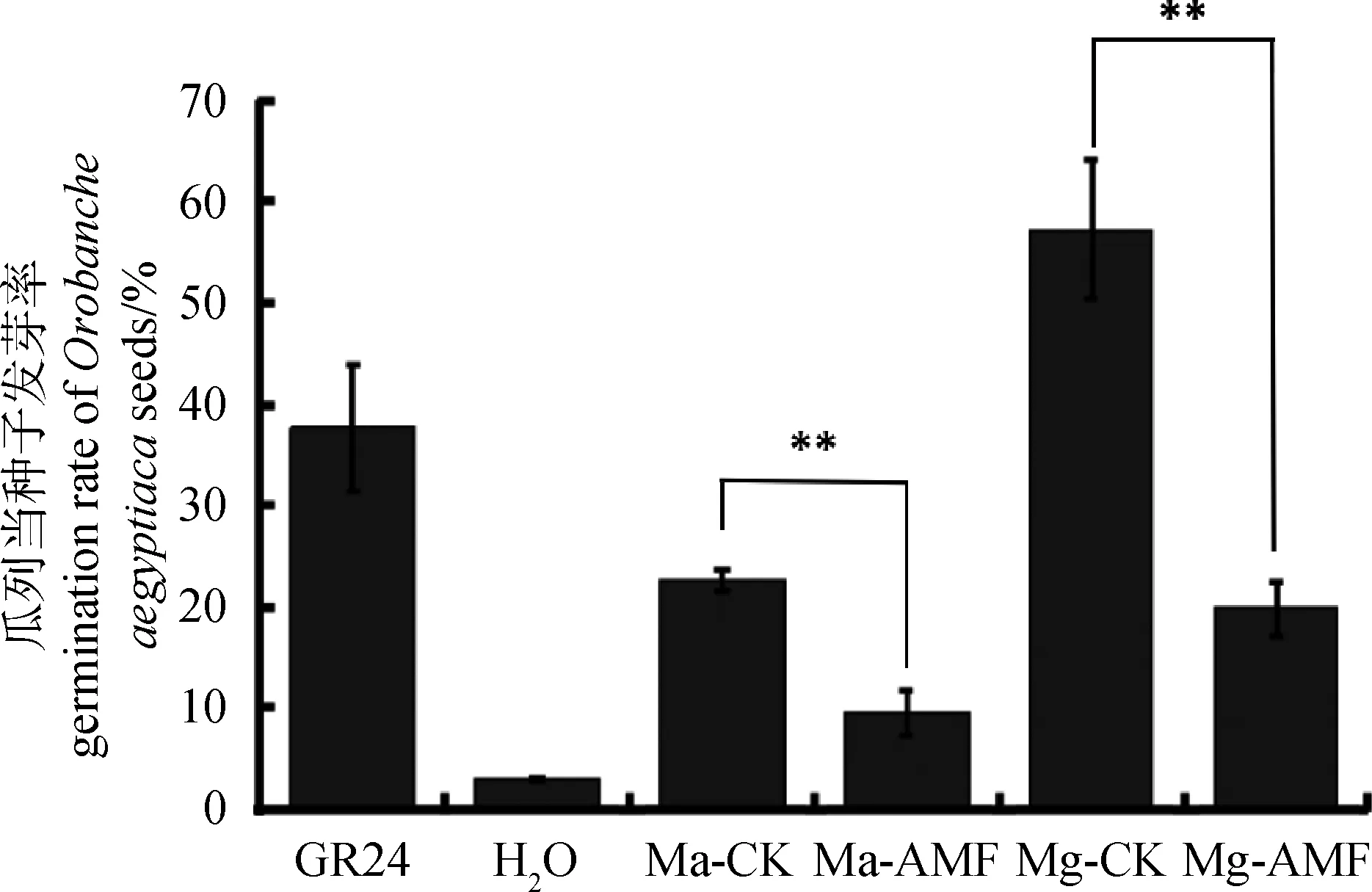
图3 柳枝稷根系甲醇浸提液处理瓜列当的发芽率Fig.3 Germination rate of Orobanche aegyptiacainduced with extracts from switchgrass roots
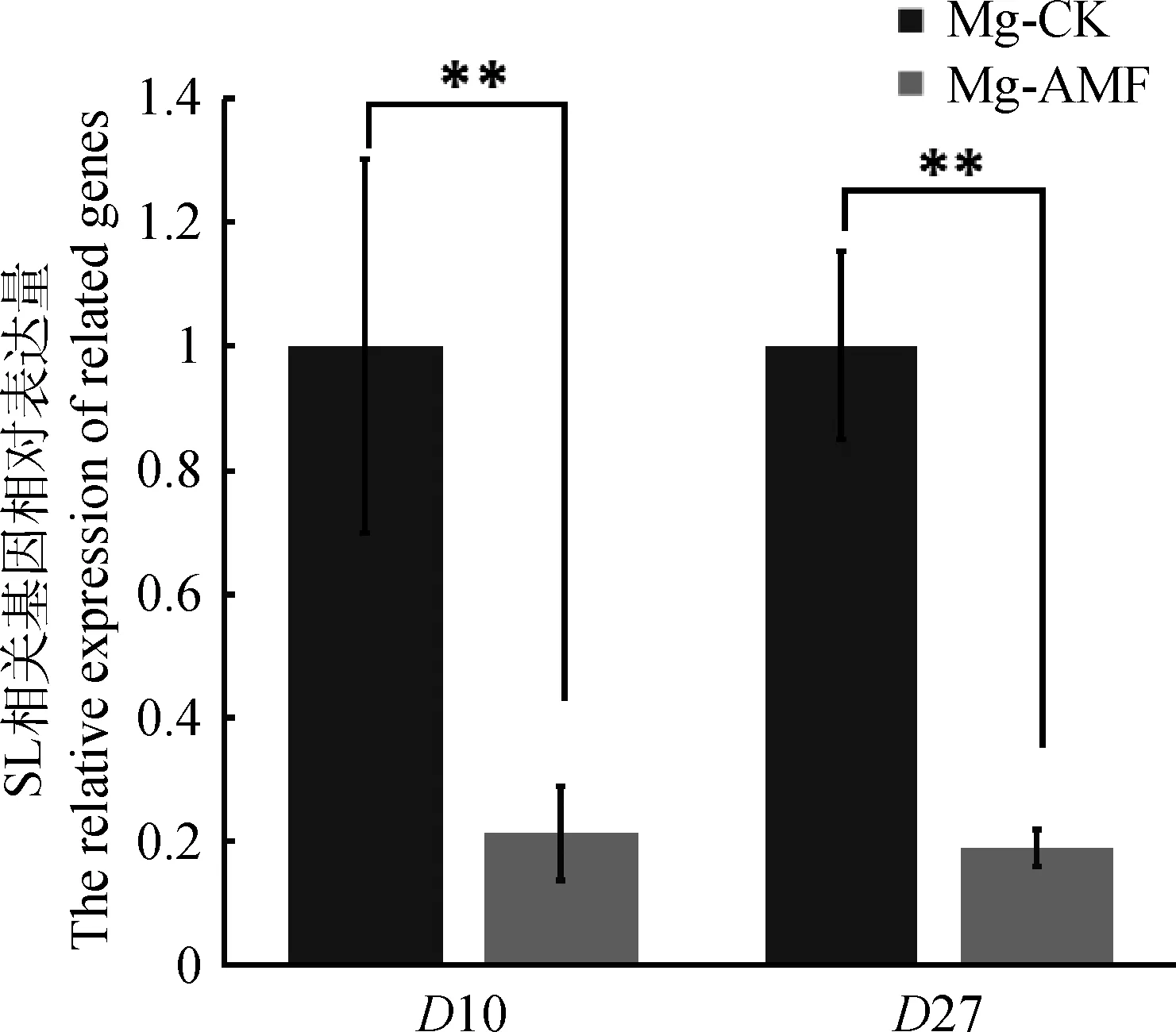
图4 柳枝稷中SLs合成基因表达量检测Fig.4 Expression of genes involved in SLs synthesis in switchgrass
菌根真菌促进柳枝稷苗期生长的过程中,独脚金内酯很可能发挥着重要的调控作用。植物与菌根真菌的共生起始于信号交换,寄主植物根系可能通过分泌一种“分枝因子”(branching factors,BFs)来诱导丛枝真菌菌丝大量分枝,从而增加菌丝与寄主植物根系接触的机会[24]。而真菌自身也会释放一种可扩散的“Myc因子”(myc factor,MF)诱导寄主植物根系相关基因的表达,从而为菌根真菌的侵染提供有利条件[25]。目前BFs已被证实是SLs[13],而MFs的成分目前仍不清楚。寄主植物根系分泌的SLs可以通过诱导一系列的分子和细胞生物学事件,促进丛枝菌丝形成大量分枝。Ma和Mg接种菌根真菌后,其叶绿素含量、生物量、株高与对照相比都有了显著升高,说明当植物与真菌的共生关系形成后,植物的营养状况会因为共生真菌的作用而得到改善。在这种情况下,我们用根系的甲醇提取液处理寄生植物瓜列当,测定其发芽率,发现瓜列当在菌根共生的柳枝稷的提取液处理后,发芽率显著低于未接菌柳枝稷的处理,而SLs的重要作用就是促进寄生植物的萌发。进一步检测Mg根系中SLs合成标记基因的表达水平,发现未接菌的Mg根系中D10和D27的表达量显著高于接菌的Mg。这就说明柳枝稷在共生后会减少独角金内酯的合成,促进植物分枝的形成,对植物的生长有促进作用。相反,未接菌根真菌的柳枝稷由于得不到充分的无机营养,会加速独脚金内酯的分泌,从而诱导土壤中的菌根真菌孢子萌发和促进分枝,以期尽快形成促进营养吸收的菌根结构。
4 结论
以来自于低地型品种Alamo的2个品系Ma和Mg的柳枝稷为材料,研究丛枝菌根真菌对柳枝稷苗期生长发育的影响,研究发现丛枝菌根真菌能够成功侵入苗期柳枝稷的根内,形成有功能的共生菌根,并且能显著增加柳枝稷苗期的株高、叶绿素含量、叶片数和生物量。提取根系分泌物处理寄生植物瓜列当的种子,发现接菌后柳枝稷的根系分泌物会显著降低瓜列当的发芽率,同时观察到Mg接菌后植物体内独脚金内酯合成关键基因D10和D27表达量显著下调,提示柳枝稷形成菌根后会抑制根系独脚金内酯的合成和分泌,从而解除独脚金内酯对柳枝稷分蘖的抑制作用,促进柳枝稷苗期的生长。
