无芒隐子草种子发育过程中不同位置叶片的光合速率及重量与种子产量的相关性
2017-09-13陶奇波白梦杰孙秋瑾韩云华王彦荣
陶奇波, 白梦杰, 孙秋瑾, 韩云华, 王彦荣
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院, 甘肃 兰州 730020)
无芒隐子草(Cleistogenessongorica)为禾本科隐子草属植物,是我国西北地区重要的野生乡土草种[1]。其具有抗旱、耐寒、适口性好等特性,不仅可以作为优良的牧草,而且在城市绿化、荒漠化治理与恢复中具有广阔的应用前景[2]。当前对无芒隐子草的研究主要集中在建植技术[3]、种子萌发生态学[4-6]、围栏封育[7-8]等方面。就其种子生产方面的研究则仅限于栽培密度以及施氮肥两个方面[9-10]。
光合作用是植物完成整个生活史的原初动力,也是植物生长发育的基础和生产力高低的决定性因素[11-12]。然而,目前关于无芒隐子草光合生理方面的研究尚未见报道。大量在农作物上的研究表明,植株不同位叶的光合特性以及生物量存在显著差异,对种子产量的贡献也不同[13-14]。红花(Carthamustinctorius)不同位叶的光合特性存在显著差异,其上位叶净光合速率与单株花产量呈极显著正相关[15]。罗俊[16]等研究表明能源甘蔗(Saccharumofficinarum)分蘖期第3叶的叶宽和水分利用效率以及第5叶的叶宽对产量影响较大。本文通过在无芒隐子草种子发育各时期测定其生殖枝自上而下第1~7叶的净光合速率以及重量,阐明种子发育各时期不同位置叶片的净光合速率、叶片重量以及含水量、穗重量的变化趋势,并探究以上指标与种子产量的相关性,以期为无芒隐子草种子生产提供更加具有针对性的技术指导。
1 材料与方法
1.1 研究区概况
试验地位于兰州大学草地农业科技学院民勤试验站(38°44′N, 103°1′E),海拔1 307 m,年均温为8.3℃,年平均降水量127.7 mm,蒸发量2 623 mm,无霜期162 d,年日照时数约为3 073 h。土壤为沙壤土,0~30 cm土壤速效氮、磷、钾含量分别为31.08, 17.48, 118.36 mg·kg-1,全氮含量为7.76 g·kg-1,有机质含量为12.19 g·kg-1,土壤pH值为8.01。
1.2 试验方法与测定指标
于2016年7月,在行距30 cm、播种后第3年且长势均衡的无芒隐子草种子生产田中随机选取保留了完整7枚叶片的生殖枝为试验材料,并自上而下将7枚叶片分别编为1~7号。
1.2.1不同位叶净光合速率及光合贡献率 在盛花期、灌浆期、乳熟期、蜡熟期以及完熟期随机选取10个生殖枝,于晴朗无风的上午,用LI-6400便携式光合仪,在自然光照下测定每个生殖枝自上而下第1~7叶的净光合速率Pn(net photosynthetic rate),每个叶片测定3个数值。并按照如下公式计算不同位叶的光合贡献率:
光合贡献率(%)=(该叶片净光合速率/第1~7叶总光合速率)×100%
1.2.2不同位叶重量、重量比例以及含水量 在盛花期、灌浆期、乳熟期、蜡熟期以及完熟期随机选取10个生殖枝,按照叶片位置顺序剪取各个生殖枝的叶片,称鲜重后分别装袋,于80oC烘至恒重,用万分之一天平称干重,并按照如下公式计算叶重量比例及叶片含水量:
叶重量比例=(该叶重量/第1~7叶总重量)×100%
叶片含水量=[ (鲜重-干重)/鲜重]×100%
1.2.3种子发育过程中小穗重量 在盛花期、灌浆期、乳熟期、蜡熟期以及完熟期随机选取10个生殖枝,手工摘除其小穗,于80℃烘至恒重,用万分之一天平称干重。
1.2.4种子产量 在完熟期随机选取10个生殖枝,手工采种,风干至种子含水量为8%左右后,用万分之一天平称重,即为单个生殖枝的种子产量。
1.3 数据分析
用SPSS 19.0统计软件对各指标进行方差分析,并对各指标与种子产量进行相关分析。
2 结果与分析
2.1 无芒隐子草种子发育过程中各位叶净光合速率
相同位叶在各时期的净光合速率有显著差异(P<0.05)(表1)。各位叶的净光合速率随种子发育总体呈下降趋势。其中,灌浆期第1~6叶显著高于其他时期,而第7叶的最大净光合速率出现在盛花期。除第7叶的净光合速率随种子发育一直呈显著下降外,其余各位叶在盛花期至乳熟期变异不大,而自乳熟期以后则随种子发育显著降低。同一时期不同位叶的净光合速率同样存在显著差异(P<0.05)(表1)。在相同的种子发育期内,同一生殖枝上的各位叶自上而下总体呈先增加后降低的趋势。在盛花期,第2~5叶显著高于其他位置叶片;灌浆期第3~4叶显著高于其他叶片,第1,6叶显著高于第7叶;乳熟期2~5叶相对较高,随后随生殖枝部位降低呈现显著下降;蜡熟期与完熟期第2~3叶净光合速率显著高于其他位叶,其后亦随生殖枝部位降低而下降。在各时期,第1,6,7叶净光合速率均显著小于其他位叶。
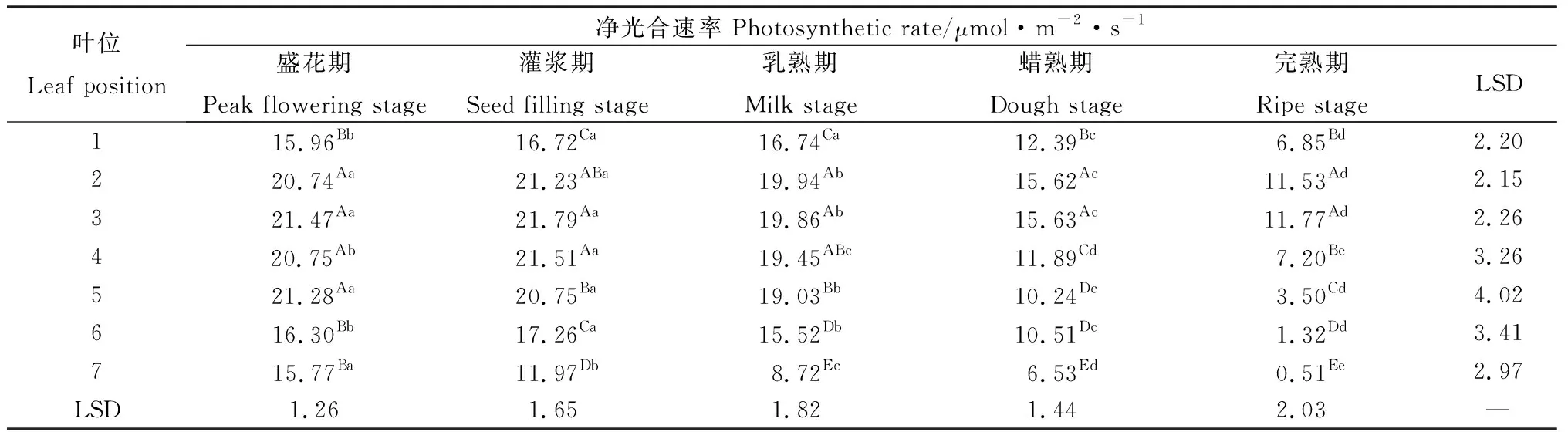
表1 无芒隐子草种子发育过程中各位叶净光合速率Table 1 Photosynthetic rate of C. songorica leaves in different position during seed development
注:同列不同大写字母表示不同位叶间差异显著(P<0.05),同行不同小写字母表示不同生育期间差异显著(P<0.05),下同
Note:Different capital letters in the same column indicate significant difference between positions at the 0.05 level, different lowercase letters in the same row indicate significant difference between stages at the 0.05 level. The same below
2.2 无芒隐子草种子发育过程中各位叶光合贡献率
相同位置叶片随着种子发育其光合贡献率表现出较大的波动(表2)。第1~2叶随种子发育始终呈现显著的上升趋势(P<0.05),而第3~4叶在乳熟期之前光合贡献率变化不大,之后则显著上升。在完熟期,第1~4叶的净光合速率占据了整个生殖枝叶片净光合速率的87.55%。第5~6叶的光合贡献率在乳熟期前差异不明显,随后则表现出显著的降低趋势。第7叶除在蜡熟期有所增加外,均随种子发育而显著下降。在同一时期,各位置叶片的净光合速率亦有显著差异(P<0.05)(表2)。同一生殖枝上的各位叶自上而下表现出先增加后降低的规律。盛花期第3叶与第5叶显著高于其他叶片;在灌浆期与乳熟期,第2~5叶显著高于其他位叶;蜡熟期与完熟期第2~3叶的光合贡献率占有绝对优势,其余叶片则随位置下降而显著降低。
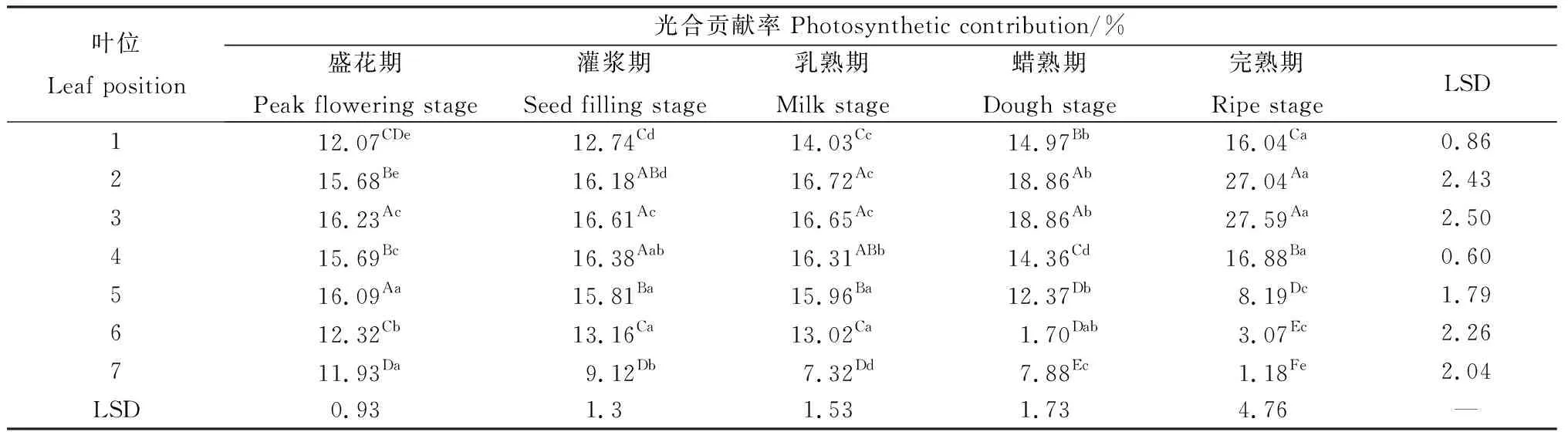
表2 无芒隐子草种子发育过程中各位叶光合贡献率Table 2 Photosynthetic contribution rate of C. songorica leaves in different position during seed development
2.3 无芒隐子草种子发育过程中各位叶重量
无芒隐子草生殖枝上相同位叶的重量在各发育期差异显著(P<0.05)(表3)。其中,第1~2叶在种子发育过程中表现出先增加后降低的趋势,自盛花期至灌浆期显著增加,而后则显著降低。第3~7叶始终随着种子发育呈现出降低的态势,其中,第3~5叶从盛花期至蜡熟期显著降低。第6~7叶则从盛花期至乳熟期显著降低,此后无显著变化。在同一时期内,不同位叶重量同样有显著差异(P<0.05)(表3)。在各时期,各位叶自上而下表现出先增加后降低的趋势,第4叶的重量在各时期均显著高于其他位叶,各个时期第5至第7叶均连续显著降低。多重比较表明,盛花期、灌浆期、乳熟期及完熟期1,2叶之间的重量无显著差异,蜡熟期2,3叶间无显著差异。
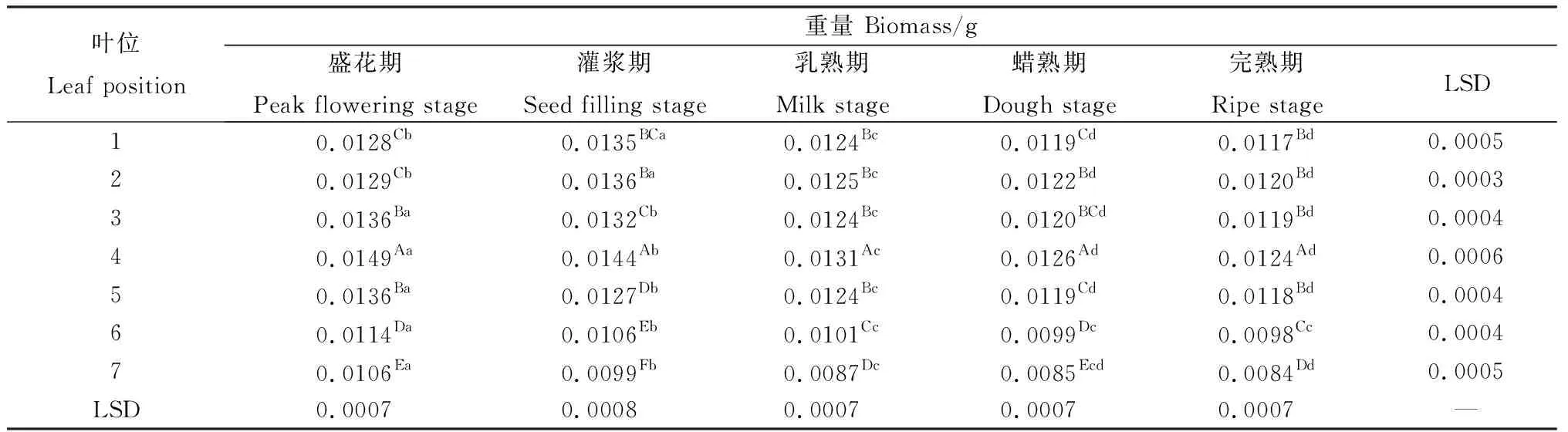
表3 无芒隐子草种子发育过程中各位叶重量Table 3 Biomass of C. songorica leaves in different position during seed development
2.4 无芒隐子草种子发育过程中各位叶重量比例
相同位叶在各个生育期的叶重量比例存在显著差异(P<0.05)(表4)。前3枚叶片随种子发育有上升趋势,第1~2叶盛花期叶重量比例显著小于其他时期;第3叶在整个种子发育期略有上升,但是差异不明显。第4,7叶在盛花期至乳熟期显著降低,之后无显著变化;第5~6叶除盛花期至灌浆期外,在各个时期差异均不显著。在同一生育期内,各位叶的叶重量比例变化显著(P<0.05)(表4)。各时期第4位叶的叶重量比例显著高于其他位叶,同时第6~7叶显著低于其他位叶,表明中部叶片构成了无芒隐子草生殖枝叶片重量的主要部分。
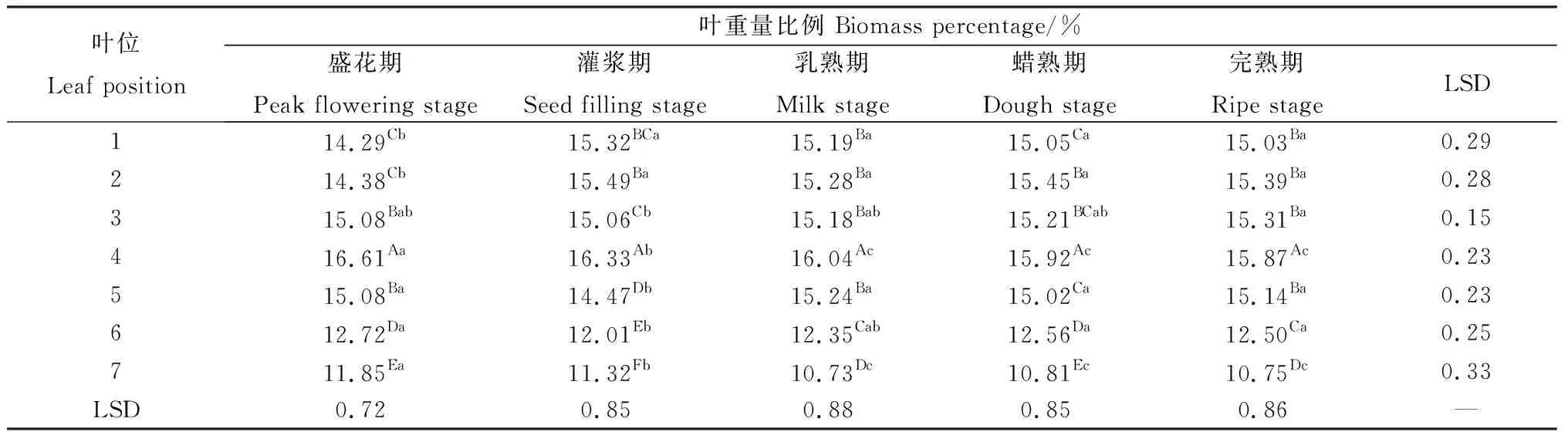
表4 无芒隐子草种子发育过程中各位叶重量比例Table 4 Biomass percentage of C. songorica leaves in different position during seed development
2.5 无芒隐子草种子发育过程中各位叶含水量
同一位叶在各个生育期的叶片含水量呈现出类似规律(表5)。第1~4叶以及第7叶在盛花期与灌浆期叶片含水量差异不显著,在灌浆期以后则持续显著降低(P<0.05)。第5叶在整个种子发育期内含水量均显著降低,第6叶的最大含水量出现在灌浆期,自灌浆期以后则显著降低。同一时期不同位叶的叶片含水量表现出自上而下逐渐降低的趋势(表5),各时期第1叶含水量均显著高于其他位叶,而第6~7叶含水量均显著低于其他位置叶片,其他叶片之间普遍表现出随位置降低而显著下降的趋势,其中盛花期与灌浆期2~3叶间含水量无显著差异,蜡熟期3~4叶间含水量无显著差异。
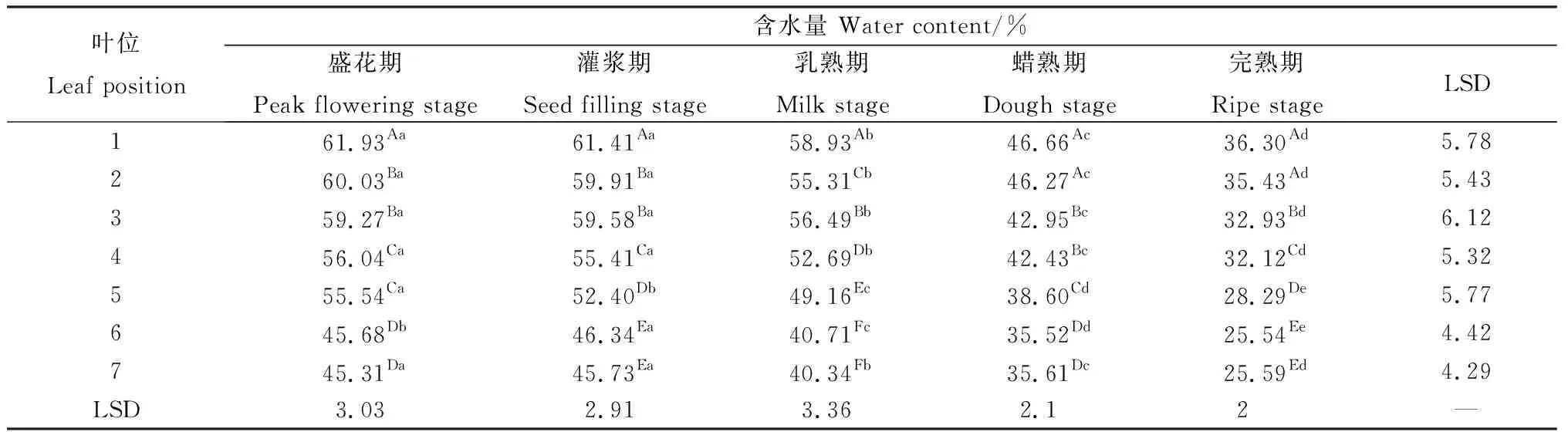
表5 无芒隐子草种子发育过程中各位叶含水量Table 5 Water content of C. songorica leaves in different position during seed development
2.6 叶片位置与时期对光合特性及生物量影响的双因素方差分析
通过双因子方差分析,可以看出叶位、种子发育期及其交互作用对净光合速率、叶生物量以及含水量均有极显著影响。种子发育期对光合贡献率及生物量比例无显著影响。叶位及种子发育期的交互作用对各个指标均有极显著影响 (表6)。
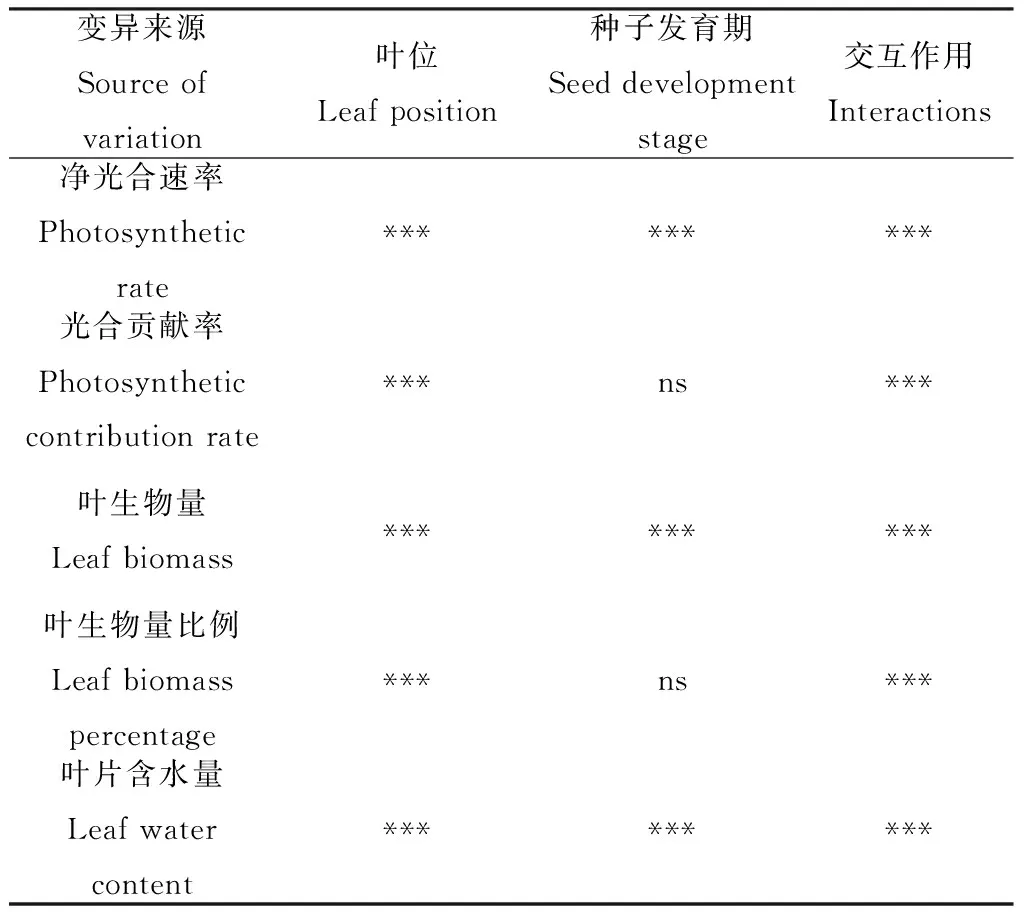
表6 叶位与种子发育期的双因素方差分析Table 6 Two-way ANOVA of leaf position, seed development stage and their interactions
注:***表示在0.001水平显著;ns:不显著
Note:***indicate significant at the 0.001 level;ns:Not significant at the 0.05 level
2.7 无芒隐子草种子发育过程中叶总生物量与穗生物量变化
随着无芒隐子草种子的发育,生殖枝上的叶总重量逐渐降低,其中盛花期与灌浆期间无显著差异,灌浆期与乳熟期间无显著差异,其余时期间则差异显著(P<0.05)。穗重量则随着种子发育持续显著增加(P<0.05)(表7),完熟期穗重量较盛花期增加了161%。
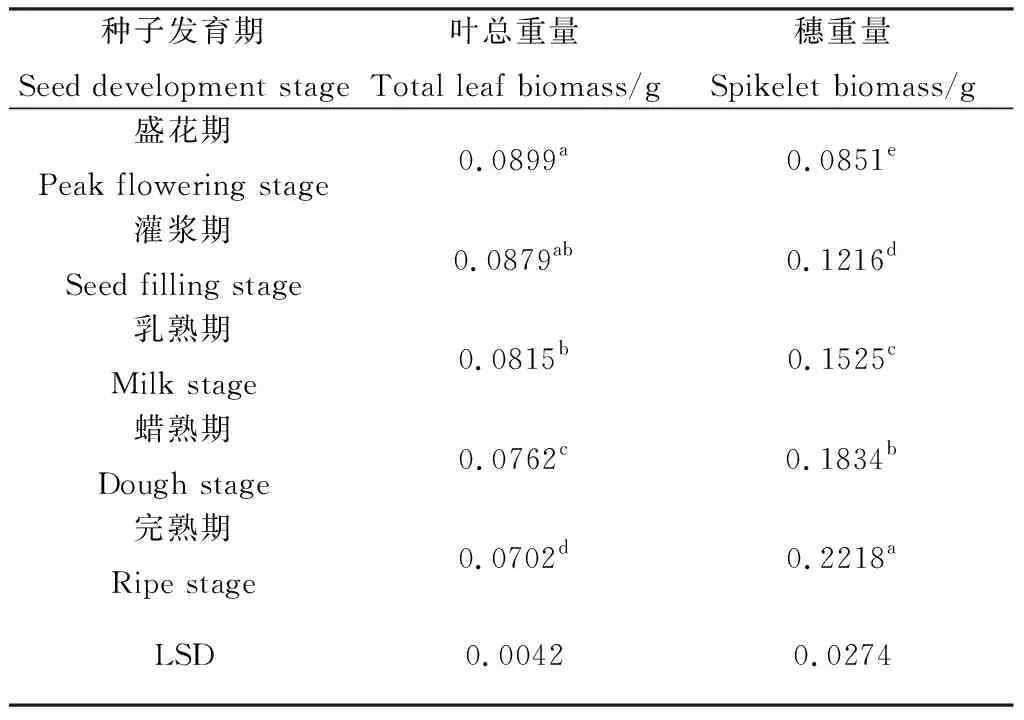
表7 无芒隐子草种子发育过程中叶总重量与穗重量Table 7 Total leaf biomass and spikelet biomass of C. songorica during seed development
2.8 无芒隐子草生殖枝种子产量与不同时期各位叶净光合速率、重量、穗生物量相关分析
各个位叶光合、形态参数与种子产量的相关性如表8所示。盛花期第1~6叶的净光合速率与重量均与种子产量呈正相关,其中第1,2,4叶的净光合速率,第1,3叶的重量与种子产量存在极显著正相关(P<0.01),第3,5叶净光合速率与第2,4,5,6叶重量与种子产量存在显著正相关(P<0.05),盛花期叶总重量与种子产量亦表现出极显著正相关关系,穗总重量则与种子产量呈现负相关。灌浆期第1~5叶的净光合速率、重量、叶总重量以及穗重量与种子产量之间表现出正相关,然而第6~7叶的净光合速率及重量与种子产量存在负相关关系,尤其是第7叶的净光合速率及重量分别与种子产量存在显著负相关与极显著负相关。乳熟期第1~6叶净光合速率及第1~5叶重量与种子产量呈正相关,其他位叶的净光合速率与重量则与种子产量呈负相关,乳熟期叶总重量与种子产量存在负相关关系,而穗重量则与种子产量呈显著正相关。蜡熟期2~7叶的净光合速率与种子产量存在负相关关系,第2,5,7叶重量亦与种子产量呈负相关,叶总重量与种子产量呈现显著负相关,穗重量则与种子产量呈现极显著正相关。完熟期第1~7叶净光合速率、第2~4与第6~7叶重量与种子产量均呈负相关,尤其以第3,7叶净光合速率与种子产量表现出极显著负相关,叶总重量与种子产量表现出显著负相关,穗重量则与种子产量呈极显著正相关。
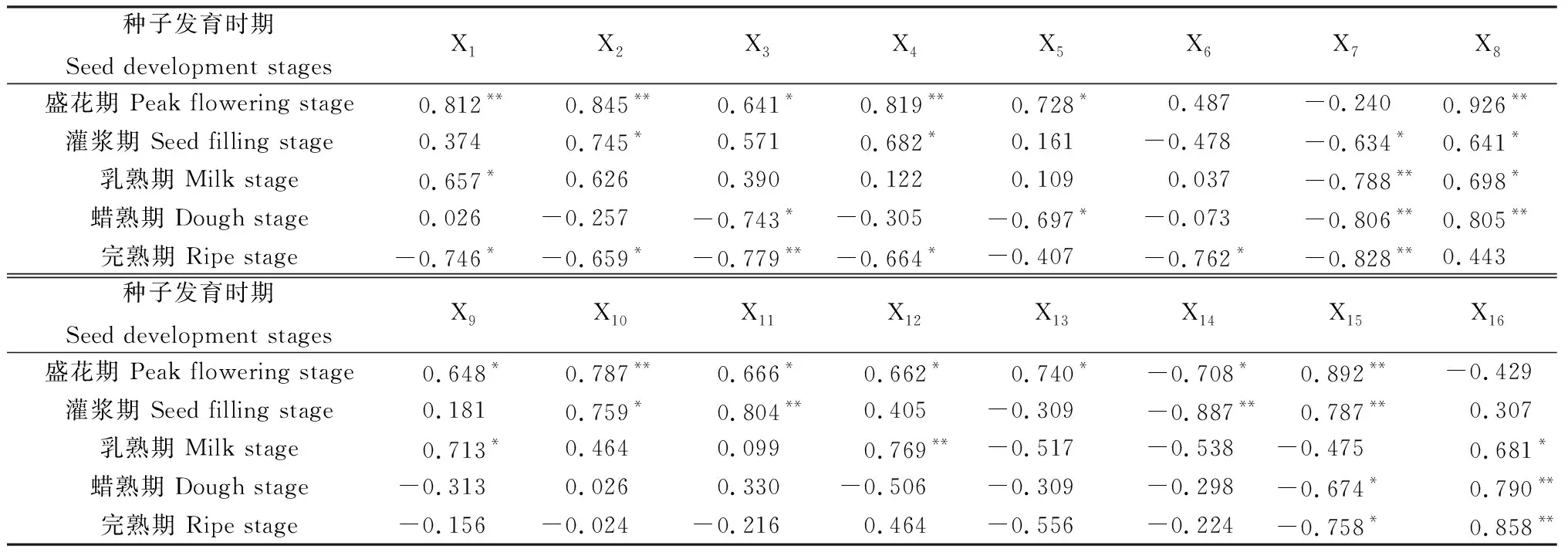
表8 无芒隐子草种子发育过程中各位叶光合速率及重量与种子产量相关分析Table 8 Correlation analysis between seed yield and photosynthetic rate,biomass of C. songorica leaves in different position during seed development
注:X1~X7为第1~7叶净光合速率,X8~X14为第1~7叶重量,X15为叶总重量,X16为穗重量,*表示显著相关(P<0.05),**表示极显著相关(P<0.01)
Note:X1~X7indicates photosynthetic rate from the first leaf to the seventh leaf, X8~X14indicates biomass from the first leaf to the seventh leaf, X15indicates total leaf biomass, X16indicates spikelet biomass, * indicates significant correlation at the 0.05 level, ** indicates extremely significant correlation at the 0.01 level
3 讨论与结论
3.1 无芒隐子草种子发育过程中各位叶净光合速率与光合贡献率变化
光合作用是产量形成的原动力,本研究说明在不同生育期,各位叶净光合速率与光合贡献率均有显著差异,并且同一位叶在各个时期的净光合速率与光合贡献率也有明显变化。在各时期,第2~5叶的光合贡献率始终占据绝对优势。李伟[17]等研究认为黄瓜(Cucumissativus)幼苗第3~4净光合速率最大,而第1,5叶较低。玉米(Zeamays)抽雄后不同位叶光合强度由高到低依次为中部>上部>下部[18],与本试验结果一致。类似的结果亦可见前人在向日葵(Helianthusannuus)[19]、生姜(Zingiberofficinale)[20]以及砂仁(Amomumvillosum)[21]等作物上的研究。韩清芳[22]等研究了紫花苜蓿(Medicagosativa)不同位叶净光合速率日变化,指出其日平均值依次为上位叶>中位叶>下位叶。水稻(Oryzasativa)抽穗期自上而下的4枚叶片净光合速率亦呈递减趋势[23]。各位叶的净光合速率主要受到叶片自身因素以及环境因子两方面影响。玉米上位叶叶绿体基质含量显著高于下位叶[24],且叶绿素含量亦随着叶位上升而增加[25],这被认为是造成不同位叶间光合特性差异的主要内部因素。从外部环境因子考虑,净光合速率主要受光合有效辐射(PAR)的影响[26],上位叶处在较好的光照与通风环境中,因此净光合速率普遍大于下位叶。本文着重探讨无芒隐子草不同位叶光合特性与种子产量的相关性,因此仅测定了种子发育过程中各位叶净光合速率,关于不同位叶的叶绿体超微结构、叶绿素含量以及生理生态因子的影响等则有待于进一步研究。
3.2 无芒隐子草种子发育过程中各位叶重量、重量比例、含水量以及生殖枝穗重量变化
任何作物的生长都可分为营养生长与生殖生长两个阶段,两阶段是对立统一的。营养生长是生殖生长的基础,其积累和储藏的物质,在生殖生长阶段流向植物生殖器官,促进产量形成;另一方面,过度的营养生长则会抑制生殖生长,造成减产。小麦(Triticumaestivum)、水稻等作物上的研究表明,种子产量最终取决于源、库容和物质运输能力[27]。在本试验中,无芒隐子草各位叶随着种子发育,重量均显著降低,其中第1~2叶最大重量出现于灌浆期,其他叶片最大重量出现在盛花期,同时,叶总重量的最大值出现在盛花期及灌浆期,这表明在盛花期以及灌浆期之后,叶片作为源器官,将其储存物质输送给了种子以及穗等库器官,在本试验中,生殖枝上穗的重量随着种子发育不断增加,亦印证了该观点。在蜡熟期至完熟期这一个阶段,各位叶重量虽略微下降,但没有显著差异,表明叶片对种子产量的贡献主要发生在盛花期至乳熟期这一阶段,自乳熟期以后,种子获得的储存物质或许主要由小穗提供,分析发现自乳熟期后小穗重量与种子产量始终呈显著正相关,也在一定程度上证明了该结论。
3.3 无芒隐子草种子发育过程中各位叶净光合速率、重量变化与种子产量相关性
不同位叶在种子形成过程中起到的作用是不同的。盛花期前6位叶的净光合速率和重量均与种子产量呈正相关,在灌浆期与乳熟期,前5叶的净光合速率和重量亦与种子产量正相关,而在蜡熟期,仅有第1叶的净光合速率、第1, 3, 4叶重量与种子产量呈正相关,至完熟期,则仅有第1叶重量与种子产量呈正相关。在围绕光合特性与作物产量的研究中,不同学者在不同作物上的研究结果并不一致,玉米上位叶光合速率和贡献率显著高于下位叶,从而对成粒率、籽粒充实度、千粒重和产量起决定性作用[25]。然而亦有研究表明光合速率与产量相关性很小[28],收获指数增加与光合周期增长才是产量提高的主因。在本试验中,前5枚叶片的净光合速率与重量在乳熟期以前始终对种子产量起到积极作用,而第6位叶净光合速率与重量仅仅在盛花期与种子产量表现出正相关,第7位叶在种子发育过程中则始终与种子产量负相关。因此,上位叶即1~5位的叶片是与种子产量密切相关的,尤其是在盛花期至乳熟期这一阶段,此结论与于洪兰[29]等人在水稻上的研究结果一致。在河西走廊地区,无芒隐子草盛花期至乳熟期,对应7月下旬至8月下旬这一时间段[9],正是该地区褐斑病(Rhizoctoniasolani)、白粉病(Blumeriagraminis)等田间病害最为盛行的时期[30],在该时段,采取适宜的田间管理措施控制病虫害,保证叶片健康完整是种子高产的基础。同时,采取密度调控、施肥、灌溉等管理措施,设法增大叶片光合与形态参数,也会对种子产量产生积极影响。
