川西北高寒区不同沙化类型草地优势种群空间格局分析
2017-09-13邓东周鄢武先陈德朝
邓东周, 贺 丽, 鄢武先, 陈德朝
(四川省林业科学研究院, 四川 成都 610081)
植物种群分布格局是研究种群特征、种群间相互作用以及种群与环境关系的重要手段,用以说明种群在空间的配置状况或分布特点,也是空间水平上种群个体的彼此相互关系的反映,是现代生态学研究的主题之一[1-3]。优势种群的生态生物学特征及其发生发展的动态,对群落的外貌、结构、演替方向都有至关重要的影响[4]。通过研究分析优势种群分布格局,不仅可以了解群落内部物种的分布状况,而且有助于掌握种间相互作用规律以及种群分布与环境之间存在的因果关系。深入研究种群空间分布格局及动态,可为密度控制、群落演替和草地资源的合理利用提供理论依据。
川西北高寒沙地类型区主要指高寒地区草地退化形成的沙化土地,该类型区位于四川西北部,地处青藏高原东南缘,属我国长江、黄河源头,是《全国主体功能区规划》确定的国家重点生态功能区之一,生态区位重要,是长江、黄河流域重要的生态安全屏障。区内森林、草原和湿地等生态系统,在水源涵养、水资源补给、水土保持、生物多样性保护、区域气候调节等方面起着重要作用。目前对川西北沙化土地的研究主要集中于沙化成因[5-6],沙地治理技术[7],不同沙化类型植被特征等方面的研究[8-10],而对于该区域不同沙化类型优势植物种群空间分布格局的研究报道甚少。本文采用方差均值比率法、聚集强度等方法研究了川西北高寒沙区植物群落优势种群空间分布格局,旨在揭示影响各沙化类型优势种群空间分布格局主要因素,从而为川西北高寒沙化地植被恢复提供科学依据。
1 研究区概况与方法
1.1 研究区概况
川西北高寒区地处青藏高原东南缘,四川省的西北部,包括甘孜藏族自治州、阿坝藏族羌族自治州的28个县,海拔多在3 000 m以上。地理坐标为E 97°21′~104°24′,N 28°16′~34°19′。气候特点:属高原高寒地区,气候严寒,冬长夏短,春秋短,寒冷干燥,日照充足,昼夜温差大,年冻土时间长达6个月。根据若尔盖、红原等县的气象资料,年平均气温在-2.5℃~6℃,年降水量500~900 mm,且集中在5-9月,占年降水量的70%以上。研究区域植被以高山草甸为主。
1.2 样方设置与调查
根据川西北高寒区沙化土地的分布特征,本研究确定选择若尔盖县、阿坝县、理塘县、稻城县、色达县5个典型沙化县为研究区,结合5个县的沙化情况设置调查样线,样线需沿海拔梯度尽可能囊括所有的沙化类型。沿样线每隔100 m设置样方,根据四川省第四次沙化监测数据,若尔盖、理塘2县分布有4种类型的沙化草地,样方数为8*4*2=64个;稻城、阿坝、色达3县分布有露沙地、固定沙地、半固定沙地等3种类型的沙化草地,样方数为8*3*3=72个,共计136个1 m*1 m的样方,按每个县、每种沙化类型均有样方的原则选取4~8个优势群落样方,记录植物的名称、盖度、高度和株数等数据,然后进行物种多度等数据计算。
1.3 数据分析
多度计算方法:多度=某物种株数/样方内各种植物的总株数。
为了避免不同指数的片面性,测定生物种群空间分布格局类型采用的指数有扩散系数 (ID),聚集度大小指数(ICS),Green指数(GI),各指数计算公式如下:
(1)扩散系数 (ID)(方差均值比):
当ID< 1时为均匀分布;ID= 1时为随机分布;ID> 1时为聚集分布。
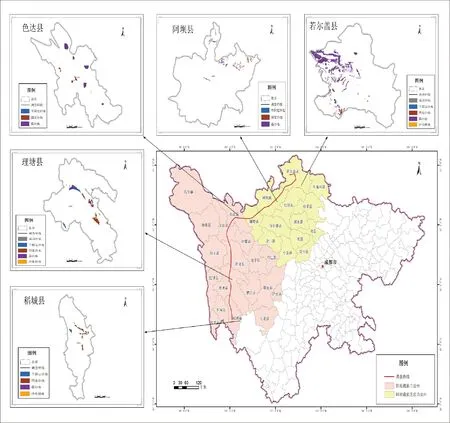
图1 研究区域样线的布设
Fig.1 Line transect layout of study area
格局强度是种群空间分布格局的一个重要指标,采用聚集频度指数(ICF),平均拥挤度指数(IMC),缀块指数(IP)等指数来表征聚集强度。各指数计算公式如下:
2 结果与分析
2.1 露沙地优势种群空间分布格局
在露沙上分布的植物群落种类较少,常见的植物群落有高山嵩草(Kobresiapygmaea)、四川嵩草(K.setchwanensis)、粗壮嵩草(K.robusta)、线叶嵩草(K.capillifoli)等。各群落中优势种群为高山嵩草、四川嵩草、粗壮嵩草、线叶嵩草等4个种群。
4个优势种群的空间分布格局测定结果如表1所示,根据ID、ICS、GI值显示,各优势种群在空间上呈现聚集分布,但聚集程度不同。聚集强度指数计算结果显示,聚集强度大小表现为高山嵩草种群>四川嵩草种群>粗壮嵩草种群>线叶嵩草种群。在该沙化类型中优势种群的ICF(K)值均较小,表明在该区域出现的频率较小;IMC值和IP值均较大,表明平均在同一样方内每个嵩草个体的邻居数多,受到的拥挤效应强,聚集分布强。这主要是由于嵩草类植物生活型和繁殖方式相同,以丛生为主,发展迅速,嵩草群落中其他植物分布较少,易形成独立的优势种群,
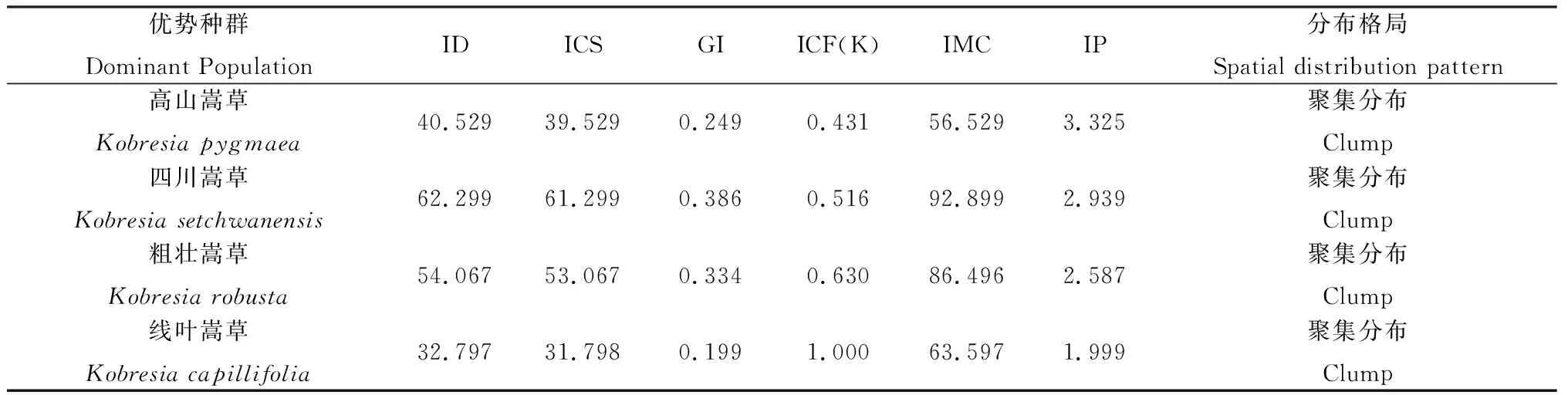
表1 露沙地优势种群空间分布格局测定结果Table 1 The spatial distribution pattern of dominant population in exposure sandy land
2.2 固定沙地优势种群空间分布格局
在固定沙地上分布的植物群落种类较多,常见的植物群落有细叶亚菊(Ajaniatenuifolia)、鹅绒委陵菜(Potentillaanserina)、楔叶委陵菜(Potentillacuneata)、黄帚橐吾(Ligulariavirgaurea)、细叶西伯利亚蓼(Polygonumsibiricumvar.thomsonii) 等。各群落中优势种群为细叶亚菊、鹅绒委陵菜、楔叶委陵菜、黄帚橐吾、细叶西伯利亚蓼等6个种群。在理塘县分布有较多的马蹄黄群落,马蹄黄为优势种群。
从表2可知,各优势种群均呈聚集分布。ICF(K)、IMC、IP计算结果显示,3种指数大小均可以说明鹅绒委陵菜种群在该沙化类型聚集分布强度最大,细叶亚菊和马蹄黄的种群聚集分布强度相对较低。这与鹅绒委陵菜主要以其发达的匍匐茎来进行无性繁殖,发展迅速,耐践踏,易形成独立的优势种群有关。细叶亚菊种群ICF指数最高,主要是因为细叶亚菊是多年生草本植物,主要依靠种子繁殖后代,种子借助风力传播范围较广,且其种子在此生境中的萌发率较高,出现在区段的频率较高,每个个体的邻居数少,受到的拥挤效应较小,即IMC值较小。
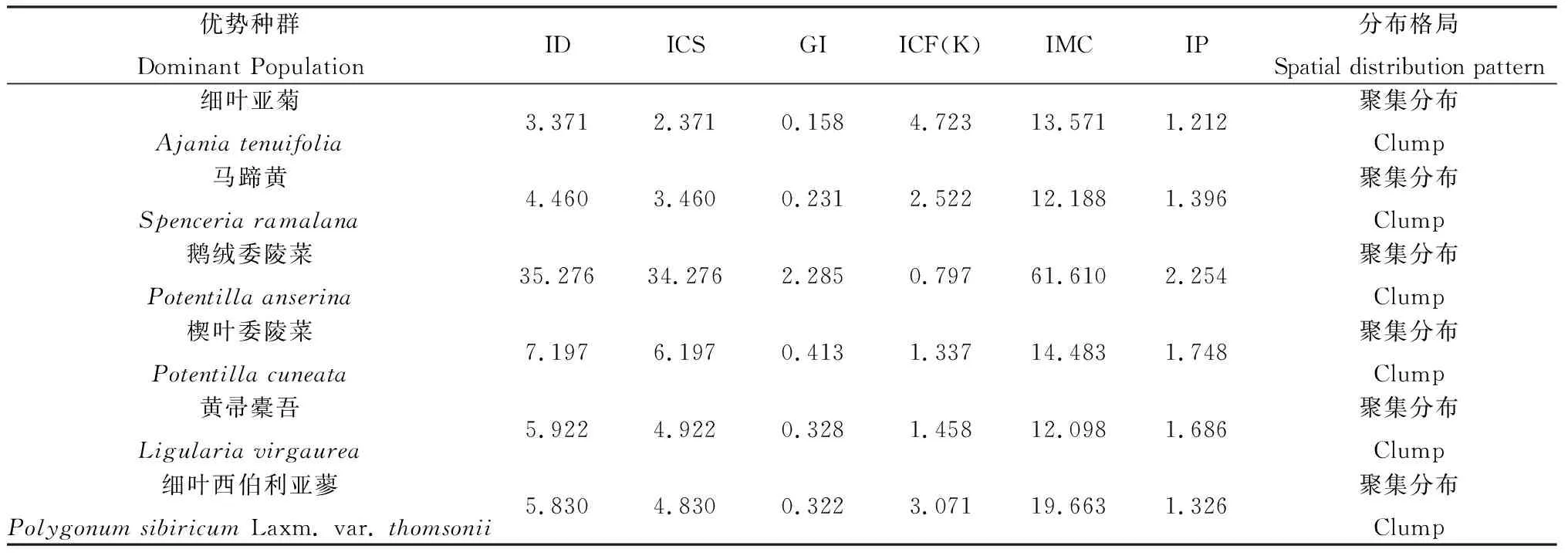
表2 固定沙地优势种群空间分布格局测定结果Table 2 The spatial distribution pattern of dominant population in fixed sandy land
2.3 半固定沙地优势种群空间分布格局
半固定沙地上常见的植物群落有白花枝子花(Dracocephalumheterophyllum)、露蕊乌头(Aconitumgymnandrum)、楔叶委陵菜(Potentillacuneata)、细叶西伯利亚蓼等。各群落中优势种群为白花枝子花、露蕊乌头、楔叶委陵菜、细叶西伯利亚蓼等4个种群。在理塘县还分布有较多的舟叶橐吾群落。
4个优势种群的空间分布格局如表3所示,根据ID、ICS、GI值大小可以判定半固定沙地优势群落空间分布格局为聚集分布。ICF(K)、IMC、IP值表明楔叶委陵菜种群聚集程度大于其他优势种群,其余几种优势聚集强度差异较小,这一方面与楔叶委陵菜以发达的匍匐茎来进行无性繁殖有关,其余几种植物主要是靠种子繁殖,借助风力传播范围较广,所以出现的频率高,即ICF(K)值较高,每个个体的邻居数少,受到拥挤效应较小;另一方面楔叶委陵菜耐贫瘠,能在土壤贫瘠的半固定沙地成较强的聚集分布,对于其余几种优势种群而言,对土壤条件要求稍高,聚集程度较低。
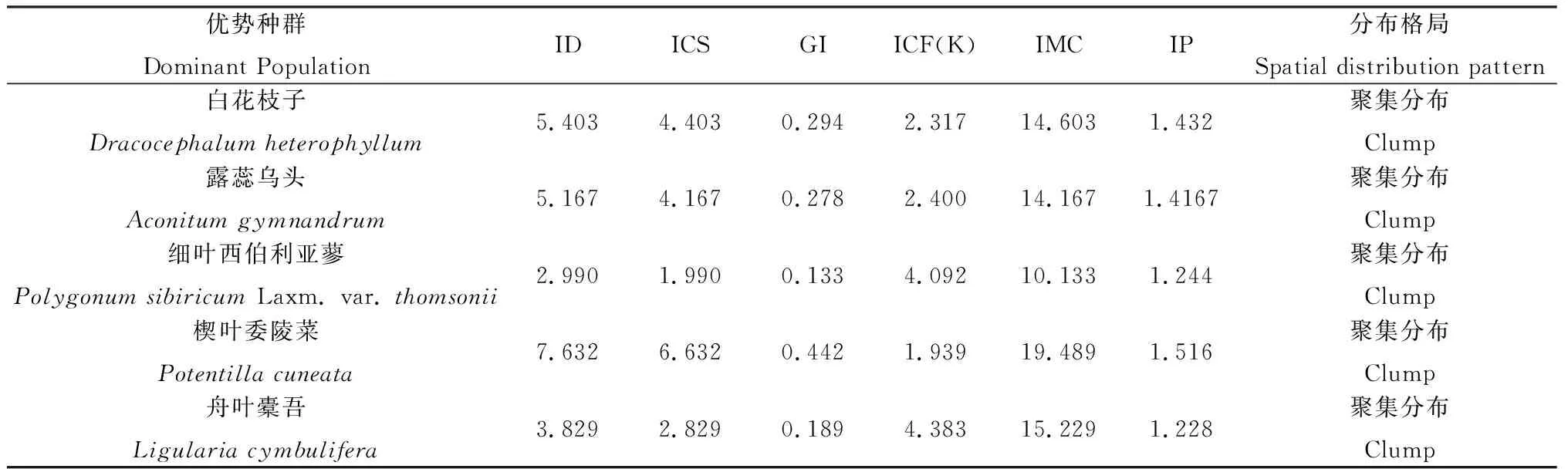
表3 半固定沙地优势种群空间分布格局测定结果Table 3 The spatial distribution pattern of dominant population in semi fixed dune
2.4 流动沙地优势种群空间分布格局
在流动沙地上常见的植物群落极少,主要有青藏苔草(Carexmoorcroftii)、赖草(Leymussecalinus)、异株矮麻黄(EphedraminutaFlorin var.dioeca)等群落。各群落中优势种群为青藏苔草、赖草、异株矮麻黄等3个种群,其他植物分布极少。
3个优势种群的空间分布格局如表4所示,根据ID、ICS、GI值大小可知各种群均呈现聚集分布。聚集强度指数ICF(K)、IMC、IP分析结果显示,3种优势种群聚集强度差异不明显,说明流动沙地土壤微生境对优势种群空间分布格局的影响强于物种的生物学特性。因为流动沙地土壤极为贫瘠,只有极少耐旱、耐贫瘠的植物才能存活。

表4 流动沙地优势种群空间分布格局测定结果Table 4 The spatial distribution pattern of dominant population in shifting sandy land
2.5 聚集指数相关关系分析
从表5可以看出,ID,ICS,GI,IMC之间呈显著的正相关且相关系数几乎等于1,说明他们表征相同的生态学意义。ICF与ID,ICS,GI,IMC,IP之间呈负相关关系,说明他们表征的生态学意义相反。但他们的分布格局都是聚集分布。
注:*表示显著相关P<0.05;**表示极显著相关P<0.01
Note: *and**indicate significant correlation at the 0.05 and 0.01 level, respectively
3 讨论
种群个体在水平空间上的分布格局,不仅与种群本身的生物学特性有关,同时也受生境条件或种群间效应的直接影响[17-20]。通过对优势种群分布格局的测定,可以揭示群落空间结构的特征,有助于了解这些种群的某些生物生态学特性及一定环境因子对种群行为或生存的作用。川西北高寒区不同沙化类型草地优势种群呈聚集分布,且露沙地优势种群聚集强度大于其余3种沙化类型,聚集程度强,使种群在某一斑块上形成优势,抵抗外来种的侵入和定居,从而维持种群的稳定和续存。这主要与嵩草植物的生物学特性和土壤微生境的影响密切相关。就植物而言,由于露沙地优势种群为嵩草种群,植物个体矮小,对于个体矮小的草本植物而言,其种子散布的距离和范围更小,且种子小而数量大,更容易形成格局规模较小的聚集分布,聚集强度大[21];就沙化类型而言,露沙地土壤养分和水分资源较为丰富,植物可以充分利用土壤养分资源,限制其他植物种生长。这与大多数自然种群的稳定分布格局为聚集分布结果一致[22]。露沙地、固定沙地同一沙化类型优势种群聚集强度差异较大,主要与其生物学特性相关。半固定沙地、流动沙地同一沙化类型优势种群聚集强度差异较小,这两类沙化类型的土壤极为贫瘠,地下资源不能满足植物大面积繁殖的需要,土壤环境对其聚集分布的影响占主导作用。同一优势种群在固定沙地上聚集强度大于半固定沙地,如细叶西伯利亚蓼种群、楔叶委陵菜种群,这主要是由细叶西伯利亚蓼种群的发生和发展特性以及具体的生境条件所决定的,体现了种群对不同沙化类型的生存策略或进化适应机制,这与张炜银等[23]对薇甘菊种群在不同生境中的分布格局研究结果一致。
不同聚集强度指标的测度, 并不是度量同一种群聚集程度的不同方法, 而是从不同的角度来度量同一种群的聚集特性。在种群空间分布格局研究中,频数分布的方法由于简单易行, 且有相应的计算机软件,因此应用最为广泛。对于聚集度大小指数(ICS),Green指数(GI),聚集频度指数(ICF),平均拥挤度指数(IMC),缀块指数(IP)各项指数而言,均是由扩散系数(ID)衍生而成的,他们在本质上是相同的或具有较大的相似性,从表5可看出,ID,ICS,GI,IMC之间的相关系数几乎等于1,IP与ID,ICS,GI,IMC的相关系数相对较低。这与徐连秀等[24]人的研究结果相似。对各沙化类型优势种群分布格局类型和聚集强度指数计算的结果进行排序,其结果总体一致。由于聚集频度指数(ICF)相当于负二项分布中的K值,与种群密度没有关系,在种群总体数量因随机死亡而减少时,K保持不变,而在同一样方内每个个体的邻居数会发生变化,IMC、IP值会发生变化,因此,用K作为度量聚集程度的指数是适宜的,因此本文中对优势种群聚集程度的比较均以聚集频度指数(ICF)大小为主。多数种群空间格局强度分析研究均采用ICF(K)值作为聚集程度判断的指数[21, 24-28]。
4 结论
本文定量研究了川西北高寒区若尔盖县、阿坝县、理塘县、稻城县、色达县5个典型沙化县不同沙化类型草地优势种群空间分布格局,研究结果表明,影响优势种群空间分布格局的主要因素因沙化类型的不同而不同,露沙地和固定沙地影响优势种群空间分布格局的主要因素为物种生物学特性,半固定和流动沙地影响优势种群空间分布格局的主要因素为土壤微生境。因此,在川西北高寒区沙化土地治理过程中,可以根据影响种群空间分布格局的主要因素针对性的采取恢复措施,对于露沙地和固定沙地而言,建议采取播种牧草,增加物种的聚集程度,露沙地群落更加稳定,固定沙地逐渐向露沙地演替;对于半固定和流动沙地而言,建议采取改良土壤微生境与播种牧草多措施结合,固定流沙,促进植被恢复。
