黄土高原中西部刺槐人工林生态系统碳密度及其影响因子
2017-09-12牛春梅关晋宏程然然李国庆吴春荣程积民
牛春梅,关晋宏,程然然,李国庆,吴春荣,程积民,杜 盛,*
1 西北农林科技大学林学院,杨凌 712100 2 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100 3 中国科学院水利部水土保持研究所,杨凌 712100 4 甘肃省治沙研究所,兰州 730000
黄土高原中西部刺槐人工林生态系统碳密度及其影响因子
牛春梅1,2,关晋宏1,程然然2,3,李国庆2,3,吴春荣4,程积民2,3,杜 盛2,3,*
1 西北农林科技大学林学院,杨凌 712100 2 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100 3 中国科学院水利部水土保持研究所,杨凌 712100 4 甘肃省治沙研究所,兰州 730000
为阐明黄土高原中西部刺槐人工林碳密度区域分布特征及其主要影响因子,基于野外样地调查和室内样品分析估算了黄土高原中西部4个栽培区域的刺槐人工林生态系统碳密度及其分布特征,并利用相关性分析和主成分分析分析了影响生态系统碳密度的主要因子(林分、地形、土壤和气候等)。结果表明:调查区5个林龄的刺槐人工林生态系统生物量为34.13—133.08 t/hm2,不同区域之间各组分生物量存在显著性差异。植被层平均碳含量为221.93—454.67 g/kg,总体上表现为乔木层平均碳含量高于灌、草层,枯落物层平均碳含量最低,不同区域乔木、灌木、草本平均碳含量均存在显著性差异。刺槐人工林生态系统碳密度均值为106.86 t/hm2,其中土壤层碳密度占刺槐人工林生态系统总碳密度的64.09%,是刺槐人工林生态系统碳密度的主要组成部分。植被层碳密度为38.68 t/hm2,其中乔木层碳密度(33.88 t/hm2)占植被层碳密度的87.58%,灌木、草本、枯落物所占比例依次为1.98%(0.77 t/hm2)、2.00%(0.77 t/hm2)、8.43%(3.26 t/hm2)。不同区域土壤、生态系统碳密度均存在显著性差异。相关性分析和主成分分析表明,刺槐人工林生态系统碳密度与林龄、降水量呈显著正相关关系,与林分密度、平均气温、海拔和坡度的相关关系不显著,上述林分因子、地形因子和环境因子转化的主成分方差累积贡献率为91.07%,其中林龄和降水量是影响刺槐人工林生态系统碳密度的主要因子,方差贡献率为37.22%。
黄土高原中西部;刺槐人工林;碳密度;生物量
在全球气候变化背景下,大气CO2的吸收与排放已成为许多学科的研究热点之一。森林是陆地生态系统的主体,对大气CO2的吸收、固定和循环发挥着重要作用[1- 4]。近年来,随着退耕还林等生态工程的实施,人工林得到迅速发展,我国已成为世界人工林面积最大的国家[5],针对人工林碳汇功能的关注度也日益增加[6]。因此,对人工林生态系统碳固存及其影响因子的研究,对于合理评估人工林生态系统长期生产力以及开展有效的经营管理以提高其综合服务功能具有重要意义。
黄土高原中西部地区横跨干旱、半干旱和半湿润气候区,是我国生态环境脆弱区域之一,植被恢复多以保持水土和改善区域生态环境为主要目的。近年来,除了水土保持功能之外,半干旱地区人工植被特别是人工林生态系统的固碳功能也受到广泛关注[7-8]。关于黄土高原地区森林固碳的研究有部分学者进行过报道[9- 10],刘迎春等[11]对黄土丘陵区不同树种间碳储量及固碳潜力进行了比较,艾泽民等[12]研究了黄土丘陵区不同林龄刺槐人工林碳氮储量及分配格局,孟蕾等[13]、杨晓梅等[14]都对黄土高原子午岭区森林碳储量与碳密度进行了估算,但相关研究多是针对较小尺度的某一林区展开的,且对森林生态系统碳密度的分配特征方面的研究较多,而对较大尺度上生态系统碳密度空间变化特征及其与林分和环境因子间关系的研究相对较少。这在一定程度上制约了较大区域尺度上森林碳密度估算的准确性与可靠性。因此,需要开展更多基于区域尺度样地调查的精确参数来减小碳密度估算结果的不确定性。在研究内容上,本研究不仅对森林生态系统碳密度进行了估算,而且从地形、环境、林分等方面对影响森林生态系统碳密度的因素进行研究,以全面精确地估算黄土高原中西部森林生态系统碳储量。
刺槐(Robiniapseudoacacia)是黄土高原的主要造林树种,其人工林广泛分布于半湿润和半干旱地区,甚至在干旱地区也有栽培,在水土保持和生态防护方面起着重要作用[15]。研究刺槐人工林生态系统碳密度及其影响因子,有利于深入了解黄土高原人工林生态系统碳储存能力。本研究以黄土高原中西部4个栽培区域的刺槐人工林为研究对象,分析其碳密度特征及主要影响因子,为深入了解整个黄土高原地区人工林碳固存及人工林经营管理提供依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于甘肃省白银市靖远县、平凉市泾川县、庆阳市正宁县中湾林场和陕西省延安市公路山林地4个区域。靖远县位于甘肃省中部(36°—37°15′ N,104°13′—105°15′ E),属温带干旱半干旱气候,海拔1300—3017 m,年均温9°C左右,年降水量228.7 mm。泾川县位于甘肃省东部(34°59′—35°18′ N,107°38′—107°58′ E),属暖温带半干旱气候,海拔950—1225 m,年均温10.6°C,年降水量584.1 mm。中湾林场地处黄土高原中部子午岭区南段(35°17′ N,108°27′ E),属温带半湿润气候,海拔1246—1756 m,年均温9.2°C,年降水量609.7 mm。延安市公路山(36°25′ N,109°32′ E)属温带半干旱气候,海拔1245—1395 m,年均温10.3 ℃,年降水量516 mm。调查区内样地优势树种均为刺槐,林下有稀疏的草本植物,主要有大披针苔草(Carexlanceolata)、披碱草(Elymusdahuricus)、三褶脉紫菀(Asterageratoides)、广布野豌豆(Vicialilacina)、蛇莓(Duchesneaindica)、铁杆蒿(Artemisiasacrorum)等。
1.2 研究方法
1.2.1 调查样地设置
2011年和2012年,在刺槐人工林分布较广的4个区域内设置标准样地14块,其中白银2块、庆阳3块、延安3块、平凉6块,位于平凉的6块样地按林龄又分为平凉1(中龄林组,3块)和平凉2(成熟林组,3块)。原则上样地面积设置为20 m×50 m,但由于地形破碎等原因,部分样地面积设置为20 m×30 m。利用GPS确定每个样地的位置,并记录每个样地的海拔、坡度等地形因子以及林分密度等林分因子,利用生长锥法测定林分年龄,利用中国气象数据网网站获取各研究区域年均温(AMT)、年降水量(MAP)(1981—2010)等环境因子数据,所选样地基本信息情况见表1。
1.2.2 样地调查及植被层生物量测定
乔木生物量测定:对标准样地内DBH≥2 cm的乔木每木检尺,记录其胸径(cm)和树高(m),并按大中小径级选择3—5株样木,分叶、枝、干、根采集植物样品,将相同器官的样品混合,再取各器官混合样(不少于300 g)带回实验室。刺槐乔木生物量根据森林固碳课题办公室提供的异速生长方程[16]进行计算,由胸径和树高求得样地每株刺槐各器官生物量,各器官生物量之和为乔木单株总生物量,根据样地所有乔木单株生物量之和求得样地乔木总生物量(kg)并转换为单位面积生物量(t/hm2)。
式中,WS为乔木干生物量(kg);WB为乔木枝生物量(kg);WL为乔木叶生物量(kg);WR为乔木根生物量(kg);D为单株乔木胸径(cm);H为单株乔木高度(m)。
灌木、草本和枯落物生物量的测定:在每个样地内沿对角线设置3个2 m×2 m灌木样方,记录灌木种类后,将样方内所有灌木全部收获,分叶、枝和根称鲜重并取样。草本的调查方法与样方设置(1 m×1 m)与灌木相同,草本分地上和地下部分称鲜重并取样。枯落物调查则将草本样方内枯落物全部收获后称鲜重并取样。将采集的灌木、草本和枯落物样品带回实验室,置于85°C烘箱烘干至恒重,测定其含水率。根据各器官含水率换算为其干质量求得灌木、草本各器官及枯落物生物量。
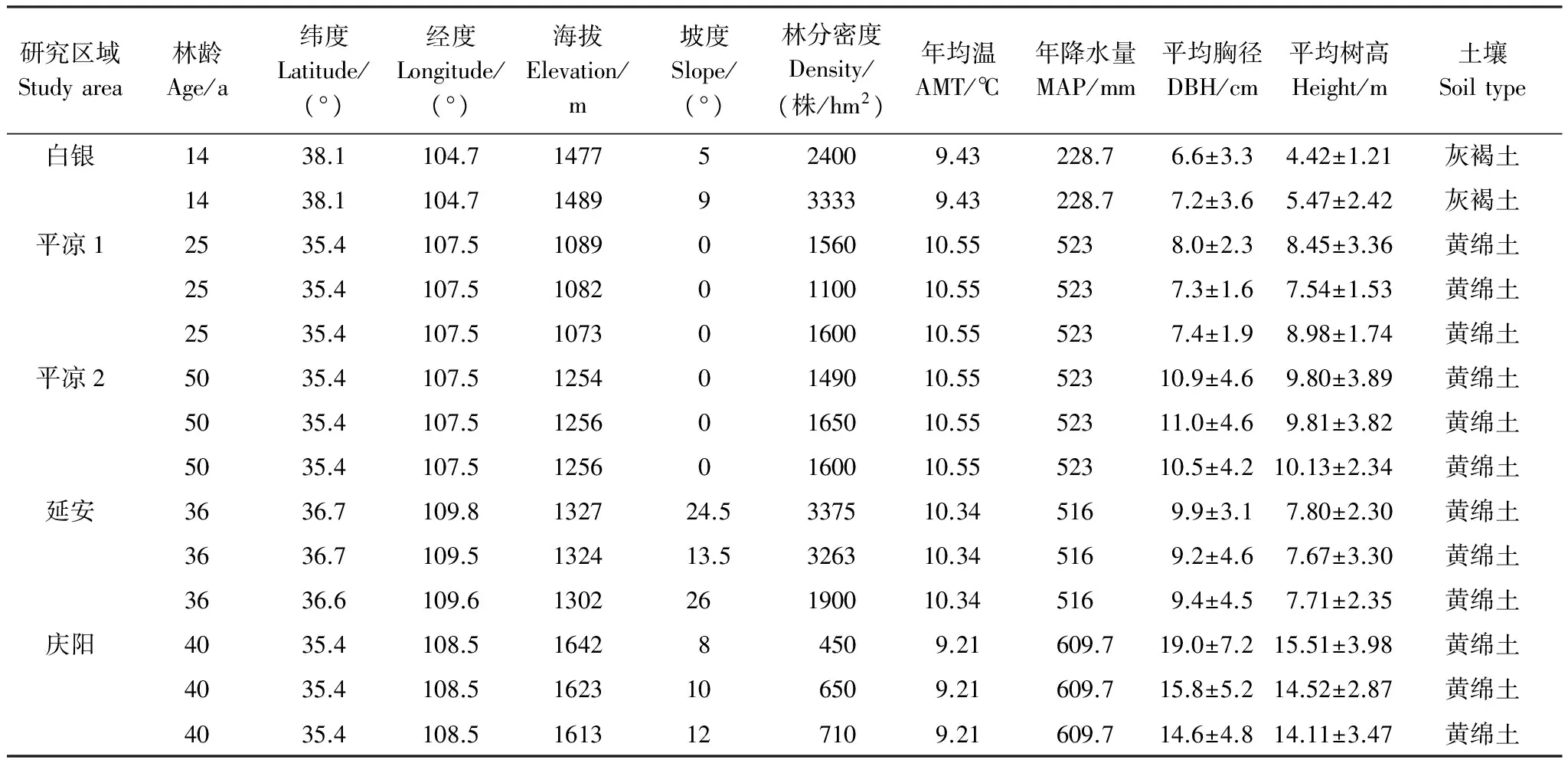
表1 样地基本概况
土壤调查采用剖面法:在调查样地内挖土壤剖面,深至100 cm。沿剖面按0—10 cm,10—20 cm,20—30 cm,30—50 cm,50—100 cm不同深度,用环刀取各个土层的原状土,带回实验室置于105°C烘箱烘干至恒重,计算各土层土壤容重。在样地的坡上、坡中、坡下,使用内径为6 cm的土钻,按照上述五个层次每层随机取3钻土,同一土层混合为一个样品后带回实验室,用于土壤有机碳含量测定。
1.2.3 碳含量测定
将带回实验室的植物样品烘干至恒重,磨碎过0.25 mm筛,装塑封袋保存待测。土壤样品带回室内阴凉处自然风干,研磨后过2 mm筛(将大于2 mm的砾石单独称重),再用四分法取部分样品研磨过0.25 mm筛,装袋标记后备测。植物样品和土壤样品碳含量测定均采用重铬酸钾-硫酸氧化外加热法。
1.2.4 碳密度计算
生态系统总碳密度为乔木、灌木、草本、枯落物和土壤各层碳密度之和。乔木层碳密度根据乔木各器官生物量乘以相应碳含量进行计算。灌木层碳密度根据灌木各器官生物量乘以对应碳含量进行计算。草本层碳密度根据地上和地下部分生物量乘以相应碳含量进行计算。枯落物层碳密度根据枯落物层生物量乘以其碳含量进行计算。土壤有机碳密度(SOCD,t/hm2)计算公式为:

式中,Ci为第i层土壤有机碳含量(g/kg),BDi为该层土壤平均容重(g/cm3),Di为该层土壤厚度(cm),Ri为直径>2 mm的石砾所占百分比(%),由于研究区域砾石含量较少,因此Ri为0。
1.2.5 平均碳含量计算
采用生物量加权平均法计算植被层各组分平均碳含量。计算乔木层各器官生物量比例,按各器官碳含量与其对应生物量比例相乘计算乔木层平均碳含量。计算灌木层各器官生物量比例,按各器官碳含量与其对应生物量比例相乘计算灌木层平均碳含量。根据草本层地上、地下部分碳含量与其对应生物量比例相乘求得草本层平均碳含量。
1.2.6 数据处理
利用Excel 2010和SPSS 18.0软件进行统计分析。采用单因素方差分析(one-way ANOVA)和最小显著差数法(LSD)进行差异显著性检验,采用Pearson法进行相关性分析,对影响生态系统碳密度的因子进行主成分分析。图表中数据为平均值±标准差。
2 结果与分析
2.1 不同区域刺槐人工林生态系统生物量
黄土高原中西部刺槐人工林总生物量在34.13—133.08 t/hm2之间,4个区域生物量之间存在显著差异(P<0.05)(表2),这可能由于各地区气候和林分特性存在差异。乔木层是生物量的主体,占总生物量的69.32%—85.60%,不同区域乔木层生物量表现为:延安(109.14 t/hm2)>庆阳(103.01 t/hm2)>平凉2(92.25 t/hm2)>平凉1(31.65 t/hm2)>白银(26.72 t/hm2)。白银和平凉1样地乔木层生物量显著低于其他样地,这可能与这2个样地的刺槐人工林多为幼、中龄林有关。在研究区内只有平凉1、平凉2、延安样地中有灌木,灌木层生物量最大值出现在平凉2样地。不同区域草本层、枯落物层生物量之间差异显著(P<0.05)。
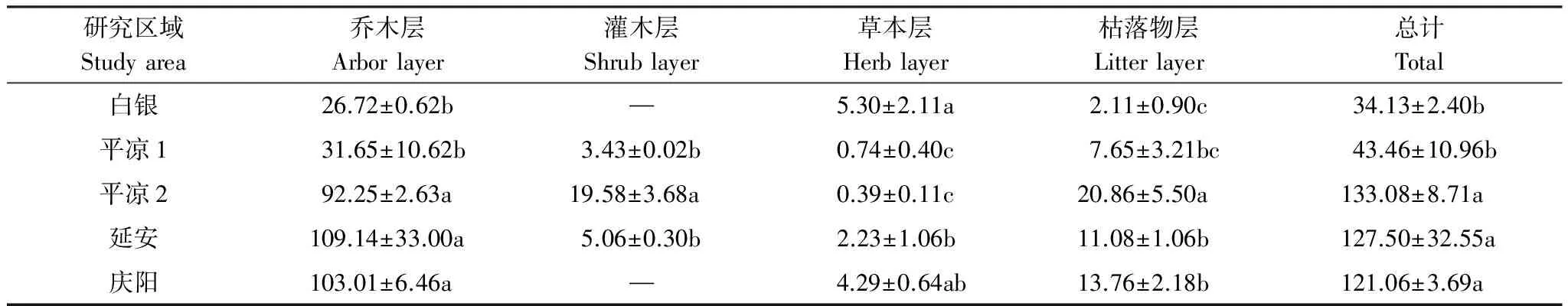
表2 各研究区域刺槐人工林生态系统生物量/(t/hm2)
同列不同小写字母表示不同研究区域间差异显著(α=0.05)
2.2 不同区域刺槐人工林生态系统各组分碳含量
黄土高原中西部刺槐人工林生态系统各组分(枯落层除外)平均碳含量在4个区域(灌木为2个区域)间差异显著(P<0.05)(表3)。乔木层平均碳含量大小顺序为:延安(454.67 g/kg)>白银(434.89 g/kg)>庆阳(430.53 g/kg)>平凉2(430.06 g/kg)>平凉1(429.56 g/kg)。灌木层平均碳含量以平凉1样地最高,平均为434.42 g/kg,是平凉2样地的1.47倍。不同区域草本层、枯落物层平均碳含量分布范围分别为259.89—339.06 g/kg和221.93—315.81 g/kg。草本层、枯落物层平均碳含量的最大值分别出现在延安样地(339.06 g/kg)和庆阳样地(315.81 g/kg)。总体上表现为乔木层平均碳含量高于灌、草层,枯落物层平均碳含量最低,这可能是由于部分枯落物进入分解或半分解状态。

表3 各研究区域刺槐人工林平均碳含量/(g/kg)
2.3 不同区域刺槐人工林生态系统碳密度
黄土高原中西部刺槐人工林生物量碳密度在17.04—58.82 t/hm2之间,不同区域大小顺序依次为:延安(58.82 t/hm2)、庆阳(51.90 t/hm2)、平凉2(44.35 t/hm2)、白银(19.75 t/hm2)、平凉1(17.04 t/hm2)(图1)。乔木层碳密度与生物量碳密度的大小顺序一致。灌木层碳密度以延安样地(2.15 t/hm2)最高,其次为平凉2样地(0.14 t/hm2),最低的为平凉1样地,仅为0.02 t/hm2。草本层碳密度大小顺序为:庆阳(1.46 t/hm2)>白银(1.37 t/hm2)>延安(0.69 t/hm2)>平凉1(0.22 t/hm2)>平凉2(0.13 t/hm2)。枯落物层碳密度在不同区域大小顺序依次为:庆阳(5.40 t/hm2)、平凉2(4.41 t/hm2)、平凉1(3.21 t/hm2)、延安(2.73 t/hm2)、白银(0.56 t/hm2)。

图1 各研究区域刺槐人工林生态系统不同组分碳密度Fig.1 Carbon density in different components of Robinia pseudoacacia plantation ecosystem in each study area
黄土高原中西部刺槐人工林生态系统总碳密度为67.94—135.91 t/hm2,平均碳密度为106.86 t/hm2(图1)。土壤碳密度为48.39—91.57 t/hm2,占生态系统总碳密度的54.83%—74.93%(图1)。土壤层碳密度在不同区域大小顺序依次为:平凉2(91.57 t/hm2)、庆阳(80.19 t/hm2)、延安(71.40 t/hm2)、平凉1(50.91 t/hm2)、白银(48.39 t/hm2)。其中,白银和平凉1与平凉2、延安、庆阳3个样地差异显著(P<0.05)。生态系统碳密度与土壤碳密度在不同样地顺序一致,其中平凉2、延安和庆阳3个样地之间,白银和平凉1之间差异不显著(P>0.05),白银和平凉1与这3个样地有显著性差异(P<0.05)。平凉2样地生态系统碳密度(135.91 t/hm2)是白银样地(67.94 t/hm2)的2倍。不同组分碳密度均值大小排序为:土壤层(68.49 t/hm2)>乔木层(33.88 t/hm2)>枯落物层(3.26 t/hm2)>灌木层(0.77 t/hm2)和草本层(0.77 t/hm2)。
2.4 刺槐人工林生态系统碳密度的影响因子
刺槐人工林生态系统碳密度随林龄增大而显著增大,随林分密度变化趋势不明显(图2)。相关分析结果表明:刺槐人工林生态系统碳密度与林龄间存在极显著相关关系(P<0.01,R2=0.7081),与林分密度间无显著相关关系(P>0.05,R2=0.0084)。刺槐人工林生态系统碳密度随降水量增大而增大,随温度变化趋势不明显(图2)。相关分析结果表明:刺槐人工林生态系统碳密度与降水量间存在显著相关关系(P<0.05,R2=0.3103),与温度间无显著相关关系(P>0.05,R2=0.0017)。
对刺槐人工林碳密度与海拔、坡度做相关分析,结果表明:刺槐人工林生态系统碳密度与海拔、坡度间均无显著相关关系(P>0.05)。这可能与研究区域的海拔和坡度梯度不明显有关。
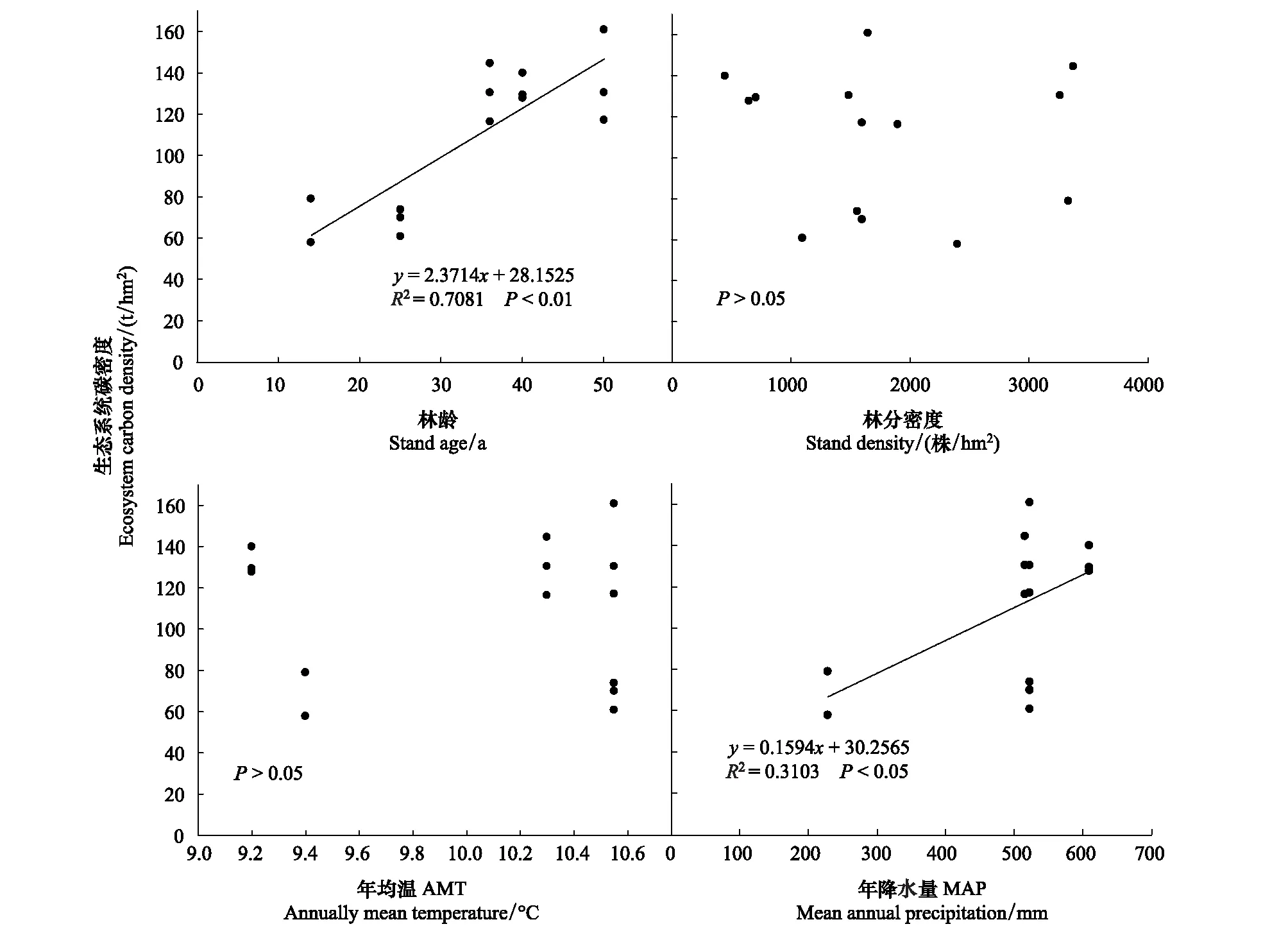
图2 刺槐人工林生态系统碳密度与林龄,林分密度,年均温,年降水量之间的关系Fig.2 The relationship between ecosystem carbon density and stand age, stand density, AMT, MAP
2.5 刺槐人工林生态系统碳密度影响因子的主成分分析
对影响刺槐人工林生态系统碳密度的林分因子(林龄和林分密度)、地形因子(海拔和坡度)和环境因子(年均温和年降水量)进行主成分分析。通过表4可以看出,在特征值大于1的三个主分量中,第一主成分主要综合了林龄、降水量对生态系统碳密度的影响,方差贡献率为37.22%;第二主成分则综合了海拔和温度的变异信息,方差贡献率为34.74%;第三主成分主要综合坡度的信息,方差贡献率为19.10%;上述三个主分量的总方差贡献率达到91.069%,说明前三个主成分能反映林分因子、地形因子和环境因子对刺槐人工林碳密度影响效应的绝大部分信息。第一主成分的方差贡献率最大,说明林龄、降水量对生态系统碳密度起主要的影响作用,其他因子的影响作用相对较小。

表4 刺槐人工林碳密度影响因子主成分的特征值和贡献率
3 结论与讨论
3.1 刺槐人工林生态系统植被层碳含量
研究结果表明,黄土高原中西部刺槐人工林植被层平均碳含量为221.93—454.67 g/kg,总体上表现为乔木层碳含量高于灌、草层,枯落物层碳含量最低,这可能是由于部分枯落物进入分解或半分解状态。可见若采用500 g/kg或450 g/kg作为植被生物量与其碳储量之间的换算系数[17- 19],将在一定程度上高估该地区碳密度。刺槐人工林乔木层平均碳含量(435.94 g/kg)低于该地区辽东栎碳含量(441.74 g/kg)[20]和油松碳含量(503.80 g/kg)[15],这说明该地区刺槐人工林的碳吸存能力较弱。调查区刺槐人工林乔木层平均碳含量高于热带半干旱气候区植被碳含量(373.50 g/kg)[21],但却低于我国南亚热带森林植被碳含量的估计值(531.33 g/kg)[22]。这一方面是由于不同树种碳吸附能力不同,另一方面不同区域环境下的植被接受到的水热条件存在差异。因此,在不同的地区和植被类型下使用相应的碳含量才能够减小估算误差,更加精确地估算植被碳密度。
3.2 刺槐人工林生态系统各组分碳密度
研究结果表明,乔木层与土壤层为刺槐人工林生态系统碳密度的主体,分别占生态系统碳密度的20.01%—40.90%和54.83%—74.93%。本研究区刺槐人工林植被层碳密度总体均值为38.68 t/hm2,略低于李海奎等[23]估算的西北地区植被碳密度(42.82 t/hm2)以及方精云等[24]估算的我国森林植被碳密度(41.00 t/hm2),但是却远低于世界森林碳密度平均值(86.00 t/hm2)[25];土壤碳密度平均值为68.49 t/hm2,远低于中国森林土壤碳密度平均水平(193.55 t/hm2)[26]和世界森林土壤碳密度平均水平(189.00 t/hm2)[25]。土壤碳密度约为植被碳密度的2.02倍,该数值与全球森林土壤碳密度与植被碳密度的比值接近(约为2.2倍)[26]。黄土高原中西部刺槐人工林植被层、土壤层均低于全国平均水平,这主要由于:一方面本研究区的刺槐人工林平均林龄为14—50 a,成熟林和过熟林占的比重较小,故各组分碳密度较小;另一方面黄土高原中西部地区年降水量少且相对集中,易形成严重的水土、养分流失,土壤贫瘠,同时植被生长面临着严重的水分胁迫,林分立地质量较差。
黄土高原中西部刺槐人工林生态系统碳密度随林龄增加而增加,与Li等[29]关于黄土高原刺槐碳密度随林龄变化的研究结果以及Pregizer等[30]关于全球森林与林龄的研究结果一致,说明黄土高原中西部刺槐人工林现阶段仍具有一定的固碳潜力。植被层碳密度在中幼龄林中最小(25a,17.04 t/hm2),其次是成熟林(50a,44.35 t/hm2),而在近熟林达到最大值(36a,58.82 t/hm2),这可能是由于在该区域,刺槐生长受到水分限制,成熟林中普遍存在顶梢枯死及“小老树”现象[27];土壤层碳密度随林龄的增加而增加(从14a的48.39 t/hm2到50 a的91.57 t/hm2),与Foote等[28]关于土壤碳储量与林龄关系的研究结果一致,表明在刺槐林达到成熟林过程中,土壤可作为稳定的碳库存在;生态系统碳密度随林龄增加而增加。因此,在该地区刺槐人工林的经营管理中,应加强对幼中龄林的抚育,同时对成熟林适时、合理间伐,以提高该地区林木的固碳能力和经济效益。
3.3 刺槐人工林生态系统碳密度影响因子
森林生态系统的环境因子之间彼此联系,且交互在一起影响着生态系统碳密度,对单个环境因子的分析不足以解释生态系统碳密度的控制机制[30]。本研究结果表明:刺槐人工林生态系统碳密度与林龄、降水量均呈显著正相关(P<0.01,P<0.05)(图2),与Li等[29,31]的研究结果一致。研究区刺槐人工林林龄介于14—50 a之间,植被处于生长期,随林龄增加,土壤碳密度逐渐积累。生态系统碳密度随林龄增加不断增大,说明黄土高原中西部刺槐人工林在不同林龄阶段均存在持续固碳的潜力。生态系统碳密度与降水量之间存在相关关系[32],本研究区域的降水量范围是200—610 mm,基本不能满足植物生长发育的需要,所以降水量是该地区碳密度的主要限制因子。
生态系统碳密度随林分密度的变化趋势不显著(图2),可能是因为研究样本数偏小,尚无法完全客观的反映出碳密度随林分密度变化的规律。而由于本研究的区域跨度较小,温度梯度变化不大,在该研究区温度不是制约刺槐人工林生态系统碳密度的主要因子(图2),更有可能是和其他因子共同作用影响生态系统碳密度,所以温度不应作为单独的影响因子来考虑[33]。
海拔是导致土壤有机碳密度变异的主导因子[34],对碳密度的影响主要是通过水热条件的改变间接实现;坡度通过对土壤水分和养分的分配以及林冠层接受光照的面积影响生态系统碳密度。本研究区域内,刺槐人工林生态系统碳密度与地形因子(海拔、坡度)无显著相关关系,这可能有两方面原因:一方面研究区域的海拔(1073—1642 m)、坡度变化范围不大;另一方面可能是其他影响因子的作用掩盖了地形因子的作用,主成分分析表明:林龄、降雨量是影响黄土高原中西部刺槐人工林生态系统碳密度的主要因子。另外,刺槐人工林生态系统不仅包括土壤层还包括植被层,而地形因子对植被层的影响相对较小。
不同区域森林生态系统碳密度的差异是温度、水分等环境因子与林分密度、林龄等林分因子以及海拔、坡度等地形因子综合作用的结果。因此在进行较大尺度森林生态系统碳密度的研究时,应综合考虑地形、环境因子、林分因子以及人为管理因素等。
[1] 方精云, 陈安平. 中国森林植被碳库的动态变化及其意义. 植物学报, 2001, 43(9): 967- 973.
[2] Cao M K, Woodward F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change. Global Change Biology, 1998, 4(2): 185- 198.
[3] 王效科, 冯宗炜, 欧阳志云. 中国森林生态系统的植物碳储量和碳密度研究. 应用生态学报, 2001, 12(1): 13- 16.
[4] 刘国华, 傅伯杰, 方精云. 中国森林碳动态及其对全球碳平衡的贡献. 生态学报, 2000, 20(5): 733- 740.
[5] 徐济德. 我国第八次森林资源清查结果及分析. 林业经济, 2014, (3): 6- 8.
[6] 黄从德, 张健, 杨万勤, 张国庆. 四川人工林生态系统碳储量特征. 应用生态学报, 2008, 19(8): 1644- 1650.
[7] 贺亮, 苏印泉, 季志平, 辛占良. 黄土高原沟壑区刺槐、油松人工林的碳储量及其分布特征研究. 西北林学院学报, 2007, 22(4): 49- 53.
[8] 王蕾, 张景群, 王晓芳, 苏印泉. 黄土高原两种人工林幼林生态系统碳汇能力评价. 东北林业大学学报, 2010, 38(7): 75- 78.
[9] 杨晓梅, 程积民, 孟蕾. 黄土高原天然柴松林碳储量与碳密度特征. 中国水土保持科学, 2010, 8(2): 41- 45.
[10] 张景群, 苏印泉, 康永祥, 徐喜明, 秦乐. 黄土高原刺槐人工林幼林生态系统碳吸存. 应用生态学报, 2009, 20(12): 2911- 2916.
[11] 刘迎春, 王秋凤, 于贵瑞, 朱先进, 展小云, 郭群, 杨浩, 李胜功, 胡中民. 黄土丘陵区两种主要退耕还林树种生态系统碳储量和固碳潜力. 生态学报, 2011, 31(15): 4277- 4286.
[12] 艾泽民, 陈云明, 曹扬. 黄土丘陵区不同林龄刺槐人工林碳、氮储量及分配格局. 应用生态学报, 2014, 25(2): 333- 341.
[13] 孟蕾, 程积民, 杨晓梅, 韩娟娟, 范文娟, 胡秀娟. 黄土高原子午岭人工油松林碳储量与碳密度研究. 水土保持通报, 2010, 30(2): 133- 137.
[14] 杨晓梅, 程积民, 孟蕾, 韩娟娟. 黄土高原子午岭森林碳储量与碳密度研究. 水土保持学报, 2010, 24(6): 123- 126.
[15] 申家朋, 张文辉, 李彦华, 何景峰, 张辉. 陇东黄土高原沟壑区刺槐和油松人工林的生物量和碳密度及其分配规律. 林业科学, 2015, 51(4): 1- 7.
[16] 中国科学院先导专项森林固碳课题办公室. 中国森林生态系统固碳现状、速率、机制和潜力课题生物量估算方程. 2014. 中国科学院华南植物园(未公开出版).
[17] 方精云. 中国森林生产力及其对全球气候变化的响应(英文). 植物生态学报, 2000, 24(5): 513- 517.
[18] Kauppi P E, Mielikäinen K, Kuusela K. Biomass and carbon budget of European forests, 1971 to 1990. Science, 1992, 256(5053): 70- 74.
[19] 马钦彦, 陈遐林, 王娟, 蔺琛, 康峰峰, 曹文强, 马志波, 李文宇. 华北主要森林类型建群种的含碳率分析. 北京林业大学学报, 2002, 24(5/6): 96- 100.
[20] 王娟, 陈云明, 曹扬, 周建云, 侯磊. 子午岭辽东栎林不同组分碳含量与碳储量. 生态学杂志, 2012, 31(12): 3058- 3063.
[21] 郑帷婕, 包维楷, 辜彬, 何晓, 冷俐. 陆生高等植物碳含量及其特点. 生态学杂志, 2007, 26(3): 307- 313.
[22] 刘恩, 王晖, 刘世荣. 南亚热带不同林龄红锥人工林碳贮量与碳固定特征. 应用生态学报, 2012, 23(2): 335- 340.
[23] 李海奎, 雷渊才, 曾伟生. 基于森林清查资料的中国森林植被碳储量. 林业科学, 2011, 47(7): 7- 12.
[24] 方精云, 郭兆迪, 朴世龙, 陈安平. 1981—2000年中国陆地植被碳汇的估算. 中国科学D辑:地球科学, 2007, 37(6): 804- 812.
[25] Dixon R K, Brown S, Houghton R A, Solomon A M, Trexler M C, Wisniewski J. Carbon pools and flux of global forest ecosystems. Science, 1994, 263(5144): 185- 190.
[26] 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 2000, 24(5): 518- 522.
[27] 侯庆春, 黄旭, 韩仕峰, 张孝中. 关于黄土高原地区小老树成因及其改造途径的研究 Ⅲ: 小老树的成因及其改造途径. 水土保持学报, 1991, 5(4): 80- 86.
[28] Foote R L, Grogan P. Soil carbon accumulation during temperate forest succession on abandoned low productivity agricultural lands. Ecosystems, 2010, 13(6): 795- 812.
[29] Li T J, Liu G B. Age-related changes of carbon accumulation and allocation in plants and soil of black locust forest on Loess Plateau in Ansai County, Shaanxi Province of China. Chinese Geographical Science. 2014, 24(4): 414- 422.
[30] Pregitzer K S, Euskirchen E S. Carbon cycling and storage in world forests: Biome patterns related to forest age. Global Change Biology, 2004, 10(12): 2052- 2077.
[31] Li T J, Ren B W, Wang D H, Liu G B. Spatial variation in the storages and age-related dynamics of forest carbon sequestration in different climate zones-evidence from black locust plantations on the Loess Plateau of China. PLoS One, 2015, 10(3): e0121862.
[32] Fang J Y, Piao S L, Tang Z Y, Peng C H, Ji W. Interannual variability in net primary production and precipitation. Science, 2001, 293(5536): 1723.
[33] 吕超群, 孙书存. 陆地生态系统碳密度格局研究概述. 植物生态学报, 2004, 28(5): 692- 703.
[34] 解宪丽, 孙波, 周慧珍, 李忠佩. 不同植被下中国土壤有机碳的储量与影响因子. 土壤学报, 2004, 41(5): 687- 699.
EcosystemcarbondensityofRobiniapseudoacaciaplantationsandinfluencingfactorsinthemiddleandwesternLoessPlateau
NIU Chunmei1,2, GUAN Jinhong1, CHENG Ranran2,3, LI Guoqing2,3, WU Chunrong4, CHENG Jimin2,3, DU Sheng2,3,*
1CollegeofForestry,NorthwestA&FUniversity,Yangling712100,China2StateKeyLaboratoryofSoilErosionandDrylandFarmingonLoessPlateau,NorthwestA&FUniversity,Yangling712100,China3InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling712100,China4GansuDesertControlResearchInstitute,Lanzhou730000,China
Plantations are believed to be a mitigating measure against the predicted rising CO2levels. As important carbon sinks,Robiniapseudoacaciaplantations have been planted extensively on the Loess Plateau owing to the ability of this species to adapt to different soil and climatic conditions and its tolerance to drought and erosion. However, little is known regarding the efficiency ofR.pseudoacaciaregarding biomass allocation, ecosystem carbon density partitioning, and factors influencing these aspects, especially in different planted regions. Thus, the objective of the present study was to investigate the ecosystem carbon density ofR.pseudoacaciaplantations and evaluate their relationship with different influencing factors in diverse planted regions. In the present study, a set of 14 sample plots were established in 4 major planted areas ofR.pseudoacaciaplantations (Baiyin, Pingliang, Yan′an, and Qingyang) in the middle and western Loess Plateau, China. Field investigations and laboratory analyses were undertaken to quantify the ecosystem carbon density and analyze its influencing factors. The main results are that the average biomass in the plantation ecosystems of five different age groups was 34.13—133.08 t/hm2and the average carbon concentration of the vegetation pool was 221.93—454.67 g/kg. The average biomass in different components was significantly different among the four distribution regions, as were the average carbon concentrations of arbor layer, shrub layer, and herb layer. The carbon concentrations in the different components were in the order of: arbor layer (435.94 g/kg) > shrub layer (365.19 g/kg) > herb layer (310.53 g/kg) > litter layer (264.08 g/kg). The average carbon density of the studied ecosystems was 106.86 t/hm2. The proportion of soil layer carbon density was 64.09%, and soil layer carbon density was the major constituent of the carbon density of the ecosystem. The carbon density of the vegetation layer was 38.68 t/hm2. The carbon density of the arbor layer was 33.88 t/hm2, which accounted for 87.58% of the vegetation layer carbon density. The carbon density of the shrub layer, herb layer, and litter layer were 0.77 t/hm2, 0.77 t/hm2, and 3.26 t/hm2, respectively, which accounted for 1.98%, 2.00%, and 8.43% of the vegetation layer, respectively. Soil organic carbon density and ecosystem carbon density were all significantly different among the four regions. Correlation analysis indicated that ecosystem carbon density was positively correlated with stand age and precipitation. In contrast, its relationships with forest stand density, slope gradient, elevation, and annually mean temperature were not statistically significant. Principal component analysis indicated that stand age and precipitation were the first principal component affecting ecosystem carbon density; elevation and annually mean temperature were the second principal component; and the slope gradient was the third principal component. These variables accounted for 91.07% of the variance of ecosystem carbon density. The effect of different factors on ecosystem carbon density was discrepant. Stand age and precipitation were the primary factors affecting ecosystem carbon density. The differences in forest ecosystem carbon density were due to the combined effects of climate, topography, forest stand factors, and management. Therefore, these influencing factors need to be considered when using forest-investigated data to estimate forest ecosystem carbon stock, especially at larger scales.
middle and western Loess Plateau;Robiniapseudoacaciaplantations; carbon density; biomass
中国科学院战略性先导科技专项资助项目(XDA05050202)
2016- 04- 29; < class="emphasis_bold">网络出版日期
日期:2017- 03- 22
10.5846/stxb201604290824
*通讯作者Corresponding author.E-mail: shengdu@ms.iswc.ac.cn
牛春梅,关晋宏,程然然,李国庆,吴春荣,程积民,杜盛.黄土高原中西部刺槐人工林生态系统碳密度及其影响因子.生态学报,2017,37(15):5049- 5058.
Niu C M, Guan J H, Cheng R R, Li G Q, Wu C R, Cheng J M, Du S.Ecosystem carbon density ofRobiniapseudoacaciaplantations and influencing factors in the middle and western Loess Plateau.Acta Ecologica Sinica,2017,37(15):5049- 5058.
