不同园林植物土壤呼吸及影响因子特征
2017-09-12司志国
夏 冰, 司志国
(河南职业技术学院 环境艺术工程系, 郑州 450046)
不同园林植物土壤呼吸及影响因子特征
夏 冰, 司志国
(河南职业技术学院 环境艺术工程系, 郑州 450046)
以郑州市4种园林植物(合欢、龙爪槐、紫叶李和海棠)为试材,采用动态密闭气室红外CO2分析仪(IRGA)法测定了不同园林植物土壤根区土壤呼吸特征,并通过测定园林植物根区土壤养分含量和酶活性等土壤环境因子(土壤温度和湿度等),研究不同园林植物根区土壤呼吸影响因子,为城市园林植被建设及景观配置提供科学依据。结果表明:(1) 不同园林植物大气温度的日变化趋势均为单峰曲线;大气湿度日变化均呈“V”字形变化趋势;0—10 cm土壤温度与大气温度变化趋势相一致,在14:00左右达到峰值;土壤湿度均呈“V”字形变化趋势,在14:00达到最低,此后有所回升;(2) 在日变化尺度上,4种园林植物土壤呼吸均表现为单峰型,且峰值出现的时间基本一致,在14:00左右达到最大,最低值出现在早上6:00,相同时间土壤呼吸速率基本表现为合欢>龙爪槐>紫叶李>海棠;(3) 4种园林植物土壤呼吸与土壤温度和土壤湿度之间关系以指数方程拟合最好;(4) 4种园林植物土壤养分含量和土壤酶活性的变化趋势相一致,表现为合欢>龙爪槐>紫叶李>海棠,而4种园林植物根区土壤全磷含量差异不显著(p>0.05);(5) 偏相关性分析可知,不同园林植物土壤呼吸均与土壤养分和土壤酶活性呈现出一定的相关性,其中合欢和龙爪槐的相关系数高于紫叶李和海棠。
园林植物; 土壤呼吸; 土壤养分; 土壤酶活性
园林植物是城市—自然—景观复合生态系统,作为城市生态环境建设的主体,在改善生态环境、减少阳光辐射、空气湿度和调节气候等方面具有重要作用,已成为衡量城市生态文明和城市现代化的重要标志[1-3]。此外,园林植物还可以通过自身的蒸腾、蒸散、吸收、吸附、反射等功能,降低外界温度、固碳释氧、抗污染(吸收粉尘,Cl2,SO2,CO等)、降低噪音、保护多样性等[4-5]。随着生态城市概念的提出、建设和发展,人们越来越注意到园林植物在城市生态中的意义和价值。另一方面,城市园林植物作为城市生态系统的重要组成部分,不仅会影响城市的生态环境质量,而且城市园林植物的配置是否合理也将对城市的热岛效应及全球的碳变化具有一定的影响[4-5]。土壤作为生态系统中的重要组成部分,在生态系统的物质循环和能量流动方面起着重要作用[6-8]。土壤呼吸释放的CO2是全球碳循环中活跃的组成部分,对土壤碳释放及大气中碳平衡起着至关重要的作用,主要包括土壤微生物呼吸、根系呼吸、土壤动物呼吸的生物学过程[6,9]。据不完全统计,全世界土壤呼吸每年向大气释放的碳高达80~100 Pg,大约是化石燃料燃烧释放CO2的10倍之多,而在陆地生态系统中,土壤碳库是大气碳贮量的2~3倍,通过土壤呼吸作用向大气释放的CO2约占全球CO2交换量的1/4左右[7-8],深入认识生态系统土壤呼吸过程对减少全球碳预算的不确定性与预测未来气候变化具有重要意义[10]。
当前,尽管国内外诸多学者对陆地土壤呼吸做了大量深入的研究[11-13],大部分研究集中在森林、草地和农田等生态系统中,对城市园林植物土壤呼吸研究则鲜有报道。然而,在全球城市化加速的背景下,城市景观正在快速向自然和农业生态系统蔓延,再加上生态城市中的不确定因素导致园林植物土壤呼吸速率存在的地域性差异,使得量化土壤呼吸及其影响因子尤为困难,特别是对于发展中的生态环保型城市,研究生态型城市不同园林植物土壤呼吸特征及其影子因子显得尤为重要[14-15],因此,研究园林植物土壤呼吸不仅有助于了解其在区域碳循环中的作用,还有利于深入认识城市化对陆地生态系统碳循环的影响。有鉴于此,本文通过对郑州市大型公园现状分布的全面调查,最后确定在人民公园、碧沙岗公园、紫金山公园和世纪欢乐园选择4种园林植物测量园林植物土壤呼吸速率及土壤影响因子(园林植物根区土壤养分和酶活性),旨在探讨不同园林植物土壤呼吸日变化特征,分析土壤温度、湿度、土壤养分及酶活性对土壤呼吸的影响,通过相关性分析探讨园林植物土壤呼吸的影响因子,以期深入了解生态型城市园林植物土壤呼吸作用的变化过程及变化机理,从而为城市生态系统碳循环和城市建设中园林植物的选择及其应用提供基础数据和理论支持。
1 材料与方法
1.1 研究区自然概况
郑州市地处河南省中部偏北,黄河中游的南岸(112°42′—114°14′E,34°16′—34°58′N),北临黄河,西依嵩山,东南部依靠黄淮平原。属暖温带大陆性季风气候,四季分明,呈现春季温暖少雨,夏季炎热多雨,秋季晴朗日照长,冬季寒冷多风。年平均气温14.4℃,极端最高温度42.3℃,极端最低温度-17.9℃,7月份最热,平均27.3℃,1月份最冷,平均0.2℃。年降水量652.9 mm,多集中在夏季(6—8月),占全年降水量的52.3%。土壤类型较多,以褐土和潮土分布面积最大,分别占土壤面积的64.4%和30.17%,土壤湿度较大,地势差异较为悬殊,为多种园林植物生长提供了有利的环境生活条件。郑州市园林植物种类繁多,可开发利用的园林观赏植物资源丰富,据不完全统计,种子植物共2 000多种,属于150科800多属,与全国植物科属种数相比,科占35.2%,属占16.4%,种占8.4%,足以说明郑州市植物区系成分的丰富,园林植物约有42科106属200余种(含变种、变型和亚种)。20世纪80年代中期,郑州市绿化覆盖面积居全国省会城市前列,为郑州赢得了“绿城”的美誉。2000年郑州市提出创建国家园林城市的目标,通过大规模的建造绿地工程,先后获得“河南省园林城市”、“全国园林绿化先进城市”,2006年正式成为“国家园林城市”。
1.2 土壤呼吸测定
2015年8—9月,对郑州市内的不同公园园林植物进行调查统计,结果发现:合欢(Albizziajulibrissin)、龙爪槐(Sophorapendula)、海棠(Prunuscerasifera)、紫叶李(Chaenomelesspeciosa)为园林植物中的主要园林植物,故选这4种具有代表性的园林植物作为试验试材(表1)。

表1 不同园林植物基本生长特性
在郑州市人民公园、碧沙岗公园、紫金山公园和世纪欢乐园选择4种园林植物(合欢、龙爪槐、海棠、紫叶李),每种园林植物选取3株,分别在其根区(主根分布周围)布设监测点并标记,在第一次测定土壤呼吸之前,提前一天将测定基座嵌入土壤中,在每个基座内存留的地表植被自土壤表层彻底剪除,聚氯乙烯圆柱体经过24 h的平衡后,土壤呼吸速率会恢复到基座放置前的水平,从而避免了因安置气室对土壤扰动而造成的短期内呼吸速率波动。采用动态密闭气室红外CO2分析仪(IRGA)法,测定仪为美国Li-6400便携式气体分析系统和Li-6400土壤呼吸室,土壤呼吸日变化测定为白天每隔2 h测定1次,在6:00—18:00点之间测量,每个监测点1次测定2个重复取平均值,在观测土壤呼吸的同时,使用手持长杆电子温度探针测定0—10 cm深处的土壤温度,同时使用TDR测定每个监测点附近0—10 cm范围内的土壤湿度,同时取监测园林植物根区0—10 cm混合土样两份,带回实验室—份自然风干(15~20 d)去除碎片和部分根后过0.5 mm筛,测定土壤养分含量,另一份保温冰箱4℃保存用于测定土壤酶活性。
1.3 土壤养分的测定
土壤养分的测定参照鲍士旦的《土壤农化分析》[16]:土壤有机碳含量(g/kg)测定采用重铬酸钾氧化外加热法;全氮(g/kg)用凯氏定氮法;全磷(g/kg)用NaOH熔融—钼锑抗比色法;全钾(g/kg)采用火焰分光光度法。
土壤酶活性的测定:根区土壤酶活测定用分光光度计进行比色法测定,测定酶活种类为纤维素酶(1 g土样30 min内分解产生1 mg葡萄糖所需的酶量);转化酶(1 g土样24 h内分解产生1 mg葡萄糖所需的酶量);脲酶(1 g土样24 h内分解产生1 mg氨基氮所需的酶量)[16]。
采用Excel 2007和SPSS 18.0进行数据分析,单因素方差分析(One-way ANOVA),Pearson相关性系数检验各指标的相关性。采用指数方程对土壤呼吸(Rs)与土壤温度和湿度间进行回归分析。由原始数据拟合得到的多元回归关系经统计学检验得到拟合度参数R2,并在p<0.05和p<0.01水平检验相关系数的显著性。
2 结果与分析
2.1 不同园林植物环境因子日动态
4种不同园林植物所在大气温度日变化规律基本一致(图1),大气温度的日变化趋势均为单峰曲线,在早上6:00左右最低,在14:00左右达到峰值,14:00之后有降低的趋势;4种不同园林植物所在大气湿度日变化规律基本一致,大气湿度日变化与大气温度的日变化趋势相反,在早上6:00左右最高,在16:00左右达到峰值(最低),16:00以后有回升的趋势;4种不同园林植物土壤温度与大气温度变化趋势相一致,最大值有所延迟,在16:00左右达到峰值,14:00之后有降低的趋势;4种不同园林植物土壤湿度与大气温度变化趋势相一致,均呈“V”字形变化趋势,在14:00左右达到最低,此后有所回升,不同园林植物最低值有所波动。
2.2 不同园林植物土壤呼吸日变化
4种园林植物土壤呼吸具有明显的日变化规律,且均为单峰型,峰值出现时间基本一致(图2)。土壤呼吸速率均在14:00左右达到最大(土壤和大气温度最高的时刻),最低值出现在早上6:00和晚上18:00,在6:00—8:00,4种园林植物土壤呼吸升高缓慢,8:00以后急剧上升,达到最大值以后急剧降低,在18:00以后,基本达到平稳的趋势。合欢土壤呼吸日变化范围为0.56~1.93 μmol/(m2·s),龙爪槐土壤呼吸日变化范围为0.52~1.87 μmol/(m2·s),海棠土壤呼吸日变化范围为0.53~1.65μmol/(m2·s),紫叶李土壤呼吸日变化范围为0.55~1.73 μmol/(m2·s),相同时间土壤呼吸速率基本表现为合欢>龙爪槐>紫叶李>海棠。
2.3 园林植物根区土壤养分
4种园林植物根区土壤有机碳含量变化范围为11.5~14.2 g/kg,具体表现为合欢>龙爪槐>紫叶李>海棠(图3),其中合欢和龙爪槐根区土壤有机碳含量差异不显著(p>0.05),二者显著高于海棠和紫叶李(p<0.05),海棠和紫叶李根区土壤有机碳含量差异不显著(p>0.05);4种园林植物根区土壤全氮含量变化范围为1.12~1.35 g/kg,具体表现为合欢>龙爪槐>紫叶李>海棠,其中合欢和龙爪槐根区土壤全氮含量差异不显著(p>0.05),二者显著高于海棠和紫叶李(p<0.05),海棠根区土壤全氮含量最低(p<0.05),显著低于其他植物(p<0.05);4种园林植物根区土壤全磷含量变化范围为0.86~0.91 g/kg,具体表现为合欢=海棠>紫叶李>龙爪槐,4种园林植物根区土壤全磷含量差异并不显著(p>0.05);4种园林植物根区土壤全钾含量变化范围为16.17~28.35 g/kg,具体表现为合欢>龙爪槐>紫叶李>海棠,其中合欢和龙爪槐根区土壤全钾含量差异不显著(p>0.05),二者显著高于海棠和紫叶李(p<0.05),海棠根区土壤全钾含量最低(p<0.05),显著低于其他植物(p<0.05)。

图1不同园林植物环境因子日动态变化
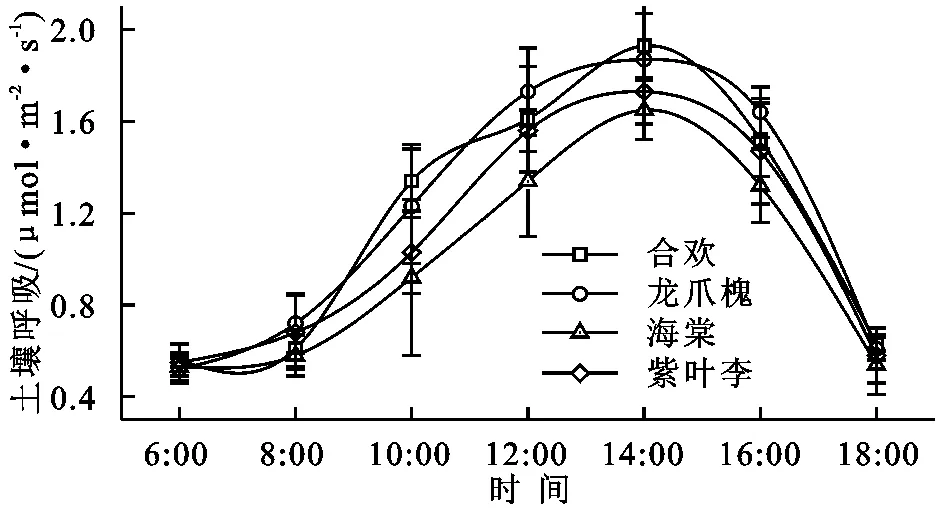
图2园林植物土壤呼吸日变化
2.4 园林植物根区土壤酶活性
4种园林植物根区土壤纤维素酶活性变化范围为1.5~2.5 mg/(g·min),具体表现为合欢>龙爪槐>紫叶李>海棠(图4),其中合欢和龙爪槐根区土壤纤维素酶活性差异不显著(p>0.05),二者显著高于海棠和紫叶李(p<0.05),海棠根区土壤纤维素酶活性最低(p<0.05),显著低于其他植物(p<0.05);4种园林植物根区土壤脲酶活性变化范围为3.54~6.32 mg/(g·24 h),具体表现为合欢>龙爪槐>紫叶李>海棠,其中不同园林植物根区土壤脲酶活性差均异显著(p<0.05);4种园林植物根区土壤转化酶活性变化范围为2.46~6.23 mg/(g·24 h),具体表现为合欢>龙爪槐>紫叶李>海棠,其中不同园林植物根区土壤转化酶活性差均异显著(p<0.05);4种园林植物根区土壤转化酶活性变化范围为1.43~2.73 mg/(g·24 h),具体表现为合欢>龙爪槐>紫叶李>海棠,其中合欢和龙爪槐根区土壤转化酶活性差异不显著(p>0.05),二者显著高于海棠和紫叶李(p<0.05),海棠和紫叶李根区土壤转化酶活性差异不显著(p>0.05)。
2.5 土壤温度和湿度对土壤呼吸的影响
为进一步探讨园林植物土壤呼吸与其显著影响因子土壤温度和湿度的关系,国内外学者一般采用线性模型、二次方程、指数模型等多种方法进行拟合,本试验结果中将土壤呼吸强度与土壤0—10 cm温度进行曲线拟合,筛选出最佳拟合度的方程(R2最大,p值最小),得出的不同拟合方程,由表2可知,土壤呼吸与土壤温度之间关系以指数方程拟合最好。4种园林植物土壤呼吸与土壤温度的指数关系达到了极显著水平(p<0.01),且指数模型的决定系数较大,故指数模型的拟合效果最好,从指数模型来看,合欢根区土壤呼吸与土壤温度的指数关系具体表现为y=6×10-4x3.3569(R2=0.8569,p<0.01,F=95.36),龙爪槐根区土壤呼吸与土壤温度的指数关系具体表现为y=1×10-4x8.3654(R2=0.8236,p<0.01,F=92.41),海棠根区土壤呼吸与土壤温度的指数关系具体表现为y=4×10-4x2.5847(R2=0.7234,p<0.05,F=85.69),紫叶李根区土壤呼吸与土壤温度的指数关系具体表现为y=3×10-4x2.0345(R2=0.7812,p<0.01,F=89.37)。4种园林植物土壤呼吸与土壤湿度之间关系以指数方程拟合最好,合欢根区土壤呼吸与土壤温度湿度的指数关系具体表现为y=0.5623x-1.9635+2.0369(R2=0.8835,p<0.01,F=102.54),龙爪槐根区土壤呼吸与土壤湿度的指数关系具体表现为y=0.0523x-3.0214+0.5212(R2=0.8914,p<0.01,F=1113.25),海棠根区土壤呼吸与土壤湿度的指数关系具体表现为y=0.5694x-2.0361+1.0325(R2=0.8023,p<0.01,F=95.26),紫叶李根区土壤呼吸与土壤湿度的指数关系具体表现为y=0.6132x-1.0324-0.1367(R2=0.8157,p<0.01,F=98.37)。

注:不同小写字母表示差异显著(p<0.05),下同。
图3不同园林植物根区土壤养分

图4不同园林植物根区土壤酶活性
2.6 园林植物土壤呼吸的主要影响因素
土壤呼吸主要是植物的根以及土壤微生物的呼吸,植物的生长、微生物的氧化分解等活动都要受周围环境如土壤温度、水分、气温、空气湿度等的影响,因此,这些环境因子也是影响土壤呼吸强度的重要因素。4种园林植物土壤呼吸强度与各环境因子的Pearson相关关系均显著(表2),但有时由于某一共同变量的干扰会导致2个变量间的假相关,为排除其他因素的干扰,对各因子分别作偏相关分析,控制其他几个变量以分析土壤呼吸与另一个变量的净相关关系。结果表明,合欢根区土壤呼吸与有机碳、全氮和全钾含量呈极显著的相关关系(p<0.01),与纤维素酶、转化酶活性和蔗糖酶活性呈显著的相关关系(p<0.05);龙爪槐根区土壤呼吸与有机碳、全氮和全钾含量呈极显著的相关关系(p<0.01),与脲酶和转化酶活性呈显著的相关关系(p<0.05);海棠根区土壤呼吸与有机碳、全氮和转化酶活性呈极显著的相关关系(p<0.01),与全磷、全钾、纤维素酶和蔗糖酶活性呈显著的相关关系(p<0.05);紫叶李根区土壤呼吸与全氮和转化酶活性呈极显著的相关关系(p<0.01),与有机碳、全钾和蔗糖酶活性呈显著的相关关系(p<0.05)。

表2 土壤温度和湿度对土壤呼吸的影响
注:*表示p<0.05;**表示p<0.01。

表3 不同园林植物土壤呼吸的主要影响因素
注:*和**分别表示在0.05,0.01水平相关性显著和极显著(双尾检验)。
3 讨 论
4种园林植物所在的大气温度和湿度均存在明显的日变化规律,通过测量气温的日变化可知,在早、晚差值可达10℃或者以上。白天较高的温度有利于植物进行光合作用,下午和晚上较低的温度下,园林植物呼吸作用渐弱,有利于有机物质的积累。一天之中14:00左右温度达到最高,由于土壤呼吸速率对地表温度响应较为明显,因此4种园林植物土壤呼吸速率在此时段达到峰值。本研究中园林植物土壤呼吸日变化呈现明显的单峰曲线,日变化峰值出现在14:00,最小值出现在早上6:00左右,此时段地表温度较低、湿度较大,随时间的变化,气温逐渐上升,土壤呼吸速率也逐渐增加,土壤中的微生物和根系呼吸旺盛,导致CO2排放强度增大,土壤呼吸达到最高值,根系和微生物活性最强,此后,随着温度的降低,土壤呼吸则急剧降低[6,9]。除此之外,土壤呼吸速率与土壤(0—10 cm)温度的变化趋势基本吻合,但不同园林植物土壤(0—10 cm)温度的峰值出现较土壤呼吸峰值的出现均有所延迟,主要是由于土壤(0—10 cm)温度达到高峰期需要一个热传递过程。
土壤呼吸速率主要由温度、湿度、环境因子等共同作用所驱动,对于城市园林植物生态系统,水热环境因子是影响其土壤呼吸最为主要的两大因素,土壤湿度在一定程度上降低土壤呼吸速率对土壤温度的响应,通常情况下,土壤湿度会促进土壤呼吸[11-12]。本研究中园林植物土壤呼吸作用受土壤湿度的影响较为明显,土壤呼吸速率与土壤(0—10 cm)湿度呈负线性相关关系,与此同时,土壤温度升高,根系呼吸和土壤生物活性增强,土壤中产生的CO2增多,土壤温度影响土壤中CO2向大气的输送过程,CO2向大气的排放增强,因此土壤温度与土壤呼吸呈正的线性相关[11-12]。通过进一步的相关性分析可知(表3),不同的园林植物土壤呼吸均与土壤养分和土壤酶活性有一定的相关性。
土壤养分主要来自于地表枯枝落叶层的分解补充与积累[17-18]。本研究中4种园林植物土壤养分含量不尽一致,主要受土壤母质及地上植物有效吸收的影响,土壤养分含量也有一定的差异,在城市绿化进程中我们可以根据植物对养分的需求量来选择合适的园林植物。然而4种园林植物土壤全磷差异并不显著,主要是由于磷素作为一种沉积性元素,其分解作用缓慢;土壤酶活性是土壤养分循环和土壤微生物代谢活性的重要指标,能够反映土壤养分累积、分解转化规律和土壤中各种生化过程的强度及其方向。不同园林植物对环境的敏感度导致了根区土壤酶活性的差异,因此土壤酶活性与土壤养分的变化趋势相一致,这与前人的研究结果相吻合[19]。此外,不同园林植物在代谢过程中各种酶系的活力对环境的变化都很敏感,微弱的环境也会对根细胞中酶系的活力产生影响,进而影响其土壤呼吸,因此园林植物对环境的胁迫适应性反应仍是一系列的复杂生理生态学问题。
4 结 论
(1) 不同园林植物大气温度的日变化趋势均为单峰曲线;大气湿度日变化均呈“V”字形变化趋势;0—10 cm土壤温度与大气温度变化趋势相一致,在14:00左右达到峰值;土壤湿度均呈“V”字形变化趋势,在14:00达到最低,此后有所回升。
(2) 在日变化尺度上,4种园林植物土壤呼吸均表现为单峰型,且峰值出现的时间基本一致,在14:00左右达到最大,最低值出现在早上6:00,相同时间土壤呼吸速率基本表现为合欢>龙爪槐>紫叶李>海棠。
(3) 4种园林植物土壤呼吸与土壤温度和土壤湿度之间关系以指数方程拟合最好。园林植物土壤养分含量和土壤酶活性的变化趋势相一致,表现为合欢>龙爪槐>紫叶李>海棠,而4种园林植物根区土壤全磷含量差异不显著(p>0.05)。偏相关性分析可知,不同园林植物土壤呼吸均与土壤养分和土壤酶活性呈现出一定的相关性。
本文简单地分析了不同园林植物根区土壤呼吸影响因子,研究发现温度是影响园林植物根区土壤呼吸的主要环境因子,另外在土壤养分和酶活性对土壤呼吸速率也产生了很大影响,但因试验周期较短,涉及的影响因子还不够全面,应进行深入的、连续的观测,而且更多情况下是多个因子相结合共同对土壤呼吸作用产生影响。因此,城市园林植物土壤呼吸特征分析需要更细致、更详细的研究,进而为全球陆地土壤碳循环的研究提供科学参考。
[1] Smith D R, Asmail S R. Next-generation sequencing data suggest that certain nonphotosynthetic green plants have lost their plastid genomes[J]. New Phytologist, 2014,204(1):7-11.
[2] Kusaba M, Tanaka A, Tanaka R. Stay-green plants:what do they tell us about the molecular mechanism of leaf senescence[J]. Photosynthesis Research, 2013,117(1/3):221-234.
[3] Strehler B L, Arnold W. Light production by green plants[J]. The Journal of General Physiology, 1951,34(6):809-820.
[4] Horton P, Ruban A V, Walters R G. Regulation of light harvesting in green plants[J]. Annual Review of Plant Biology, 1996,47(1):655-684.
[5] Karhu K, Auffret M D, Dungait J A J, et al. Temperature sensitivity of soil respiration rates enhanced by microbial community response[J]. Nature, 2014,513(7516):81-84.
[6] Keidel L, Kammann C, Grünhage L, et al. Positive feedback of elevated CO2on soil respiration in late autumn and winter[J]. Biogeosciences Discussions, 2014,11:8749-8787.
[7] Hamdi S, Moyano F, Sall S, et al. Synthesis analysis of the temperature sensitivity of soil respiration from laboratory studies in relation to incubation methods and soil conditions[J]. Soil Biology and Biochemistry, 2013,58:115-126.
[8] Aanderud Z T, Jones S E, Schoolmaster D R, et al. Sensitivity of soil respiration and microbial communities to altered snowfall[J]. Soil Biology and Biochemistry, 2013,57:217-227.
[9] Lovelock C E, Feller I C, Reef R, et al. Variable effects of nutrient enrichment on soil respiration in mangrove forests[J]. Plant and Soil, 2014,379(1/2):135-148.
[10] Fekete I, Kotroczó Z, Varga C, et al. Alterations in forest detritus inputs influence soil carbon concentration and soil respiration in a Central-European deciduous forest[J]. Soil Biology and Biochemistry, 2014,74:106-114.
[11] Gomez-Casanovas N, Anderson-Teixeira K, Zeri M, et al. Gap filling strategies and error in estimating annual soil respiration[J]. Global Change Biology, 2013,19(6):1941-1952.
[12] Birge H E, Conant R T, Follett R F, et al. Soil respiration is not limited by reductions in microbial biomass during long-term soil incubations[J]. Soil Biology and Biochemistry, 2015,81:304-310.
[13] Whitaker J, Ostle N, Nottingham A T, et al. Microbial community composition explains soil respiration responses to changing carbon inputs along an Andes-to-Amazon elevation gradient[J]. Journal of Ecology, 2014,102(4):1058-1071.
[14] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[15] Barber S A. Soil nutrient bioavailability:a mechanistic approach[M]. John Wiley&Sons, 1995.
[16] Stoorvogel J J, Smaling E M A. Assessment of soil nutrient depletion in Sub-Saharan Africa: 1983—2000[M]. Wageningen: Winand Staring Centre, 1990.
[17] Dick R P. Soil enzyme activities as indicators of soil quality[J]. Defining soil quality for a sustainable environment, 1994 (definingsoilqua):107-124.
[18] Bandick A K, Dick R P. Field management effects on soil enzyme activities[J]. Soil Biology and Biochemistry, 1999,31(11):1471-1479.
[19] Caldwell B A. Enzyme activities as a component of soil biodiversity:a review[J]. Pedobiologia, 2005,49(6):637-644.
SoilRespirationandImpactFactorsofDifferentGreenPlantsinZhengzhouCity
XIA Bing, SI Zhiguo
(DepartmentofEnvironmentalArtEngineering,He′nanVocationalandTechnicalInstitute,Zhengzhou450052,China)
To explore soil respiration and impact factors under different green plants in Zhengzhou City by IRGA, taking four green plants (Albizziajulibrissin,Sophorapendula,PrunuscerasiferaandChaenomelesspeciosa) as experimental materials, we investigated the soil nutrients, soil enzyme activities (0—10 cm) and other environmental factors, and used exponential model to analyze the relations between soil respiration and soil temperature and humidity, to provide the scientific basis for the ecological landscape configuration, and to choose suitable greening plants and urban greening construction. The results showed that: (1) the daily air temperature showed a single peak curve, and daily atmospheric moisture had a variation trend of ‘V’ shape, soil temperature at 0—10 cm depth had a same variation trend with atmospheric temperature, which the peak occurred at 14:00, and soil moisture had a variation trend of ‘V’ shape with the local fluctuation; (2) the daily soil respiration rate showed a single peak curve, and the daily maximum of soil respiration rate occurred at 14:00 and the minimum of soil respiration rate occurred at 6:00, and at same time in a day, the soil respiration rates decreased in the order:Albizziajulibrissin>Sophorapendula>Prunuscerasifera>Chaenomelesspeciosa; (3) it was found that soil respiration was extremely significantly correlated with soil temperature and soil moisture content, thier relation could be described by exponential equation (p<0.05); (4) the soil nutrients and soil enzyme activities under these four green plants decreased in the sequence:Albizziajulibrissin>Sophorapendula>Prunuscerasifera>Chaenomelesspeciosa, while the total soil phosphorus had no difference under these four green plants (p>0.05); (5) through the partial correlation analysis to exclude other environment factors, we found that soil respiration had a correlation with soil nutrients and soil enzyme activities to some degree, and the correlation coefficient underAlbizziajulibrissinandSophorapendulawere higher than those underPrunuscerasiferaandChaenomelesspeciosa.
green plants; soil respiration; soil nutrients; soil enzyme activities
2016-08-18
:2016-08-30
2016河南省科学技术厅软科学研究计划资助“创新区视域下的低碳型中原特色景观构建研究” (162400410327)
夏冰(1984—),女,河南郑州人,硕士,讲师,主要研究方向:园林植物生态学。E-mail:Xia_bing84@163.com
司志国(1977—),男,河南长垣人,博士,副教授,研究方向:城市生态。E-mail:ZHI_G_S@sina.com
Q945.19
:A
:1005-3409(2017)05-0240-07
