东亚森林、草地碳利用效率及碳通量空间变化的影响因素分析
2017-09-12陈云明唐亚坤
安 相, 陈云明, 唐亚坤
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100; 3.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
东亚森林、草地碳利用效率及碳通量空间变化的影响因素分析
安 相1, 陈云明2,3, 唐亚坤2,3
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.西北农林科技大学 水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100; 3.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
森林和草地是陆地生态系统的重要组成部分,研究整合基于涡度相关法观测碳通量的已发表文献,共选取东亚地区40个拥有1年以上数据的通量站(森林26个,草地14个),分析碳利用效率(CUE)以及净生态系统生产力(NEP)、总生态系统生产力(GEP)、生态系统呼吸(RE)的空间变异特征及其影响因素。东亚地区的森林和草地为碳汇,且森林的碳汇显著高于草地(p<0.001),其NEP分别为328.64±256.46 gC/(m2·a),63.43±42.99 gC/(m2·a)。森林和草地的CUE分别为0.21,0.20,影响森林CUE变化的因素主要是林龄,呈线性负相关关系(p<0.001)。影响草地CUE的因素主要是年均降水量(MAP),呈线性负相关关系(p<0.05)。森林和草地的GEP,RE均是随纬度的升高而线性降低,NEP与纬度呈现二次函数关系,随纬度升高而先升高后降低。森林和草地的GEP,RE都与MAP呈线性正相关关系,且NEP与MAP呈先升高后降低的二次函数关系,其中森林和草地的饱和降水量大约为1 300 mm和390 mm。森林的GEP,RE与年均温(MAT)呈现线性正相关关系。森林和草地的GEP,RE与增强型植被指数(EVI)呈线性正相关关系。
森林; 草地; 碳利用效率; 碳通量; 涡度相关法
大气中有1/3的碳都被陆地生态系统固定[1],因此,准确评价区域乃至全球的碳收支变化对于“碳失踪”问题的研究具有重要意义[2]。森林和草地大约占到全球陆地生态系统的51%(其中森林约占31%,草地约占20%),是陆地生态系统的重要组成部分[3-4],有研究认为森林生态系统是主要的碳汇,而草地生态系统在不同的时空条件下呈碳源/汇转化的现象[5]。东亚地区是陆地生态系统重要的组成部分,因此,探明该区域森林和草地碳收支状况、碳利用效率特征及其影响因素,有助于了解气候变化背景下全球碳循环变化趋势并为植被碳汇的管理提供依据。
生态系统碳利用效率(CUE)是净初级生产力(NPP)与总初级生产力(GPP)的比值,是表征生态系统固碳能力及效率的重要参数[6-7]。Delucia等综合了1975—2005年多个通过生物计量法和模型估计法获得森林CUE的研究[8],认为不同类型森林的CUE在0.20~0.83之间波动,且发现CUE与林龄具有线性相关关系。Piao等利用生物计量法和涡度相关法获得的全球碳变化数据集[9],认为CUE可以达到0.5左右,并且认为森林CUE与温度间呈二次函数曲线关系。Zhang等利用MODIS对全球尺度陆地生态系统CUE的研究表明,其与温度和降水具有很好的相关性[10]。目前很多研究多是利用遥感法观测进行研究,但不同的研究对CUE的估算存在很大差异[11],因此,需要使用直接观测的数据分析CUE的现状及其影响因素,才能为模型的修正以及未来CUE的准确模拟提供支持。另外,CUE的空间变异特征及影响因素还不明确,且对草地生态系统碳利用效率的研究则更是鲜有报道。
GEP,RE,NEP是计算CUE的重要组成部分[7],近年来,随着涡度相关技术的进步,已经成为直接观测通量的标准方法[12],不少研究者在不同时空尺度下使用该观测法对森林、草地等生态系统的碳通量特征进行了相关研究。有研究认为GEP,NEP,RE具有一定的纬向分布差异[13-14]。Chen等对影响北半球陆地生态系统碳通量变化的气象因素、生物因素以及土壤特性进行了研究,认为GEP,RE与年均温(MAT)呈线性关系,与年均降水量(MAP)呈指数关系[15]。Law等研究了影响欧洲和美洲不同生态系统碳通量时空变化的因子,认为GEP与MAT间具有线性正相关关系,RE与MAT间具有指数相关关系[13]。Soussana等对欧洲9个草地生态系统碳通量进行了研究,认为影响草地间碳通量变异的因子主要是温度[16]。Hirata等阐释了东亚地区森林生态系统碳通量空间变异性的影响因素,认为GEP与MAT呈线性相关,RE与MAT呈指数相关关系[14]。碳通量的空间变化及其影响因素在不同的区域或植被类型间存在差异,而东亚地区森林和草地作为全球植被的重要部分,分析其碳通量的变异特征及其影响因素有助于对该地区以及全球碳汇的准确模拟以及管理。
本研究整合基于涡度相关法观测碳通量的文献,共选取东亚地区40个拥有1 a以上数据的通量站,其中森林站26个、草地站14个,以期进行以下研究:(1) 初步探明森林和草地生态系统CUE的空间变化及其影响因素。(2) 分析NEP,GEP,RE空间变化的影响因素。
1 材料与方法
1.1 通量数据收集
本研究采用的通量数据均是基于涡度相关法观测所得,数据来源于2003—2016年发表的文献。剔除异常降雨或异常高温(远高或低于多年平均降水及温度)的观测年份,最终筛选出40个观测时长大于或等于1 a的森林和草地的通量站。研究采用2000—2014年的105个有效站点年通量数据(表1),包括五个国家,中国(27个)、日本(8个)、蒙古(2个)、韩国(1个)、马来西亚(2个),其中森林站26个,草地站14个。
本研究所选取的通量站纬度范围2.97°—51.78°N,经度范围87.93°—142.10°E,气候带分布包括亚寒带、温带、亚热带以及热带。森林站包括落叶阔叶林(DBF)6个、落叶针叶林(DNF)5个、常绿阔叶林(EBF)6个、常绿针叶林(ENF)3个、针阔混交林(MF)6个,70%的森林为天然林,其中日本的Fujiyoshida站及Sapporo站为次生林。草地站包括高寒草甸4个、草甸草原2个、典型草原7个、荒漠草原1个。
1.2 气象、植被数据收集
本研究所需的气象数据主要为空气温度和降水量(表1),数据来源于与通量数据相同的文献,通过计算观测年的空气温度和降水量的平均值作为年均温(MAT)及年均降水量(MAP)。有5个站点(尖峰岭、西双版纳、黄土高原、阿柔、Sapporo)的空气温度及降水量数据在文献中没有详细列出,对于缺失的数据,本研究使用亚洲通量网(AsiaFlux,http:∥asiaflux.net/)和中国通量网(ChinaFLUX,http:∥www.chinaflux.org/)公布的多年平均空气温度及平均降水量作为研究数据。
本研究使用的生物数据为林龄和EVI(表1),获得林龄数据22个,70%的数据来源于已发表的文献,30%来源于亚洲通量网(AsiaFlux,http:∥asiaflux.net/)公布的站点概况。EVI是对归一化植被指数(NDVI)的修正,可以更好地反映植被生长状况。在本研究中,我们通过美国俄克拉荷马州地球观测与模拟研究中心(EOMF,http:∥www.eomf.ou.edu/)公布的MODIS数据集(MOD13 Q1)所提供的16 d,250 m分辨率的数据,获得每个研究站点观测年的EVI数据,其中日本Akou站2001—2003年的EVI数据缺失,在研究中将其剔除。

表1 各站点基本信息
1.3 数据可靠性及计算方法
1.3.1 数据可靠性 由于不同研究者对通量数据处理方法的差异,观测时间的差异以及观测仪器的差异导致的研究结果不同,因此进行研究前需确定数据的可靠性。当前,国际上通用的通量观测系统主要是开路式涡度观测系统(OPEC)和闭路式涡度观测系统(CPEC),在本研究中85%的数据通过OPEC系统测得,我国的通量站中只有老山通量站使用的是CPEC系统,而日本的通量站则习惯两种系统同时使用,本研究中其他国家的数据都是使用OPEC测得。Baldocchi等的研究认为开、闭路系统所测结果差异性不大,误差不高于5%[12]。本研究中采用的数据计算时长除我国的三江源站和海北站是15 min外,其他都是30 min,采样频率除日本的Akou(8 Hz)、Fujiyoshida(5 Hz)、Takayama(5 Hz)外,其余都是10 Hz,而且各研究观测年也相对集中(图1)。

图1东亚森林、草地通量数据观测时间分布
1.3.2 通量数据计算 本研究采用的数据中有80%的研究采用了夜间数据拆分法[46],通过两次或三次坐标轴旋转使平均垂直风速以及侧风速为零[47],之后利用WPL公式消除压力、温度、湿度等水热条件发生变化对通量观测带来的影响[48]。森林生态系统还需要进行储存项计算,用以消除冠层储存CO2的影响,草地生态系统不需要此计算(图2)。通过以上过程得到初步的净生态系统生产力(NEP),之后还需要剔除异常数据值,本研究参考文献的摩擦风速u*值在0.1 m/s到0.4 m/s,符合Reichstein等公布的适宜范围0.1~0.4 m/s[46]。参考文献所使用的数据插补法主要有平均日变化法、非线性回归法以及查表法等,Falge等的研究认为不同的插补法对结果影响不大,因此也可以认为通过以上方法计算所得的NEP具有可靠性[49]。然后,再通过夜间NEP建立的函数方程[46]估算出生态系统呼吸(RE),进而计算生态系统初级生产力(GEP)。
1.3.3 碳利用效率计算 一般认为生态系统碳利用效率(CUE)是表征生态系统固碳能力的重要参数,其计算公式如下[6-7]:
式中:NPP为净初级生产力;GPP为总初级生产力;Ra为植被自养呼吸。
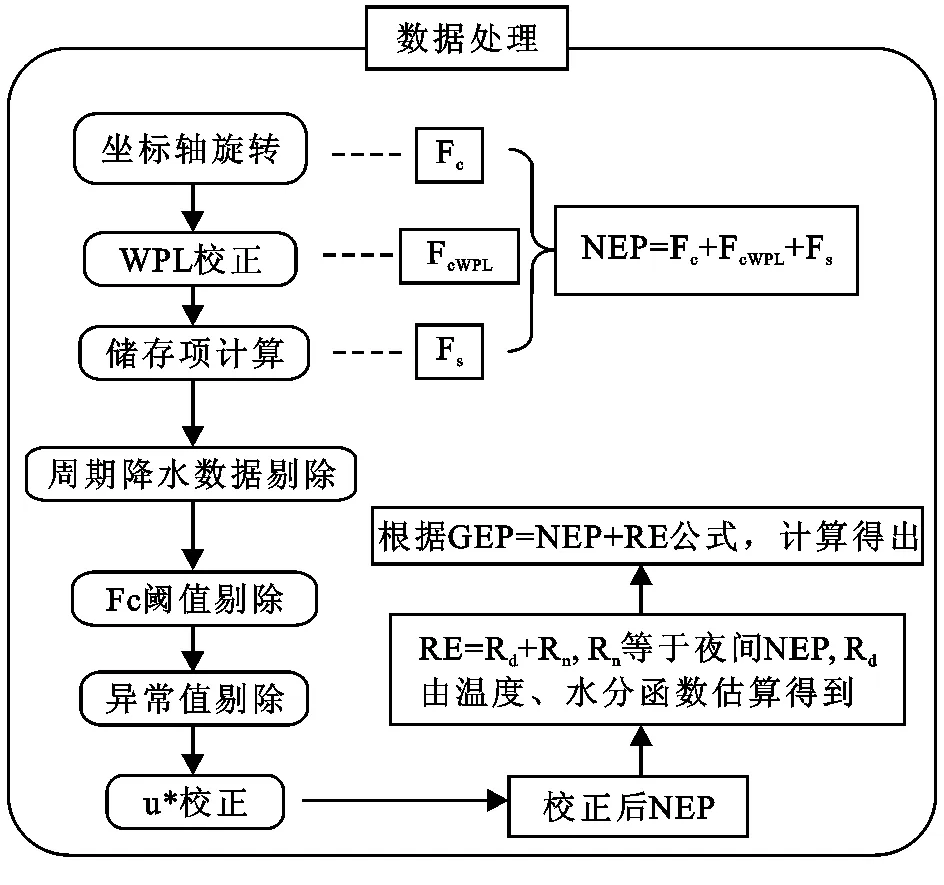
图2数据处理流程
1.3.4 统计分析 本研究利用SPSS 17.0对数据进行分析,其中单因素方差分析用于检验碳通量各组分间的差异性,Pearson相关分析用于检验气象、植被因素与CUE、碳通量各组分间的相关性。
2 结果与分析
2.1 东亚森林、草地生态系统的碳通量
东亚地区森林以及草地生态系统均呈现碳汇格局,其中森林生态系统呈现出较强的碳吸收能力,NEP,GEP,RE分别为328.64±256.46 gC/(m2·a),1 659.22±685.21 gC/(m2·a),1 330.58±664.62 gC/(m2·a)。草地的碳吸收能力较弱,其NEP,GEP,RE分别为63.43±42.99 gC/(m2·a),519.98±568.66 gC/(m2·a),456.55±574.54 gC/(m2·a)。森林的NEP,GEP,RE显著高于草地(p<0.001)(图3),GEP,RE是草地的3倍,NEP则可达到5倍。森林和草地的NEP具有较大差异的主要原因是森林的光合作用强度要远高于草地[50],而呼吸比草地更稳定,且草地更易受到外界扰动从而加剧碳的排放,而GEP,RE差异大的原因主要是植被类型的差异,森林的冠层导度更大,叶绿体含量更高,因此其吸收及释放碳的能力也更强[51-52]。
2.2 碳利用效率(CUE)及其空间格局
研究发现森林和草地的CUE无显著性差异(p=0.82),CUE的平均值分别为0.21,0.20。其中森林CUE最高的站点为浙江天目山柳杉、银杏混交林观测站,达到0.44,最低的为马来西亚的Lambir热带雨林观测站,只有0.002 9。草地CUE最高的站点为黄土高原草地站,达到0.59,最低的为日本的TERC草地站,仅有0.007 3。陈智认为北半球陆地生态系统呼吸光合比为0.87,与本研究中发现森林、草地固定的碳有80%被呼吸消耗的结果相似[37]。Kato[14]、Hirata[17]等的研究据发现森林和草地的CUE变化范围为0.20~0.30。不同的研究方法对于CUE的估计存在差异,DeLucia等认为不同的森林类型CUE的范围可以达到0.20~0.80[8],Zhang等利用MODIS影像对CUE的研究认为草地的CUE高于森林[10],但是,在本研究中并未发现两种生态系统CUE的差异,这主要是因为不同的森林类型、草地种类的CUE之间也存在差异,仅以大类划分,简单平均导致差异不显著。

注:字母a,b表示两个数值间在p<0.001水平上差异显著。
图3森林和草地间NEP,GEP,RE的差异
本研究发现东亚地区的森林、草地CUE纬向分布特点不显著(图4),这也与Zhang等的研究结果相似[10],虽然没有显著的分布特点,但可以发现较高的CUE主要分布在中纬度地区。

图4CUE与纬度的关系
2.3 碳利用效率(CUE)变化的影响因素
本研究发现森林CUE与林龄具有极显著的线性负相关关系(图5)。一般认为林龄高的森林对环境变化的响应不敏感,随着林龄的增加,森林对光、水、养分的利用效率降低,光合能力下降,但是呼吸增强,从而导致CUE下降[7,9]。本研究未发现森林CUE与MAT和MAP之间的相关关系。草地CUE与MAP间存在显著的线性负相关关系(图6),这与Zhang等对全球生态系统的研究一致[10],森林CUE与MAP无相关关系,可能是因为大部分森林站点分布的地区降水充沛,对CUE的调控作用较弱。而草地站大部分分布在北部降水较少的地区,对降水的响应相对敏感[8]。本研究中的东亚地区NEP随MAP增加先升高后降低,GEP随MAP增加而升高,因而其比值会呈现出随MAP增加而下降的趋势。

图5森林生态系统CUE与林龄的关系

图6草地生态系统CUE与MAP的关系
2.4 NEP,GEP,RE的空间变化特征
虽然东亚地区森林和草地的CUE纬向变化特征不显著,但NEP,GEP,RE均呈现显著的纬向分布特征(图7),森林站点纬度范围为2.97°—51.78°N,NEP先随纬度升高而增加,当纬度接近30°N时开始随纬度升高而降低。草地站点主要集中在30°—50°N,NEP先随纬度升高而增加,当纬度达到40°N左右时开始随纬度升高而降低。而GEP,RE均随纬度的升高而线性降低。
NEP的空间变化受到多种因素的影响,低纬度时GEP较高,但是RE也高,随着纬度的升高RE减弱,GEP也在降低,在中纬度NEP达到最大值,因此NEP受到GEP和RE的制约在纬向上呈现二次函数关系。陈智对北半球陆地生态系统的研究表明GEP,RE与纬度呈负相关关系,但是认为NEP不存在纬向变异特征[37],而王兴昌等对全球森林的研究则认为GEP与纬度具有线性负相关关系,NEP,RE均与纬度呈二次函数关系[53]。陆地生态系统的纬向变异特征在不同的研究中有很大差异,这是由植被类型差异、环境差异、研究站点选取差异以及观测年份差异等影响造成的。

图7森林、草地生态系统NEP,GEP,RE的纬向分布特点
2.5 NEP,GEP,RE变化的影响因素
2.5.1 降 水 森林和草地的NEP,GEP,RE具有纬向分布特征,而不同的纬度往往具有不同的植被、气象特征,因此NEP,GEP,RE也受到生物或环境因素的影响[5]。本研究中,森林和草地的GEP,RE与MAP呈现正相关关系(图8),随MAP增加而线性增加,这与Kato[17]、王兴昌[53]等的研究相一致。NEP与MAP呈二次函数关系,随着MAP增加,NEP先升高后降低,与王兴昌等的研究结果相似[53],其中森林和草地的饱和降水量约为1 300 mm和390 mm,GEP,RE均随MAP的增加而增加,但是RE-MAP的斜率大于GEP-MAP,即RE随MAP增加而升高的速率更高,因此当降水量达到一定值时NEP达到饱和。目前对MAP与NEP关系的研究存在很大差异,Hirata等认为森林的NEP与MAP无相关关系[14],Luyssaert等也认为NEP不受环境因素的影响[54],而Chen等的研究则认为NEP受到MAP,MAT的共同调控[50],因此MAP对NEP的调控机制还有待于进一步深入研究。
2.5.2 空气温度 与降水量相似,空气温度也被认为是影响碳通量变化的重要因子之一[17]。本研究发现森林的GEP,RE与MAT呈正相关关系(图9),随温度升高而增加,这与Kato[14]、Hirata[17]等及王兴昌[53]等的研究结果相似。NEP与MAT有呈二次函数关系的趋势,但是相关性并不显著,Hirata[14]等和王兴昌[53]等认为NEP与MAT具有二次函数关系。另外,本研究还发现草地的碳通量各组分与MAT间无显著相关性,可能是由于本研究中的草地站点偏少,且主要集中在低温地区,温度差异不明显。
2.5.3 增强型植被指数(EVI) EVI可以较好地反映植被的生长状况,而植被的生长状况对于生态系统的固碳能力有重要的影响[13]。本研究发现,森林和草地的GEP,RE与EVI呈线性正相关关系(图10),这与Chen等[15]的研究相一致。从研究中可以看出植被生长状况对植被的光合作用、呼吸作用有重要的影响,植被生长状况越良好,生态系统的GEP,RE越高。研究未发现NEP与EVI的相关关系。

图8 森林、草地生态系统的NEP,GEP,RE与MAP的关系

图9 森林、草地生态系统的NEP,GEP,RE与MAT的关系

图10 森林、草地生态系统的NEP,GEP,RE与EVI的关系
3 结 论
本研究整合基于涡度相关法观测碳通量的已发表文献,共选取东亚地区40个拥有1 a以上数据的通量站,包括森林站26个,草地站14个。初步探明了森林和草地生态系统CUE的空间变化及其影响因素,并分析了其NEP,GEP,RE空间变化的影响因素。研究发现森林的NEP,GEP,RE分别为328.64±256.46 gC/(m2·a),1 659.22±685.21 gC/(m2·a),1 330.58±664.62 gC/(m2·a),草地的NEP,GEP,RE分别为63.43±42.99 gC/(m2·a),519.98±568.66 gC/(m2·a),456.55±574.54 gC/(m2·a),森林的碳通量各组分显著高于草地。森林和草地的CUE分别为0.21,0.20,两者差异不显著,纬向分布特征不显著。本研究发现林龄是影响森林CUE变化的主要因素,呈线性负相关关系。MAP是影响草地CUE的主要因素,同样呈线性负相关关系。
森林和草地的GEP,RE均随纬度的升高而降低,但GEP随纬度的变化速率大于RE。森林和草地的GEP,RE与MAP呈线性正相关关系,NEP与MAP呈先升高后降低的二次函数关系,其中森林和草地的饱和降水量大约为1 300 mm和390 mm。森林的GEP,RE与MAT呈现线性正相关关系,NEP与MAT有二次函数关系趋势,但是相关性不显著。草地的GEP,RE,NEP与MAT无显著的相关关系。森林和草地的GEP,RE与EVI呈线性正相关关系,NEP与EVI无显著相关关系。
[1] Keeling C D, Bacastow R B, Carter A F, et al. A three dimensional model of atmospheric CO2transport based on observed winds: observation data and preliminary analysis: Aspects of climate variability in the Pacific and the Western Americas[J]. American Geophysical Union,1989,55:165-236.
[2] 于贵瑞,伏玉玲,孙晓敏,等.中国陆地生态系统通量观测研究网络(ChinaFLUX)的研究进展及其发展思路[J].中国科学D辑:地球科学,2006,36(SΙ):1-21.
[3] Ralph M. Global forest resources assessment(FAO). (2010). www. fao. org/forestry/en.
[4] Scurlock J M O, Hall D O. The global carbon sink: a grassland perspective[J]. Global Change Biology, 1998,4(2):229-233.
[5] Yu G R, Zhu X J, Fu Y L, et al. Spatial pattern and climate drivers of carbonfluxes in terrestrial ecosystems of China[J]. Global Change Biology, 2013,19(3):798-810.
[6] Ryan M G, Lavigne M B, Gower S T. Annual carbon cost of autotrophic respiration in boreal forest ecosystems in relation to species and climate[J]. Journal of Geophysical Research-Atmospheres, 1997,102:28871-28883.
[7] Chambers J Q, Tribuzy E S, Toledo L C, et al. Respiration from a tropical forest ecosystem: partitioning of sources and low carbon use efficiency[J]. Ecological Applications, 2004,14(sp4):72-88.
[8] DeLucia E, Drake J E, Thomas R B, et al. Forest carbon use efficiency: Is respiration a constant fraction of gross primary production[J]. Global Change Biology, 2007,13(6):1157-1167.
[9] Piao S, Luyssaert S, Ciais P, et al. Forest annual carbon cost: A global-scale analysis of autotrophic respiration[J]. Ecology, 2010,91(3):652-661.
[10] Zhang Y, Xu M, Chen H, et al. Global pattern of NPP to GPP ratio derived from MODIS data: effects of ecosystem type, geographical location and climate[J]. Global Ecology and Biogeography, 2009,18(3):280-290.
[11] 朱万泽.森林碳利用效率研究进展[J].植物生态学报,2013,37(11):1043-1058.
[12] Baldocchi D, Falge E, Gu L H, et al. Fluxnet:a new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities[J]. Bulletin of the American Meteorological Society, 2001,82(11):2415-2434.
[13] Law B E, Falge E, Gu L, et al. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation[J]. Agricultural and Forest Meteorology, 2002,113(1):97-120.
[14] Hirata R, Saigusa N, Yamamoto S, et al. Spatial distribution of carbon balance in forest ecosystems across East Asia[J]. Agricultural and Forest Meteorology, 2008,148(5):761-775.
[15] Chen Z, Yu G, Ge J, et al. Roles of climate, vegetation and soil in regulating the spatial variations in ecosystem carbon dioxide fluxes in the Northern Hemisphere[J]. PloS ONE, 2015, 10(4):1-14.
[16] Soussana J F, Allard V, Pilegaard K, et al. Full accounting of the greenhouse gas (CO2, N2O, CH4) budget of nine European grassland sites[J]. Agriculture, Ecosystems & Environment, 2007,121(1):121-134.
[17] Kato T, Tang Y. Spatial variability and major controlling factors of CO2sink strength in Asian terrestrial ecosystems: Evidence from eddy covariance data[J]. Global Change Biology, 2008,14(10):2333-2348.
[18] 陈德祥.尖峰岭热带山地雨林碳交换的动态特征和影响因素研究[D].北京:中国林业科学研究院,2010.
[19] 吴志祥,谢贵水,杨川,等.海南儋州地区橡胶林碳通量特征研究[J].西北林学院学报,2015,30(1):51-59.
[20] Zhang Y P, Tan Z H, Song Q H, et al. Respiration controls the unexpected seasonal pattern of carbon flux in an Asian tropical rain forest[J]. Atmospheric Environment, 2010,44(32):3886-3893.
[21] 王春林,于贵瑞,周国逸,等.鼎湖山常绿阔叶混交林CO2通量估算[J].中国科学D辑:地球科学,2006,36(S):119-129.
[22] 唐亚坤.中亚热带人工针叶林2003-2012年碳水通量年际变异的控制机制研究[D].北京:中国科学院大学,2014.
[23] 张利平.会同杉木人工林生态系统与大气间CO2通量特征的研究[D].长沙:中南林业科技大学,2010.
[24] 牛晓栋,江洪,张金梦,等.浙江天目山老龄森林生态系统CO2通量特征[J].应用生态学报,2016,27(1):1-8.
[25] 杨爽.浙江安吉毛竹林生态系统CO2通量观测研究[D].杭州:浙江农林大学,2012.
[26] 朱志鹍,马耀明,胡泽勇,等.青藏高原那曲高寒草甸生态系统CO2净交换及其影响因子[J].高原气象,2015,34(5):1217-1223.
[27] 蒋琰.北亚热带次生栎林碳收支的研究[D].南京:南京林业大学,2010.
[28] 耿绍波.河南西平杨树人工林生态系统碳通量及其环境相应研究[D].北京:北京林业大学,2011.
[29] 吴力博,古松,赵亮,等.三江源地区人工草地的生态系统CO2净交换、总初级生产力及其影响因子[J].植物生态学报,2010,34(7):770-780.
[30] Huang H, Zhang JS, Meng P, Fu YL, Zheng N, et al. Seasonal Variation and Meteorological Control of CO2Flux in a Hilly Plantation in the Mountain Areas of North China[J]. Acta Meteorologica Sinica, 2011,25(2):238-248.
[31] Du Q, Liu H Z, Feng J W, et al. Carbon dioxide exchange processes over the grassland ecosystems in semiarid areas of China[J]. Science China-Earth Science, 2012,42(5):711-722.
[32] 张法伟,李英年,曹广民,等.青海湖北岸高寒草甸草原生态系统CO2通量特征及其驱动因子[J].植物生态学报,2012,36(3):187-198.
[33] Kwon H, Kim J, Hong J, et al. Influence of the Asian monsoon on net ecosystem carbon exchange in two major ecosystems in Korea[J]. Biogeosciences, 2010,7(5):1493-1504.
[34] 王海波,马明国,王旭峰,等.青藏高原东缘高寒草甸生态系统碳通量变化特征及其影响因素[J].干旱区资源与环境,2014,28(6):50-56.
[35] 王杰,叶柏生,张世强,等.祁连山疏勒河上游高寒草甸CO2通量变化特征[J].冰川冻土,2011,33(3):646-653.
[36] 方显瑞.杨树人工林生态系统碳交换及其环境响应[D].北京:北京林业大学,2011.
[37] 陈智.北半球陆地生态系统碳交换通量的空间格局及其调控机制研究[D].北京:中国科学院大学,2015.
[38] 张文丽.内蒙古农牧交错区典型草地和农田生态系统碳通量的研究[D].北京:中国科学院植物研究所,2007.
[39] 张军辉,于贵瑞,韩士杰,等.长白山阔叶红松林CO2通量季节和年际变化特征及控制机制[J].中国科学D辑:地球科学.2006,36(S):60-69.
[40] Nakai Y, Kitamura K, Suzuki S, et al. Year-long carbon dioxide exchange above a broadleaf deciduous forest in Sapporo, Northern Japan[J]. Tellus B, 2003,55(2):305-312.
[41] 刘冉,李彦,王勤学,等.盐生荒漠生态系统二氧化碳通量的年内,年际变异特征[J].中国沙漠,2011,31(1):108-114.
[42] 董刚.中国东北松嫩草甸草原碳水通量及水分利用效率研究[D].长春:东北师范大学,2011.
[43] Wang Y L, Zhou G S, Wang Y H. Environmental effects on net ecosystem CO2exchange athalf-hour and month scales over Stipakrylovii steppe in northern China[J]. Agricultural and Forest Meteorology, 2008,148(5):714-722.
[44] Li S G, Asanuma J, Eugster W, et al. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia[J]. Global Change Biology,2005,11:1941-1955.
[45] 周艳丽,贾丙瑞,周广胜,等.中国北方针叶林生长季碳交换及其调控机制[J].应用生态学报,2010,21(10):2449-2456.
[46] Reichstein M, Falge E, Baldocchi D, et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration:review and improved algorithm[J]. Global Change Biology, 2005,11(9):1424-1439.
[47] Wilczark J M, Oncley S P, Stage S A. Sonic anemometer tilt correction algorithms[J]. Boundary Layer Meteorology.2001,99:127-150.
[48] Webb E K, Pearman G I, Leuning R. Correction of flux measurements for density effects due to heat and water vapour transfer[J]. Quarterly Journal of the Royal Meteorological Society, 1980,106(447):85-100.
[49] Falge E, Baldocchi D, Olson R et al. Gap filling strategies for defensible annual sums of net ecosystem exchange[J]. Agricultural and Forest Meteorology, 2001,107(1):43-69.
[50] Chen Z, Yu G R, Ge J P, et al. Temperature and precipitation control of the spatial variation of terrestrial ecosystem carbon exchange in the Asian region[J]. Agricultural and Forest Meteorology, 2013,182/183:266-276.
[51] Wang B, Li J, Jiang W W, et al. Impacts of the rangeland degradation on CO2flux and the underlying mechanisms in the Three-River Source Region on the Qinghai-Tibetan Plateau[J]. China Environmental Science, 2012,32(10):1764-1771.
[52] 於方,朱文泉,曹东,等.青海省因草地生态破坏造成土壤流失的经济损失核算[J].中国环境科学,2009,29(1):90-94.
[53] 王兴昌,王传宽,于贵瑞.基于全球涡度相关的森林碳交换的时空格局[J].中国科学:D辑,2008,38(9):1092-1102.
[54] Luyssaert S, Inglima I, Jung M. CO2balance of boreal, temperate, and tropical forests derived from a global database[J]. Global Change Biology, 2007,13(12):2509-2537.
FactorsAffectingtheSpatialVariationofCarbonUseEfficiencyandCarbonFluxesinEastAsianForestandGrassland
AN Xiang1, CHEN Yunming2,3, TANG Yakun2,3
(1.InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling,Shaanxi712100,China;2.StateKeyLaboratoryofSoilErosionandDrylandFarmingontheLoessPlateau,InstituteofSoilandWaterConservation,NorthwestA&FUniversity,Yangling,Shaanxi712100,China; 3.InstituteofSoilandWaterConservation,ChineseAcademyofSciencesandMinistryofWaterResources,Yangling,Shaanxi712100,China)
Forest and grassland play an essential role in the terrestrial ecosystem. The objective of this study is to analyze the spatial variation characteristics and influence factors of carbon use efficiency (CUE), gross ecosystem production (GEP), ecosystem respiration (ER), and net ecosystem production (NEP) based on more than one year data which were obtained from 40 sites (26 forest sites, 14 grassland sites) in the East Asian region from published literatures. Forest and grassland in the East Asian region both exhibit carbon sinks, while forest showed significantly higher value than grassland (p<0.001). The average NEP in forest and grassland in the East Asian are 328.64±256.46 gC/(m2·a) and 63.43±42.99 gC/(m2·a), respectively. The average CUE in forest and grassland are 0.21 and 0.20 in this region, CUE of forest is negatively correlated with the forest age and mean annual precipitation (MAP), respectively. The results show that GEP and ER in forest grassland linearly decrease with an increase in latitude. In addition, relationship between NEP and latitude can be described by quadratic function. There is a clear linear relationship between GEP, ER with MAP in both forest and grassland. The results also show that MAP and NEP first increase and then decrease with a quadratic function relationship, NEP is the maximum when amounts precipitation are 1 300 mm and 390 mm for forest and grassland, respectively. GEP and ER in forest show a positive relationship with mean annual temperature. Moreover, GEP and RE in both forest and grassland showed a positive linear correlation with vegetation index.
forest; grassland; CUE; carbon dioxide flux; eddy covariance
2016-09-29
:2016-10-24
西北农林科技大学博士科研启动费(Z109021508);中央高校基本科研业务费(Z109021606);公益性行业(气象)科研专项(重大专项)(GYHY201506001-3)
安相(1990—),男(锡伯族),辽宁抚顺人,硕士研究生,从事陆地生态系统碳汇相关研究。E-mail:xiangan10@163.com
陈云明(1967—),男,陕西渭南人,博士,研究员,主要从事植被与水土保持的相关研究。E-mailymchen@ms.iswc.ac.cn
S718.5;X171.1
:A
:1005-3409(2017)05-0079-09
