克隆整合有利于喜旱莲子草(Alternantheraphiloxeroides)入侵
2017-09-08方龙香吕晓倩奚道国游文华依成武杜道林
方龙香,吕晓倩,奚道国,游文华,依成武,杜道林
(1:江苏大学环境与安全工程学院,镇江 212013)(2:合肥工业大学建筑与艺术学院,合肥 230009)
克隆整合有利于喜旱莲子草(Alternantheraphiloxeroides)入侵
方龙香1,吕晓倩2,奚道国1,游文华1,依成武1,杜道林1
(1:江苏大学环境与安全工程学院,镇江 212013)(2:合肥工业大学建筑与艺术学院,合肥 230009)
多数恶性入侵水生植物都是克隆植物,阐明克隆整合属性对其潜在入侵性的影响对了解其入侵机制具有重要意义. 以喜旱莲子草(Alternantheraphiloxeroides)为研究对象,研究其克隆整合(匍匐茎连接)对入侵本地黄花水龙(Ludwigiapeploides)种群时的光合性能、生长、竞争力和资源分配的影响,探讨克隆整合对该植物入侵性的贡献. 结果表明,竞争显著降低了喜旱莲子草的光系统Ⅱ最大量子产量、总生物量、分株数和总匍匐茎长;但克隆整合却显著促进了竞争环境中喜旱莲子草的生长,提高了其光合特性、根冠比以及对本地物种的竞争力,因而有利于其入侵性的增强.
入侵植物;生理整合;竞争力;根冠比;光合表现;喜旱莲子草
外来植物入侵对全球和区域生物多样性和生态系统构成了巨大的威胁[1-2]. 在水生生态系统中,多数成功入侵的外来植物均为具有非常强克隆繁殖能力的克隆植物. 例如,几种恶性入侵水生植物凤眼莲(Eichhorniacrassipes)、喜旱莲子草(Alternantheraphiloxeroides)和粉绿狐尾藻(Myriophyllumaquaticum)等能在水中疯狂克隆繁殖形成致密的毡层,排挤水生生态系统中几乎其他所有植物[3-5]. 目前,入侵植物克隆性状已逐渐成为当前克隆植物生态学和入侵生态学的研究热点之一[6-7].
克隆植物的成功入侵与其独特的克隆生活史性状密切相关,而克隆整合(clonal integration)是这些克隆性状中最为突出的一个[7]. 克隆整合是克隆生理整合的简称,是指由源-汇关系驱动的,克隆植物分株之间具有共享水分、碳水化合物和营养物质等的能力[8-9]. 克隆整合被证实能帮助植物分株在胁迫环境下拓殖和生长,在剧烈的环境变化下生存和快速恢复,并占据新的空白生境;能增强分株的竞争能力,进而有利于其入侵周边群落[10-13]. 克隆整合的这些积极作用能帮助克隆植物比非克隆植物和克隆能力比较弱的植物表现出更好的性能. 因此,外来植物的入侵性可能和克隆植物的克隆整合特性密切相关[7,14]. 尽管如此,目前多数对入侵克隆植物的研究常忽视克隆整合属性的潜在影响,这可能会导致对这些入侵植物的入侵性缺乏全面的认识,特别是关于克隆整合是否会有利于外来植物入侵本地植物群落的认识尚不统一[15-16].
本实验以入侵水生植物喜旱莲子草为研究对象,采用温室控制实验的方法,研究克隆整合(匍匐茎连接)对喜旱莲子草的光合性能、生长和竞争力的影响,以及喜旱莲子草克隆整合对本地植物群落的影响,以期从植物克隆性状方面深入了解入侵水生植物的入侵机制,并为这些植物的防控提供理论依据.
1 材料和方法
1.1 实验材料
喜旱莲子草又名空心莲子草、水花生和革命草等,起源于南美的巴拿马运河流域[17]. 为多年生草本植物,具有匍匐茎和根状茎,生长在池沼、水沟等地,为水陆两栖植物. 自1930年作为饲料引入我国江浙沪皖等地区,被广泛种植,后逸为野生[17-18]. 因其具有极强的克隆繁殖能力,且能适应各种外界环境,已成为危害性极大的入侵物种,为我国环境保护部首批公开的16种恶性入侵物种之一[17-18]. 近年来国内外研究发现,克隆整合是决定喜旱莲子草生长、克隆繁殖和建群的主要因素之一[11,13,15,19-21].
选取的竞争背景乡土植物为黄花水龙(Ludwigiapeploides),是多年生挺水草本植物,具有匍匐茎,克隆繁殖能力很强,主要分布于我国的中部和南部. 在湿地环境中,常与喜旱莲子草共存,易形成竞争关系[16,20].
以上两种植物材料均采自于太湖贡湖湾(31°25′~31°28′N, 120°15′~120°21′E),材料均为生长良好无虫害的植株顶端部分,带回温室培养用于实验.
1.2 实验设计
本实验在江苏大学环境与安全工程学院的温室中进行,实验期间温室的昼夜平均温度分别为25±1℃和 15±1℃,光照强度为90%的自然光照(700~1200 μmol/(m2·s)),昼夜周期为14 h∶ 10 h. 实验处理为匍匐茎连接(克隆整合)和植物竞争的两因素析因设计(图1). 将采集的喜旱莲子草断枝置于河沙基质中预培养1周后,从中选取24个大小相当的喜旱莲子草克隆片段(每个克隆片段含4个相连分株,长度为13.25±0.20 cm)用于实验. 其中,每个喜旱莲子草克隆片段被分成两部分:2个靠近基部相对较老的分株称作“基部分株”,2个远离基部靠近端部相对较嫩的分株称作 “端部分株”. 实验的容器为24个白色塑料筐(长×宽×高=50 cm×40 cm×15 cm),每个塑料筐亦被分为两部分,即长度为20 cm的基部和长度为30 cm的端部(图1). 通过塑料膜的分别包裹使容器的两部分之间的资源(水分和营养)以及植株根系互不干扰. 实验容器内的基质为洗净的沙子和湖泥的混合物(体积比为1∶1)以及5 g缓释肥(Osmocote, 其元素比例N∶P∶K=16∶8∶12),厚度约15 cm.
2014年5月5日,依据在自然湿地中生长的黄花水龙密度,分别在其中的12个实验容器的端部种植20株黄花水龙的克隆片段(长度为15.12±0.25 cm),用于形成种群来设置竞争环境. 约4周以后,待黄花水龙种群生长稳定之后,将挑选的24个喜旱莲子草的克隆片段匍匐放置于实验容器基质上(穿过塑料膜),每个容器1棵植株,植株的端部分株放置在容器端部中间,相对应的基部分株放置于容器的基部(图1). 再经过1周的适应性培养,待喜旱莲子草植株在新环境重新扎根后开始实验处理:将12个容器中的喜旱莲子草端部分株和基部分株相连的匍匐茎用剪刀切断以阻断克隆整合,另外12个容器的植株匍匐茎保持连接以检验克隆整合作用(图1). 实验期间每天浇水,保持各处理的基质上覆盖2 cm深的水以模拟湿地环境. 8周以后,实验进行收获.

图1 实验设计示意图(每个容器中均为1个喜旱莲子草的克隆片段,容器左侧为2个基部分株,右侧为2个端部分株;基部和端部分株切断(上)或保持连接(下),端部分株处于空白生境或植物竞争生境)Fig.1 Schematic representation of the experimental design (There was one clonal fragment of Alternanthera philoxeroides in each container, consisting of two basal ramets in the left side and two apical ramets in the right side; Stolon between basal and apical ramets was disconnected (upside) or connected (underside), with the apical ramets growing in the open habitats or vegetative habitats)
1.3 数据收集
实验结束的前1周,在实验处理的每个端部分株中选取次嫩分株上的1片完全张开且长势良好的叶片,先用叶夹进行至少30 min的暗适应以确保光系统II(PSII)反应中心完全打开,然后再用便携式叶绿素荧光测定仪(型号为PAM-2100, Walz, Effeltrich, Germany)以饱和脉冲法[22]测定其最小荧光产量(F0)和最大荧光产量(Fm). 光系统II最大量子产量(Fv/Fm)=(Fm-F0)/Fm.
实验待收获时,记录各个处理端部分株的分株数,并测量总的匍匐茎长. 然后将所有处理分株小心收获洗净并分成叶、匍匐茎和根3个部分,并将各个部分标记后放入70℃的烘箱中烘72 h至恒重后测量干重. 此外,容器端部的黄花水龙也一并收获(地上部分),并且以同样的方法测量干重.
用相对邻体效应指数(relative neighbor effect,RNE)来衡量黄花水龙对喜旱莲子草的竞争强度,其表达公式为RNE=(C-A)/max(C,A),其中A代表无竞争时喜旱莲子草的生物量,C代表有竞争时喜旱莲子草的生物量,max(C,A)代表两者中的较大值. 相对邻体指数的数值范围为-1~0,值越大表示邻体效应越小[11,15]. 若匍匐茎连接时的RNE值大于匍匐茎切断时的值,则表示克隆整合提升了喜旱莲子草的竞争能力[11,15-16].
1.4 数据分析
数值用平均值±标准误表示. 数据分析前均经过对数转换以确保满足正态分布和方差的齐次性. 利用双因素方差分析(Two-way ANOVA)检验克隆整合(匍匐茎连接)和植物竞争对端部分株的光合性能(Fv/Fm)、生长参数(生物量、分株数和匍匐茎总长)和根冠比的影响. 若处理的影响显著,再利用Studentized Tukey’s HSD对因素内不同水平间的影响进行多重比较. 用t-test检验法检验喜旱莲子草的RNE在匍匐茎连接和切断时的差异以及黄花水龙种群的生物量在有无喜旱莲子草存在时的差异. 数据分析采用SPSS 18.0统计分析软件,显著度水平设置为0.05.
表1 匍匐茎连接、竞争以及两者交互作用对喜旱莲子草端部分株生长指标影响的双因素方差分析
Tab.1 Two-way ANOVA analyses of the effects of stolon connection, competition and their interaction on the growth index of Alternanthera philoxeroides

指标匍匐茎连接竞争交互作用Fv/Fm143.07***11.91**15.29**总生物量24.39***90.75***3.73分株数7.86*52.99***2.10总匍匐茎长8.96**69.16***1.79根冠比2.240.5115.13**
*表示P< 0.05;**表示P< 0.01;***表示P< 0.001.
2 结果
2.1 喜旱莲子草的光合性能和生长
匍匐茎连接、竞争以及两者的交互作用显著影响了喜旱莲子草端部分株的Fv/Fm值(表1). 植物竞争显著降低了喜旱莲子草端部分株的Fv/Fm值(图2A). 在空白生境中,匍匐茎连接对端部分株的Fv/Fm值并没有显著影响;而在黄花水龙竞争的环境下,匍匐茎连接显著提升了喜旱莲子草端部分株的Fv/Fm值(图2A).
匍匐茎连接和竞争均显著影响喜旱莲子草端部分株的生长参数(总生物量、分株数和匍匐茎长),但二者的交互作用均不显著(表1). 竞争显著降低了喜旱莲子草端株部分的总生物量、分株数和总匍匐茎长(图2B~D). 在空白生境中,匍匐茎连接仅显著促进了端部分株的生物量积累(图2B),对其分株数和匍匐茎总长度的影响并不显著(图2C、D);在黄花水龙竞争的环境下,匍匐茎连接则显著提高了喜旱莲子草端株部分的生长指标,增幅达30%~65%(图2B~D).

图2 不同处理下喜旱莲子草端部分株的光化学最大量子产量(Fv/Fm)(A),总生物量(B),分株数(C)和匍匐茎长(D)(相同字母表示在P=0.05水平下处理间的差异不显著,下同) Fig.2 The maximum quantum yield of photosystem Ⅱ (A), total biomass (B), ramet number (C) and stolon length (D) of Alternanthera philoxeroides ramets in the apical part in different treatments (The bars sharing the same letter are not significantly different at P=0.05, the same below)
2.2 喜旱莲子草的竞争力和黄花水龙种群的生长
喜旱莲子草端部分株在匍匐茎连接时的RNE为-0.22±0.04,显著大于匍匐茎切断时的-0.40±0.05(t=4.56,P<0.05),即喜旱莲子草的竞争力在匍匐茎连接时显著大于切断时.
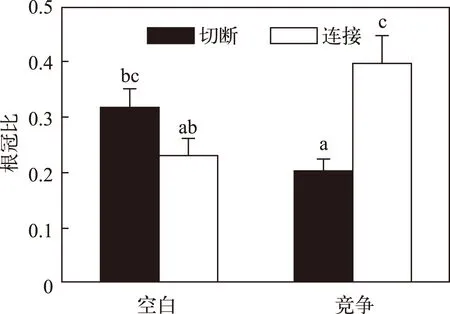
图3 不同处理下喜旱莲子草端部分株的根冠比 Fig.3 Root/shoot ratios of apical ramet of Alternanthera philoxeroides in different treatments
黄花水龙种群地上生物量在喜旱莲子草匍匐茎连接和切断时分别为72.35±4.80和77.18±5.72 g,匍匐茎连接对竞争植被黄花水龙种群的地上部分生物量没有显著影响(t=1.53,P=0.22).
2.3 喜旱莲子草的生物量分配
匍匐茎连接和竞争对喜旱莲子草端部分株的根冠比并没有显著影响,而两者的交互作用却显著影响了喜旱莲子草的根冠比(表1). 在无竞争时,相比于匍匐茎切断的分株,匍匐茎连接有降低其端部分株根冠比的趋势,但统计学上并不显著(图3);而在黄花水龙种群竞争环境下,相比于匍匐茎切断的分株,匍匐茎连接则显著升高了喜旱莲子草端部分株的根冠比(图3).
3 讨论
通常在良好的生长环境中,植物Fv/Fm值的正常范围为0.75~0.85[5,11,21],Fv/Fm值亦表示植物潜在的最大光合效率. 在无植被竞争环境中,喜旱莲子草端部分株的Fv/Fm值在相连分株匍匐茎切断和不切断的情况下都处于正常范围内且处理之间没有显著差异,表明植物没有受到环境胁迫. 而在黄花水龙植被竞争环境中,喜旱莲子草端部分株匍匐茎切断的情况下其Fv/Fm显著下降到正常范围之外,表明其分株受到强烈的种间竞争胁迫[11,16]. 然而,这种胁迫在分株匍匐茎相连的情况下(克隆整合)明显得到缓和,即其Fv/Fm值在克隆整合作用下显著上升并恢复到正常范围内,表明克隆整合能缓和植被竞争的胁迫且能显著提升分株的光合性能. 前人研究也发现克隆整合使生长在含有重金属土壤或受到其他物种强烈的竞争胁迫下的分株的Fv/Fm值下降[10,11,16]. 因此,克隆整合可缓和入侵种喜旱莲子草在入侵本地植被时所遭受的竞争胁迫作用,进而有助于增强其入侵性[11].
已有的许多研究发现,克隆整合对克隆植物的新生分株生长和生存都具有积极作用[5,11,15,20]. 研究发现在无黄花水龙竞争时,克隆整合虽然仅增加了喜旱莲子草的生物量,但当存在本地竞争植被时,克隆整合对喜旱莲子草的所有生长参数都有明显的促进作用. 这很可能是因为基株通过克隆整合作用向顶性(从基株到端株)传输碳水化合物等给相连的端株[11,21],表明克隆整合对喜旱莲子草新生分株的生长、扩散以及占据空白生境非常重要,进而有利于增强其在自然生境中的入侵性[11,21],尤其是喜旱莲子草的克隆整合可能在竞争环境中更为重要[13].
在植物群落中,为争夺共有空间和资源,共存种之间的种间竞争常常会导致竞争物种的此消彼长. 本实验中,相比无竞争时,黄花水龙的强烈种间竞争作用使喜旱莲子草的生长明显受到抑制:其端株部分总生物量下降了35%~49%,分株数下降了32%~44%,总匍匐茎长度下降了36%~41%. 表明黄花水龙种群的存在较强地抑制了喜旱莲子草的生长和扩散[15-16]. 在先前的研究中,研究者大多发现克隆整合对于克隆植物开拓空白生境、抵御环境胁迫和有效利用资源非常重要,但不会显著提高克隆植物的竞争能力[11,23]. 如Peltzer通过野外实验发现,克隆整合并没有显著提高Populustremuloides入侵本地草原植物群落时的竞争力[23]. Wang等也发现在温室控制实验条件下,克隆整合也没有提高喜旱莲子草入侵草本植物高羊茅(Festucaarundinacea)种群的竞争力[11]. 相反,王宁则发现匍匐茎连接显著提高了喜旱莲子草入侵北美车前种群的竞争能力[15]. 而在本实验中,克隆整合同样显著增强了喜旱莲子草入侵黄花水龙群落时的竞争能力(匍匐茎连接时的RNE显著大于切断时的值). 对于以上种种差异,原因可能为:一是植物的竞争能力常常因竞争对象、环境条件和时间迁移而发生动态变化[15];二是因为入侵植物的受体分株(或子株)对竞争植被的表现还取决于与其相连的供体分株(或母株)所处的环境条件. 如在近期的研究中,You等发现当喜旱莲子草基株处于营养贫瘠的生境时,克隆整合并不会提升其端株入侵本地植被时的竞争力;而当其基株生长在营养丰富的斑块时,增强了喜旱莲子草端株的竞争能力[16]. 本实验中,所用的基质为泥沙混合物和缓释肥,营养较为丰富. 因此,处于营养丰富条件的基株能通过克隆整合作用给予受竞争胁迫的端株更多的资源支持,进而提升其竞争能力,有利于其入侵.
然而,匍匐茎连接处理并没有影响竞争植被黄花水龙种群的生长,即克隆整合虽然显著提升了喜旱莲子草的竞争能力,但不影响黄花水龙种群的表现. 分析其原因,可能是由于实验初期喜旱莲子草与黄花水龙处于不平衡的竞争关系,喜旱莲子草的生物量的变化不足以影响黄花水龙种群的生长[11,16]. 这一点从喜旱莲子草在竞争存在时的生长参数有较大幅度的下降可以看出. 因此,尽管克隆整合显著促进了喜旱莲子草在黄花水龙种群中的生长、克隆繁殖,增加了其竞争力,但这些积极效应在本实验有限的时间周期中(8周)并不能影响黄花水龙种群的适合度.
克隆整合会显著影响克隆植物在异质性环境中的生物量分配,这一点在以往众多研究中得到证实[5,10-11,15,24]. 本实验中,在无竞争时,克隆整合有降低喜旱莲子草根冠比的趋势. 相反,在与黄花水龙竞争时,克隆整合则显著提高了其根冠比. 这一结果产生的原因可能是:1)无竞争时,克隆整合将更多的生物量投资到冠层,有利于喜旱莲子草占据空白生境,表明克隆整合对喜旱莲子草新生分株的生长以及占据空白生境极为重要,进而有利于提高其在自然生境中的入侵[11,15];2)有竞争时,切断处理分株的表现类似非克隆植物,将更多的生物量投入到地上部分以获取更多因竞争植被覆盖所缺失的光照和空间资源(趋贫性);连接处理的分株则体现了克隆植物劳动分工现象(环境诱导所致),即其地下资源相对于竞争而造成地上光照资源的缺乏来说更为丰富,因而分株将更多的生物量投入到地下部分以获取更丰富的地下土壤养分等资源并与相连的分株共享,从而提升植株的营养获取效率,进而提高整个克隆分株系的表现[11,16,25].
综上所述,在有竞争的环境中,喜旱莲子草的克隆整合对其光合性能和生长都有利,并且能大大提高其对本地植物的竞争能力,从而证明克隆整合在喜旱莲子草的入侵过程中发挥着重要作用. 此外,克隆整合不仅有利于竞争环境中喜旱莲子草的生长和竞争,而且显著增加了其匍匐茎长和分株数(克隆繁殖),而克隆繁殖的增强能显著增加其无性繁殖体数量,这些繁殖又可由动物和人类的介导,从而由成功入侵的种群中扩散至其他区域,进一步增加其入侵范围[20]. 总体来说,本研究表明外来植物的克隆属性(如克隆整合)在其入侵过程中扮演着重要的角色[7]. 因此,在根除或者控制喜旱莲子草的过程中,需重视其克隆整合特征而有针对性地制定防控措施.
[1] Wan Fanghao, Guo Jianying, Zhang Feng eds. Research on biological invasions in China. Beijing: Science Press, 2009. [万方浩, 郭建英, 张峰. 中国生物入侵研究. 北京: 科学出版社, 2009.]
[2] Powell KI, Chase JM, Knight TM. Invasive plants have scale-dependent effects on diversity by altering species-area relationships.Science, 2013, 339: 316-318.
[3] Villamagna AM, Murphy BR. Ecological and socio-economic impacts of invasive water hyacinth (Eichhorniacrassipes): A review.FreshwaterBiology, 2010, 55: 282-298.
[4] Schooler SS.Alternantheraphiloxeroides(Martius) Grisebach. In: Francis RA ed. A handbook of global freshwater invasive species. London and New York: Earthscan, 2012: 25-35.
[5] You WH, Yu D, Liu CHetal. Clonal integration facilitates invasiveness of the alien aquatic plantMyriophyllumaquaticumL. under heterogeneous water availability.Hydrobiologia, 2013, 718: 27-39.
[6] Dong Ming, Yu Feihai, An Shuqingetal. Ecological consequences of plant clonality.JournalofPlantEcology, 2007, 31(4): 549-551. [董鸣, 于飞海, 安树青等. 植物克隆性的生态学意义. 植物生态学报, 2007, 31(4): 549-551.]
[7] Song YB, Yu FH, Keser LHetal. United we stand, divided we fall: a meta-analysis of experiments on clonal integration and its relationship to invasiveness.Oecologia, 2013, 171: 317-327.
[8] Hutchings MJ, Wijesinghe DK. Patchy habitats, division of labour and growth dividends in clonal plants.TrendsinEcology&Evolution, 1997, 12: 390-394.
[9] Alpert P, Holzapfel C, Slonimski C. Differences in performance between genotypes ofFragariachiloensiswith different degrees of resource sharing.JournalofEcology, 2003, 91: 27-35.
[10] Roiloa SR, Retuerto R. Small-scale heterogeneity in soil quality influences photosynthetic efficiency and habitat selection in a clonal plant.AnnalsofBotany, 2006, 98: 1043-1052.
[11] Wang N, Yu FH, Li PXetal. Clonal integration affects growth, photosynthetic efficiency and biomass allocation, but not the competitive ability, of the alien invasiveAlternantheraphiloxeroidesunder severe stress.AnnalsofBotany, 2008, 101: 671-678.
[12] Moola FM, Vasseur L. The importance of clonal growth to the recovery ofGaultheriaprocumbensL. (Ericaceae) after forest disturbance.PlantEcology, 2009, 201: 319-337.
[13] Yu F, Wang N, Alpert Petal. Physiological integration in an introduced, invasive plant increases its spread into experimental communities and modifies their structure.AmericanJournalofBotany, 2009, 96: 1983-1989.
[14] Liu Jian, Li Junmin, Yu Huaetal. The relationship between functional traits and invasiveness of alien plants.BiodiversityScience, 2010, 18(6): 569-576. [刘建, 李钧敏, 余华等. 植物功能性状与外来植物入侵. 生物多样性, 2010, 18(6): 569-576.]
[15] Wang Ning. Clonal integration enhance the competitiveness of the invasive plantAlternantheraphiloxeroidestoPlantagovirginicaL..EcologyandEnvironmentalSciences, 2010, 19(10): 2302-2306. [王宁. 克隆整合提高了入侵植物空心莲子草对北美车前的竞争力. 生态环境学报, 2010, 19(10): 2302-2306.]
[16] You WH, Fan SF, Yu Detal. An invasive clonal plant benefits from clonal integration more than a co-occurring native plant in nutrient-patchy and competitive environments.PLoSOne, 2014, 9 (5): e97246.
[17] Ma Ruiyan, Wang Ren. Invasive mechanism and biological control of alligator weed,Alternantheraphiloxeroides.ChinJApplEnvironBiol, 2005, 11(2): 246-250. [马瑞燕, 王韧. 喜旱莲子草在中国的入侵机理及其生物防治. 应用与环境生物学报, 2005, 11(2): 246-250.]
[18] Pan Xiaoyun, Ge Yupeng, Zhang Wenjuetal. InvasiveAlternantherephiloxeroides: Biology, ecology and management.ActaPhytotaxonomicaSinica, 2007, 45(6): 884-900. [潘晓云, 耿宇鹏, 张文驹等. 入侵植物喜旱莲子草——生物学, 生态学及管理. 植物分类学报, 2007, 45(6): 884-900.]
[19] You WH, Yu D, Xie Detal. The invasive plantAlternantheraphiloxeroidesbenefits from clonal integration in response to defoliation.Flora, 2014, 209: 666-673.
[20] You WH, Han CM, Fang LXetal. Propagule pressure, habitat conditions and clonal integration influence the establishment and growth of an invasive clonal plant,Alternantheraphiloxeroides.FrontiersinPlantScience, 2016, 7: 568.
[21] You WH, Han CM, Liu CHetal. Effects of clonal integration on the invasive clonal plantAlternantheraphiloxeroidesunder heterogeneous and homogeneous water availability.ScientificReports, 2016, 6: 29767.
[22] Schreiber U, Bilger W, Hormann Hetal. Chlorophyll fluorescence as a diagnostic tool: basics and some aspects of practical relevance. In: Raghavendra AS ed. Photosynthesis: a comprehensive treatise. Cambridge: Cambridge University Press, 1998: 320-336.
[23] Peltzer DA. Does clonal integration improve competitive ability? A test using aspen (Populustremuloides[Salicaceae]) invasion into Prairie.AmericanJournalofBotany, 2002, 89: 494-499.
[24] Qian Yongqiang. Physiological integration and its regulation between inter-ramet of buffalograss under heterogeneous water stress [Dissertation]. Beijing: Chinese Academy of Forestry, 2009. [钱永强. 异质性水分胁迫下野牛草克隆分株间生理整合及其调控机理[学位论文]. 北京: 中国林业科学研究院, 2009.]
[25] Lü Xiaoqian, Zhang Yinlong. Effects of clonal integration and simulated sediment burial on the growth and antioxidant system of the introduced aquatic plantMyriophyllumaquaticum.JLakeSci, 2016, 28(3): 616-623. DOI: 10.18307/2016.0318. [吕晓倩, 张银龙. 克隆整合和模拟沉积物淹埋对外来水生植物粉绿狐尾藻(Myriophyllumaquaticum)生长和抗氧化系统的影响. 湖泊科学, 2016, 28(3): 616-623.]
Clonal integration facilitating for the invasion ofAlternantheraphiloxeroides
FANG Longxiang1, LÜ Xiaoqian2, XI Daoguo1, YOU Wenhua1**, Yi Chengwu1& DU Daolin1
(1:CollegeoftheEnvironmentandSafetyEngineering,JiangsuUniversity,Zhenjiang212013,P.R.China)(2:CollegeofArchitecture&Art,HeFeiUniversityofTechnology,Hefei230009,P.R.China)
Many notorious alien invader are clonal plants, thus it is of great significance to understand the effect of clonal integration on their potential invasiveness. This study investigated clonal integration (stolon connection) on the photosynthetic performance, growth, competitive ability and biomass allocation ofAlternantheraphiloxeroideswhen it invaded the nativeLudwigiapeploidesplant population, to explore the contribution of clonal integration on the invasion of this plant. We found that plant competition significantly reduced the maximum quantum yield of photosystem II, final biomass, ramet number and total stolon length ofA.philoxeroides, whereas clonal integration greatly improved the photosynthetic performance, growth measures, significantly increased the competitive ability and root/shoot ratios ofA.philoxeroidesand facilitated its invasiveness.
Invasive plant; physiological integration; competitive ability; root/shoot ratio; photosynthetic performance;Alternantheraphiloxeroides
国家自然科学基金项目(31400312,31570414)资助. 2016-11-07收稿; 2017-01-03收修改稿. 方龙香(1992~),女,硕士研究生; E-mail: 987728375@qq.com.
; E-mail: wenhuayou186@gmail.com.
DOI 10.18307/2017.0518
