水分梯度下黑河流域荒漠植物群落多样性特征
2017-08-28张晓龙周继华蔡文涛管天玉高楠楠姜联合来利明杨大文丛振涛郑元润
张晓龙,周继华 ,蔡文涛,管天玉 ,高楠楠,杜 会,姜联合,来利明,杨大文,丛振涛,郑元润,*
1 中国科学院植物研究所, 北京 1000932 中国科学院大学, 北京 1000493 清华大学水利水电工程系, 北京 100084
水分梯度下黑河流域荒漠植物群落多样性特征
张晓龙1,2,周继华1,2,蔡文涛1,2,管天玉1,2,高楠楠1,2,杜 会1,姜联合1,来利明1,杨大文3,丛振涛3,郑元润1,*
1 中国科学院植物研究所, 北京 1000932 中国科学院大学, 北京 1000493 清华大学水利水电工程系, 北京 100084
物种多样性对大尺度环境条件的响应是植物生态学的热点问题,干旱区内陆河流域的特殊环境条件如何影响物种多样性仍存在较大的不确定性。以黑河流域荒漠植物群落为对象,探讨了荒漠植物群落α多样性和β多样性对不同水分梯度的响应。结果表明:(1)荒漠植物群落随水分条件不同而变化,在区域降水梯度上,依次分布有梭梭[Haloxylonammodendron(C. A. Mey.) Bunge]、西伯利亚白刺(NitrariasibiricaPall.)、毛瓣白刺(NitrariapraevisaBobr.)、沙蒿(ArtemisiadesertorumSpreng.)、细枝盐爪爪(KalidiumgracileFenzl)、珍珠猪毛菜(SalsolapasserinaBunge)和尖叶盐爪爪[Kalidiumcuspidatum(Ung.Sternb.) Grub.]群落;随距黑河河道距离增加,主要分布有胡杨(PopuluseuphraticaOliv.)、多枝柽柳(TamarixramosissimaLebed.)、花花柴[Kareliniacaspia(Pall.)Less.]和红砂[Reaumuriasongarica(Pall.) Maxim.]群落。(2)荒漠植物群落物种多样性指数偏低,Margalef指数、Shannon-Weiner指数和Simpson指数最大值仅为0.985、1.641、0.596,群落结构简单,物种组成单一,单寡种优势群落现象极为明显,群落相对稳定。(3)在区域降水梯度上,α多样性指数大致呈先增加后降低的变化趋势,在珍珠猪毛菜群落(190 mm)达到峰值后迅速下降,Sørensen相似指数在细枝盐爪爪群落(166 mm)与珍珠猪毛菜群落(190 mm)之间达到最大值,Cody指数相对较小,表明珍珠猪毛菜群落(190 mm)物种丰富度最高,细枝盐爪爪群落(166 mm)与珍珠猪毛菜群落(190 mm)之间β多样性最小,物种更替速率最慢,群落处于较稳定阶段。(4) 随距黑河河道距离增加,α多样性指数大体呈“M”型变化,在多枝柽柳群落(1 km)和多枝柽柳群落(4.1 km)存在两个峰值,此处物种多样性相对较高,Sørensen指数在1—1.5 km和4.1—4.8 km之间出现两个较小值,Cody指数出现两个峰值,两者之间β多样性最大,为物种更替速率相对较快的过渡地带。对荒漠植被,以重要值为指标计测多样性指数较为可行,荒漠植物群落的物种多样性并不是随水分的增加而直线增加,这对维持和管理荒漠植物群落的多样性具有重要意义。
黑河流域;荒漠植物群落;α多样性;β多样性;水分梯度
植物群落是植物在长期环境变化中自我调节和适应而形成的集合体,作为生态系统物质和能量的主要提供者,是维持生态系统稳定和持续生产的物质基础[1]。其自身发展总是趋于最大限度地利用当地环境资源,构成多样的群落结构特征,以维持群落结构和功能的稳定[2- 4]。在植物群落多样性研究中,群落α多样性是反映群落内物种组成状况的一个重要指标,即物种丰富度和均匀度,β多样性表示物种沿环境梯度发生替代的程度或物种变化的速率,α、β多样性构成总体多样性[5- 7]。植物群落α、β多样性是当前群落生态学乃至整个生态学研究中十分重要的内容[8- 11],研究植物群落多样性特征对揭示群落、生态系统结构和功能演变具有极为重要的意义[12]。
干旱区内陆河流域是具有独特生态-水文过程的生态系统,各要素之间以水为纽带进行着频繁的物质和能量交换,彼此强烈影响[13-14]。黑河流域作为我国西北干旱区第二大内陆河流域,干流从祁连山发源地到尾闾居延海,横跨山地、绿洲和荒漠3种不同的地理单元,形成独特的以水为纽带的“冰川/冻土—高山植被—山前绿洲—尾闾荒漠”多源景观以及相伴而生的复合生态系统[15- 17],特殊的自然环境使其成为研究不同水分梯度下植物群落结构、功能和多样性特征的理想场所。在黑河流域,水分梯度变化主要表现在3个方面,一是上游山区随海拔变化引起的水分梯度差异,二是山下纬度地带上随区域降水变化引起的水分梯度差异,三是沿河道远近随地下水位变化引起的水分梯度差异。这3种水分梯度变化均会导致植物群落结构和功能的改变,也会导致植物群落多样性的变化。黑河中下游地区荒漠分布广泛,荒漠植物群落是植被的重要组成部分[18- 21],荒漠植物分布和演替受水分条件限制并表现出一定的响应规律[22- 25],水分成为维系荒漠地区植物群落生存和发展的最重要因子。目前,有关荒漠植物群落特征[9-10,26-27]、物种多样性及其对单一因素如土壤水、地下水等水分变化的响应研究较多[22,28- 30],对于不同类型水分梯度变化对黑河流域荒漠植物群落多样性的影响则关注较少。在黑河流域,沿自然降水梯度荒漠植物群落α、β多样性如何变化?垂直河岸带不同距离区域荒漠植物群落的α、β多样性又如何变化?这些问题还未得到很好阐释。
本文以黑河流域荒漠植物群落为研究对象,研究植物群落α、β多样性沿区域降水和黑河垂直距离远近的变化规律,探讨群落多样性与水分之间的关系,为更好理解黑河流域生态-水文关系,维持黑河流域荒漠生态系统的稳定性提供科学依据。
1 研究区概况
黑河流域是我国西北典型的内陆河流域,干流发源于祁连山北麓,途径青海、甘肃和内蒙古三省(区),全长821 km,海拔高程为900—5500 m,流域总面积约14.29x104km2[17,31]。流域地处内陆,气候恶劣,地形复杂,地势南高北低,呈连续下降趋势,地貌景观大体可分为:冰雪冻土带、高山植被带、山前绿洲带和荒漠带[16- 17,32]。上游植被茂密、冰川发育,年均降水量200—800 mm,为黑河流域径流主要形成区,中下游农田荒漠广布,降水稀少,年均降水量28—200 mm,最小年仅17 mm,为黑河流域径流的主要消耗区[31]。其中1961年和1992年尾闾西居延海和东居延海相继干涸,部分河段发生断流现象,植被生长受干旱胁迫的影响程度较重,生态环境十分脆弱,由于黑河流域生态应急输水工程的实施,中下游荒漠地区地下水每年都有固定地表河水的补给,使得黑河中下游荒漠地区地下水位不断升高。有研究表明,2000年以来,下游额济纳绿洲的地下水受到地表河水的补给,地下水埋深基本维持在3 m左右,且年际波动较小[33],基于乌兰图格嘎查垂直于河道的7口生态监测井数据,黑河下游(乌兰图格)年内最大地下水埋深为3.96 m,最小地下水埋深为1.05 m,地下水埋深与距河道远近呈正相关关系[34]。
黑河流域植被以荒漠为主,约占流域总面积的76.94%[35],荒漠植物群落分布广泛,由于受到水分条件的限制,植被分布格局明显呈斑块化分布,植被生产力低下,群落结构简单,种类组成单一。植被类型主要以荒漠戈壁成分为主,乔木有胡杨(PopuluseuphraticaOliv.)、梭梭[Haloxylonammodendron(C. A. Mey.) Bunge];灌木、半灌木有柽柳(Tamarixspp.)、西伯利亚白刺(NitrariasibiricaPall.)、沙蒿(ArtemisiadesertorumSpreng.)和红砂[Reaumuriasongarica(Pall.) Maxim.]等;草本有苦豆子(SophoraalopecuroidesLinn.)、花花柴[Kareliniacaspia(Pall.) Less.]和骆驼蓬(PeganumharmalaL.)等。
2 研究方法
2.1 野外调查
依据流域区域自然降水及距黑河远近的水分梯度布设野外观察样地,共设置样地17个,在区域降水梯度上,在流域荒漠区按自然降水梯度随机设置7个调查样地,样地选择在远离河流和公路的荒漠地区,自然降水为唯一水分来源,降水数据来自1964—2014年日降水均值[31],由小到大依次记为P1(29 mm)、P2(30 mm)、P3(65 mm)、P4(77 mm)、P5(166 mm)、P6(190 mm)、P7(291 mm)。在沿河梯度上,参照2010年在黑河下游额济纳旗乌兰图格嘎查垂直于河道布设的7 口水位监测井,在大体垂直于主河道5.5km范围内设置10个典型样地,记为T0.2、T0.5、T1、T1.5、T3.5、T3.7、T3.9、T4.1、T4.8、T5.5,其中,字母T后面的数字代表距离黑河的垂直距离(km),植被类型依据样地调查资料计算物种重要值确定。
采用随机取样法进行样方调查,每个植被调查样地设置3个重复样方进行测定。样方大小分别为:乔木20 m×20 m、灌木5 m×5 m、草本植物1 m×1 m。记录样地基本状况和群落特征,包括样地经纬度、海拔、生境、地貌和土壤等属性,以及植物种类、株数、高度、冠幅、胸(地)径、密度、盖度、频度和优势种等群落特征。野外调查时间为2015年7—8月。
2.2 数据统计分析
在统计各个样方每种植物多度、密度、盖度和频度的基础上,分别按乔木、灌木和草本植物计算样方内各植物物种的重要值,公式为:重要值IV=(相对密度+相对盖度+相对频度) /3[36]。
物种多样性指数较多,在物种重要值基础上,依据物种多样性指数应用的广泛程度,本文选取6种多样性指数,公式如下:
α多样性[37-38]
Margalef 丰富度指数
Rm=(S-1) / lnN
Simpson指数(优势度指数)
Shannon-Wiener多样性指数
Pielou 指数(均匀度指数)
E=H′/lnS
式中,Pi为种i的相对重要值,N为样方中物种个体总数,S为样方内物种数,多样性指数以重要值计算。
β多样性[37,39]:
Sørensen指数:
Cody指数:
式中,a和b分别为两样方的物种数,c为两样方的共有物种数;Sørensen指数反映群落或样方间物种组成的相似性; Cody指数反映物种沿梯度的替代速率。
3 结果
3.1 植物群落类型与分布
在自然降水梯度上,生境以戈壁平原风蚀残貌为主,群落结构简单,物种组成1—7种,植被以半灌木、灌木为主,属于典型戈壁荒漠植被类型。梭梭群落(P1)和西伯利亚白刺群落(P2)降水量少于50 mm,群落盖度低,植被稀疏,以单优势种为主,建群种重要值均为1;毛瓣白刺(NitrariapraevisaBobr.)群落(P3)和沙蒿群落(P4)降水量为50—100 mm,群落结构简单,群落高度、盖度较低,建群种重要值为0.76、0.72;细枝盐爪爪(KalidiumgracileFenzl)群落(P5)和珍珠猪毛菜(SalsolapasserinaBunge)群落(P6)降水量为100—200 mm,群落盖度较大,物种组成最多,优势物种明显,建群种重要值为0.51、0.61;尖叶盐爪爪[Kalidiumcuspidatum(Ung.Sternb.) Grub.]群落(P7)降水量大于200 mm,群落盖度最大,建群种重要值为0.69(表1)。
在沿河道距离梯度上,植物群落生境由河岸沙地向荒漠戈壁地貌过渡,植被以乔木、灌木为主,多年生草本也占很重要的地位,群落结构简单,物种组成1—6种,单优群落较多。有分布于河岸沙地上的胡杨、多枝柽柳、苦豆子和骆驼蓬等,分布于荒漠戈壁上的花花柴和红砂等。胡杨群落(T0.2)、花花柴群落(T4.8)和红砂群落(T5.5)以单优种存在,建群种重要值均为1;多枝柽柳群落(T0.5—T4.1),群落盖度较高,物种组成丰富,建群种重要值为0.46—1;多枝柽柳群落(T1)盖度最高,物种组成最为丰富,建群种重要值为0.7(表1)。
3.2 区域降水梯度上的植物群落多样性特征
在区域降水梯度上,植物群落多样性水平不高,物种组成1—7种,指数值均小于1.7,物种α多样性指数随降水增加(P1—P7)大致呈现先增加后减少的变化趋势,α多样性指数最大值出现在珍珠猪毛菜群落(P6)样地(图1)。Margalef丰富度指数(Rm)在0—0.985间变化,各群落大小依次为珍珠猪毛菜群落(P6)>沙蒿群落(P4)>细枝盐爪爪群落(P5)>毛瓣白刺群落(P3)>尖叶盐爪爪群落(P7)>梭梭群落(P2)和西伯利亚白刺群落(P1)。Shannon-Weiner指数在0—1.641之间变化,以珍珠猪毛菜群落(P6)最高(1.641),沙蒿群落(P4)、细枝盐爪爪群落(P5)、毛瓣白刺群落(P3)和尖叶盐爪爪群落(P7)等较高(0.598—1.174),Simpson优势度指数和Pielou均匀度指数分别在0—0.597、0—0.753之间变化,大致与多样性指数变化趋势一致。单优势种梭梭群落(P2)和西伯利亚白刺群落(P1),群落结构最为简单,多样性指数、均匀度指数均为0。
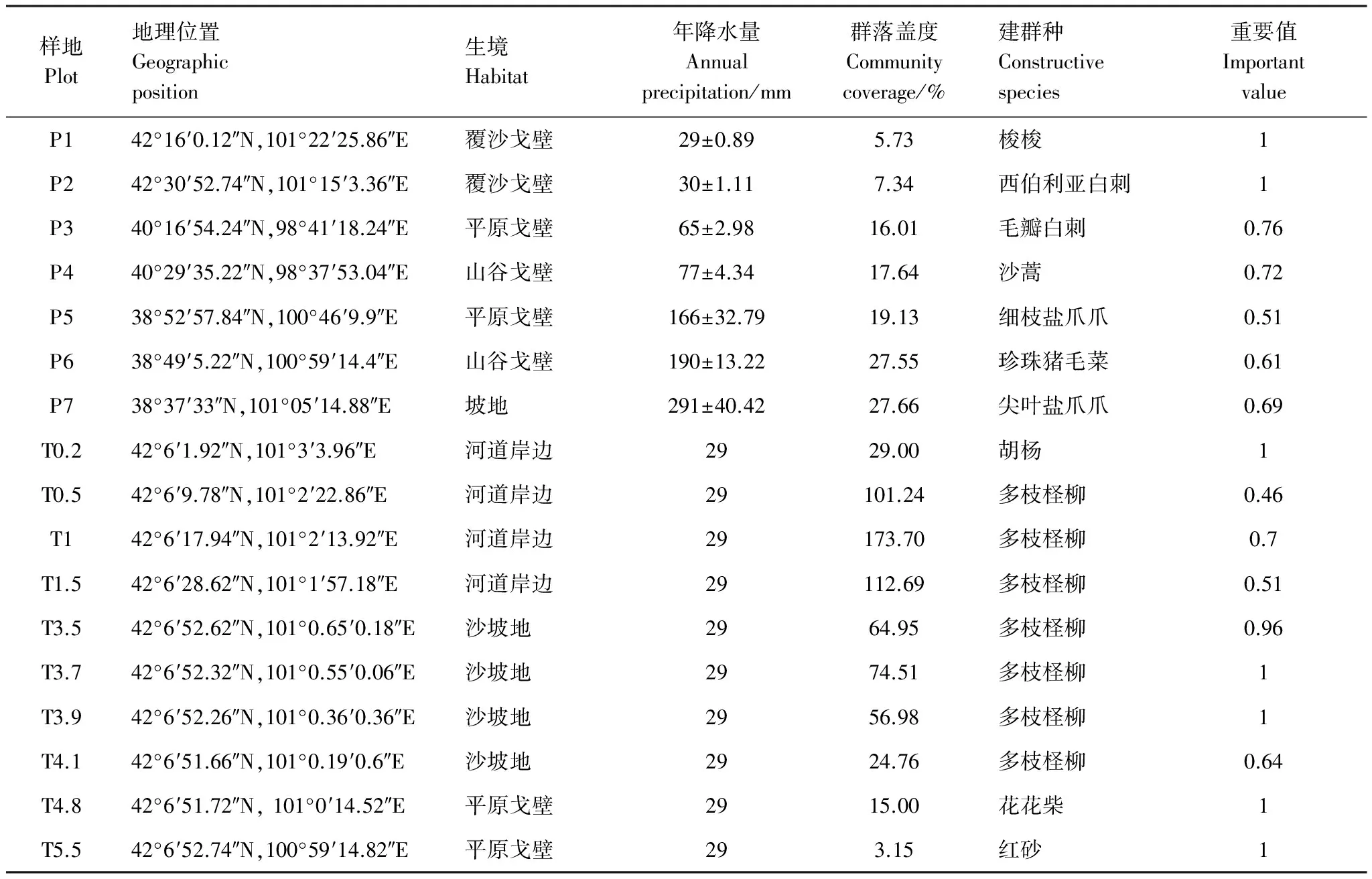
表1 样地基本信息
P1: 梭梭群落Haloxylonammodendroncommunity; P2: 西伯利亚白刺群落Nitrariasibiricacommunity; P3: 毛瓣白刺群落Nitrariapraevisacommunity; P4: 沙蒿群落Artemisiadesertorumcommunity; P5: 细枝盐爪爪群落Kalidiumgracilecommunity; P6: 珍珠猪毛菜群落Salsolapasserinacommunity; P7: 尖叶盐爪爪群落Kalidiumcuspidatumcommunity; T0.2: 胡杨群落Populuseuphraticacommunity;T0.5—T4.1, 多枝柽柳群落Tamarixramosissimacommunity;T4.8, 花花柴群落Kareliniacaspiacommunity; T5.5, 红砂群落Reaumuriasongaricacommunity
区域降水梯度之间,植物群落Sørensen指数随降水增加总体上呈先上升后下降的变化趋势(图2),细枝盐爪爪群落(P5)和珍珠猪毛菜群落(P6)之间的Sørensen相似系数最大,为0.66,而梭梭群落(P1)、西伯利亚白刺群落(P2)、毛瓣白刺群落(P3)和沙蒿群落(P4)两者之间Sørensen相似系数为0。Cody指数与Sørensen指数随降水的变化趋势大致相反,细枝盐爪爪群落(P5)和珍珠猪毛菜群落(P6)之间的Cody指数接近于最小值1。珍珠猪毛菜群落(P6)和尖叶盐爪爪群落(P7)之间的Cody指数明显高于其他群落之间的Cody指数,可见,随着降水增加,植物群落间的物种更替速率先加快、再降低,最终达到最高点。
3.2 沿河梯度下的植物群落多样性特征
在沿河梯度上,植物群落Shannon-Weiner指数呈“M”型变化趋势,在距河道近处多枝柽柳群落(T1)呈现出一个最大峰值,迅速下降后,指数值呈现出较大程度的上升趋势,多枝柽柳群落(T4.1)到达一个较大的峰值,其后随着离河道距离越来越远,指数值到达最小,接近于0(图3)。Simpson多样性指数和Pielou均匀度指数分别在0—0.564、0—0.726之间变化,大体趋势与Shannon-Weiner多样性指数相同。胡杨群落(T0.2)、多枝柽柳群落(T3.5—T3.9)、花花柴群落(T4.8)和红砂群落(T5.5)几乎呈单优势群落,群落结构最为简单,群落多样性、优势度和均匀度水平最低,接近于0。
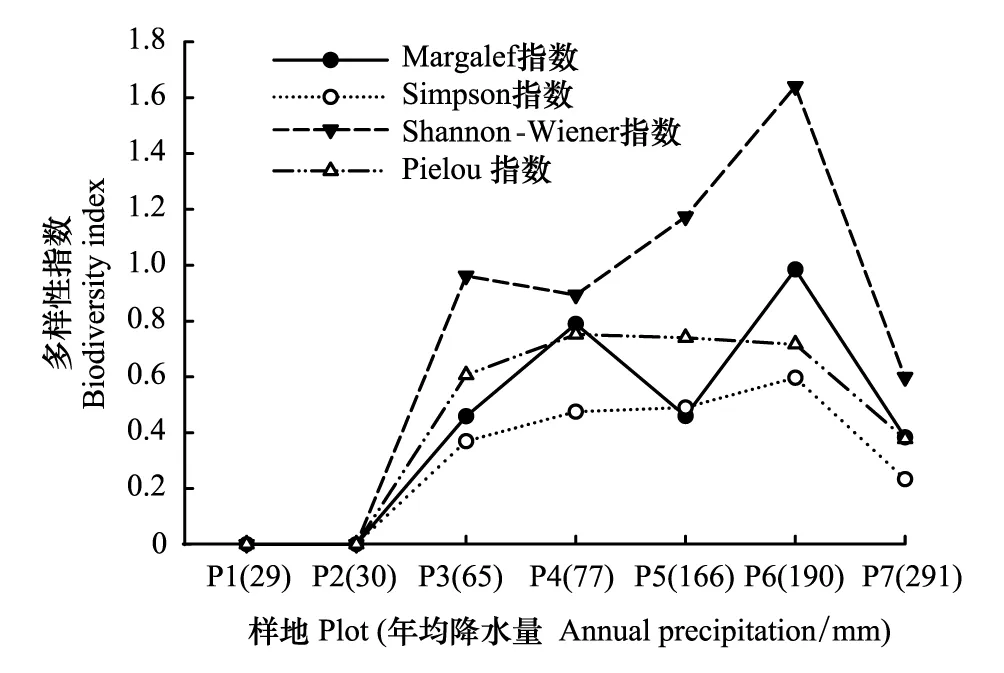
图1 α多样性指数随降水的变化Fig.1 α diversity indices along precipitation gradients P1—P7, 自然降水量呈由小到大趋势;样地代码同表1
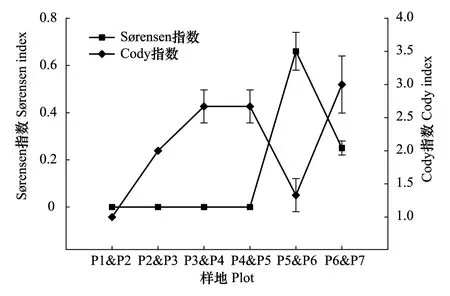
图2 不同降水梯度植物群落间Sørensen指数和Cody指数Fig.2 Sørensen and Cody indices of plant communities along precipitation gradients样地代码同表1
随距河道距离增加,不同植物群落之间的Sørensen指数总体上呈现先下降再上升后迅速下降的趋势(图4)。多枝柽柳群落(T3.7)和多枝柽柳群落(T3.9)为单优群落,群落组分相同,两者Sørensen相似指数达到最大值,多枝柽柳群落(T1)和多枝柽柳群落(T1.5)之间与多枝柽柳群落(T4.1)和花花柴群落(T4.8)之间Sørensen指数相对较低。植物群落间Cody指数在沿河梯度上的变化趋势明显与Sørensen指数相反,多枝柽柳群落(T3.7)和多枝柽柳群落(T3.9)之间Cody指数达到最小值0,多枝柽柳群落(T1)和多枝柽柳群落(T1.5)之间与多枝柽柳群落(T4.1)和花花柴群落(T4.8)之间Cody指数值明显高于其它群落。
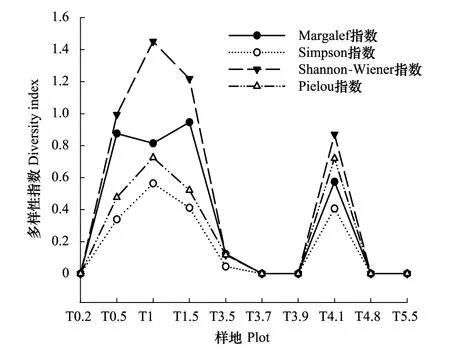
图3 α多样性指数随沿河距离的变化Fig.3 α diversity indices along vertical distance to river

图4 沿河梯度植物群落间Sørensen指数和Cody指数Fig.4 Sørensen and Cody indices of plant communities along vertical distance to river (样地代码同表1)
4 讨论
本研究聚焦于黑河流域在区域降水和沿河道距离远近两个梯度上荒漠植物群落的多样性特征。总体上,荒漠植物群落结构简单,物种组成稀少,17个荒漠植物群落中只有5个群落物种组成大于3种,单寡种优势群落现象极为明显,梭梭群落、西伯利亚白刺群落、胡杨群落等均为单优势种群落,这与荒漠地区干旱恶劣的生境条件有关[40]。植物群落多样性指数可以客观反映群落内物种组成特征,在所有水分梯度上,植物群落α多样性指数偏低,Margalef物种丰富度指数、Shannon-Weiner多样性指数和Simpson优势度指数最大值仅为0.985、1.641、0.596,植物群落物种多样性与其它荒漠区植被物种多样性水平相当[9,22,41-42],说明荒漠植物群落物种多样性水平普遍较低,进一步反映了该地区植物群落结构简单、物种组成稀少且种类分布极不均匀的荒漠植物群落特点。荒漠植物群落盖度随降水量增加明显呈现上升趋势,植被盖度的大小与降水量呈正相关关系(表1),表明荒漠植物群落盖度变化对降水量的响应非常敏感。在沿河梯度上,有研究表明,距河道1.2 km处植被盖度最大[43],本研究发现植物群落盖度最大值出现在多枝柽柳群落(T1),而不是在离河道最近的胡杨群落(T0.2),表明距河道最近植物群落盖度并非最高。
在荒漠地区,以水分梯度为特征的物种多样性格局和分布构建了荒漠生态系统的结构和功能,水分是荒漠地区生态-水文过程的关键影响因子和主要驱动力,而物种多样性对水分条件变化的响应直接影响着荒漠生态系统的功能和稳定性[44]。有研究表明,结构简单、单寡种优势荒漠植物群落往往具有较高的稳定性,物种多样性低并不代表系统稳定性差[29,45],在不同水分梯度上,如苦豆子和多枝柽柳等群落物种组成较丰富,仅分布在水分条件较好的河岸周围,处于不稳定状态,近年来,沿河地带都有上中游固定地表河水的补给,植物群落处于较稳定状态,水分是影响其生存和发展的主要因素。红砂、梭梭和西伯利亚白刺等群落物种组成单一,地处极度干旱地区,形成稳定的群落结构,这可能是荒漠植物群落长期适应干旱风沙环境而形成的适应对策。
在区域降水梯度上,荒漠植物群落α多样性指数随降水量增加总体呈先增加后降低的变化趋势,说明水分条件是限制荒漠植物群落生存和发展的最主要因子。α多样性指数在珍珠猪毛菜群落(190 mm)达到最大值,在降水量持续增大(大于200 mm)的情况下,α多样性指数在尖叶盐爪爪群落(291 mm)迅速下降。植物群落的物种多样性并不是随降水量的增加而直线增加,出现上述情况可能与尖叶盐爪爪群落自身及所处生境有关,尖叶盐爪爪属于典型的泌盐植物[46],所处环境盐分多集中在地表,形成盐结皮,含盐量可达35%—37%,在盐生环境条件下其它植物生长受限,尖叶盐爪爪形成单寡种优群落,从而导致尖叶盐爪爪群落(P7)多样指数低于珍珠猪毛菜群落(P6)样地。
在沿河梯度上,α多样性指数随距河道距离增加大体呈“M”型变化趋势,在多枝柽柳群落(T1)和多枝柽柳群落(T4.1)存在两个峰值,距河较近的胡杨群落(T0.2)物种多样性并非最高。参照2010年在黑河下游额济纳旗乌兰图格嘎查垂直于河道布设的7 口水位监测井数据,随距河道距离增加地下水埋深逐渐加深,植物群落物种多样性并不是随着地下水变化而呈直线变化趋势。理论上在一定沿河地带范围内,离河较近地区,土壤水分条件最好,植物多样性应该越高[11],这与本文结果有所不同,这主要与高地下水位变化引起土壤性质变化有关。胡杨群落(T0.2)地下水埋深为1.05—2.6 m[34],所处生境土壤水分条件好,地下水位高,土壤蒸发旺盛导致近河岸带浅层土壤盐分集聚,使植物群落面临盐碱胁迫[47],这对深根系植物胡杨和柽柳的生长有利,对于浅根和不耐盐碱的植物不利[48],因此,河岸边形成以胡杨占绝对优势的胡杨群落,植物群落的物种多样性水平较低。随距河道距离增加,地下水位下降,苦豆子、花花柴和骆驼蓬等草本植物出现,使其物种丰富度达到最大,植物群落物种多样性最大值出现在沿河梯度1—1.5 km,在此区域内适宜的土壤水分及较低的土壤盐分维持了较高的物种多样性。随距河道距离的进一步增加,地下水位持续下降,土壤水分明显下降,不能维持较高的物种多样性,物种多样性指数迅速下降。在降水梯度上,随着降水量的增加,物种多样性随降水增加,先增加后降低,在沿河梯度上,随水分条件变化,总体上呈先增加后降低的趋势,其中在沿河梯度上多枝柽柳群落(T4.1)样地α多样性指数有所升高,可能与“柽柳-黑果枸杞-花花柴”灌木草本植物组合存在有一定关系。有研究表明,柽柳主要吸收200—300 cm的深层土壤水,多年生草本植物花花柴主要吸收50—100 cm的中层土壤水,柽柳与其它物种之间水分利用存在互助关系[49-50],在土壤含水量较低的情况下,群落中各物种可以长期共存,提高对水资源的利用率,增加群落的稳定性,从而保持T4.1处较高的物种丰富度。
β多样性用于反映环境梯度下植物群落间的差异性,当一个群落向另一个群落过渡时,环境梯度变化较大,不同群落或样方间共有种越少,β多样性越大[51]。在降水梯度上,梭梭群落(P1)和西伯利亚白刺群落(P2)、西伯利亚白刺群落(P2)和毛瓣白刺群落(P3)、毛瓣白刺群落(P3)和沙蒿群落(P4)之间 Sørensen相似系数为0,原因在于在降水量小于100mm的荒漠地区,群落间生境差异较大,植物群落多为单寡种优势群落,群落间物种相似性低。随着降水量的增加,群落间的Sørensen相似系数在细枝盐爪爪群落(P5)和珍珠猪毛菜群落(P6)之间达到最大值0.66,然而在珍珠猪毛菜群落(P6)和尖叶盐爪爪群落(P7)之间迅速降低到0.25,同时Cody指数从1.33增加到3,说明细枝盐爪爪群落(P5)和珍珠猪毛菜群落(P6)之间β多样性最小,即细枝盐爪爪群落(P5)和珍珠猪毛菜群落(P6)所处区域荒漠植物群落结构最为稳定,物种更替速率最小。在沿河梯度上,Sørensen相似系数大体上呈现先下降再上升后迅速下降的趋势,Cody指数变化趋势与之相反。Sørensen相似系数在多枝柽柳群落(T1)和多枝柽柳群落(T1.5)之间和多枝柽柳群落(T4.1)和花花柴群落(T4.8)之间出现两个较小值,Cody指数则在两者出现两个峰值,说明多枝柽柳群落(T1)和多枝柽柳群落(T1.5)之间和多枝柽柳群落(T4.1)和花花柴群落(T4.8)之间β多样性最大,物种更替速率相对较快,T1&T1.5之间和T4.1&T4.8之间过渡性质明显,有研究表明,距黑河河道1.2 km处为绿洲向荒漠过渡区过渡的地带,距河道4.3 km处为荒漠过渡区和荒漠区的过渡地带[43],这与本研究结果相符。由于荒漠生境恶劣,受调查季节和空间所限,各植物群落与环境因子之间定量关系的分析有待深入研究。
[1] Lieth H. Modeling the primary productivity of the world//Lieth H, Whittaker R, eds. Primary Productivity of the Biosphere. New York: Springer-Verlag, 1975: 237- 263.
[2] Levine J M. Complex interactions in a streamside plant community. Ecology, 2000, 81(12): 3431- 3444.
[3] Lomolino M V. Elevation gradients of species-density: historical and prospective views. Global Ecology and Biogeography, 2001, 10(1): 3- 13.
[4] Lortie C J, Brooker R W, Chole P, Kikvidze Z, Michalet R, Pugnaire F I, Callaway R M. Rethinking plant community theory. Oikos, 2004, 107(2): 433- 438.
[5] 马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268- 277.
[6] 马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱβ多样性的测度方法. 生物多样性, 1995, 3(1): 38- 43.
[7] 张金屯. 数量生态学. 北京: 科学出版社, 2011: 77- 98.
[8] Franklin J F. Preserving biodiversity: species in landscapes: response. Ecological Applications, 1994, 4(2): 208- 209.
[9] 马斌, 周志宇, 张莉丽, 高文星, 陈善科, 张宝林. 阿拉善左旗植物物种多样性空间分布特征. 生态学报, 2008, 28(12): 6099- 6106.
[10] 陈杰, 郭屹立, 卢训令, 丁圣彦, 苏思, 郭静静, 李乾玺. 伊洛河流域草本植物群落物种多样性. 生态学报, 2012, 32(10): 3021- 3030.
[11] 张雪妮, 吕光辉, 王庭权, 马玉, 阿布里孜·阿不都热合曼, 赵晓英, 郭振洁, 朱修逸. 荒漠区垂直河岸带植物多样性格局及其成因. 生态学报, 2015, 35(18): 5966- 5974.
[12] Pandey S K, Shukla R P. Plant diversity in managed sal (ShorearobustaGaertn.) forests of Gorakhpur, India: species composition, regeneration and conservation. Biodiversity and Conservation, 2003, 12(11): 2295- 2319.
[13] 赵文智, 程国栋. 干旱区生态水文过程研究若干问题评述. 科学通报, 2001, 46(22): 1851- 1857.
[14] Montaldo N, Rondena R, Albertson J D, Mancini M. Parsimonious modeling of vegetation dynamics for ecohydrologic studies of water-limited ecosystems. Water Resources Research, 2005, 41(10): W10416.
[15] 卢玲, 李新, 程国栋, 肖洪浪. 黑河流域景观结构分析. 生态学报, 2001, 21(8): 1217- 1224.
[16] 卢玲, 程国栋, 李新. 黑河流域中游地区景观变化研究. 应用生态学报, 2001, 12(1): 68- 74.
[17] 黑河计划数据管理中心. 黑河流域边界数据集. 兰州: 中科院寒区旱区环境与工程研究所, 2011.
[18] 斯琴巴特尔, 秀敏. 荒漠植物蒙古扁桃水分生理特征. 植物生态学报, 2007, 31(3): 484- 489.
[19] 赵良菊, 肖洪浪, 程国栋, 宋耀选, 赵亮, 李彩芝, 杨秋. 黑河下游河岸林植物水分来源初步研究. 地球学报, 2008, 29(6): 709- 718.
[20] Odum E P. The strategy of ecosystem development. Science, 1969, 164(3877): 262- 270.
[21] Ricklefs R E. Community diversity: relative roles of local and regional processes. Science, 1987, 235(4785): 167- 171.
[22] 张佩, 袁国富, 庄伟, 薛沙沙. 黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应. 生态学报, 2011, 31(22): 6677- 6687.
[23] 刘加珍, 李卫红, 吴纯渊, 陈永金. 荒漠河岸生态系统退化与物种多样性恢复研究. 干旱区研究, 2009, 26(2): 212- 220.
[24] 张海娜, 苏培玺, 李善家, 周紫鹃, 解婷婷, 赵庆芳. 荒漠区植物光合器官解剖结构对水分利用效率的指示作用. 生态学报, 2013, 33(16): 4909- 4918.
[25] 周晓兵, 张元明, 王莎莎, 张丙昌. 模拟氮沉降和干旱对准噶尔盆地两种一年生荒漠植物生长和光合生理的影响. 植物生态学报, 2010, 34(12): 1394- 1403.
[26] 鱼腾飞, 冯起, 司建华, 席海洋. 黑河下游额济纳绿洲植物群落特征与物种多样性研究. 西北植物学报, 2011, 31(5): 1032- 1038.
[27] 王国宏, 任继周, 张自和. 河西山地绿洲荒漠植物群落种群多样性研究I生态地理及植物群落的基本特征. 草业学报, 2001, 10(1): 1- 12.
[28] 朱军涛, 于静洁, 王平, 王志勇. 额济纳荒漠绿洲植物群落的数量分类及其与地下水环境的关系分析. 植物生态学报, 2011, 35(5): 480- 489.
[29] 李新荣, 何明珠, 贾荣亮. 黑河中下游荒漠区植物多样性分布对土壤水分变化的响应. 地球科学进展, 2008, 23(7): 685- 691.
[30] 陈亚宁, 张宏锋, 李卫红, 陈亚鹏. 新疆塔里木河下游物种多样性变化与地下水位的关系. 地球科学进展, 2005, 20(2): 158- 165.
[31] Gao B, Qin Y, Wang Y H, Yang D W, Zheng Y R. Modeling ecohydrological processes and spatial patterns in the upper Heihe Basin in China. Forests, 2016, 7(1), doi: 10.3390/f7010010.
[32] 康尔泗, 程国栋, 蓝永超, 金会军. 西北干旱区内陆河流域出山径流变化趋势对气候变化响应模型. 中国科学(D辑), 1999, 29(S1): 47- 54.
[33] 蒋晓辉, 刘昌明. 黑河下游植被对调水的响应. 地理学报, 2009, 64(7): 791- 797.
[34] 周洪华, 李卫红, 木巴热克·阿尤普, 徐茜. 荒漠河岸林植物木质部导水与栓塞特征及其对干旱胁迫的响应. 植物生态学报, 2012, 36(1): 19- 29.
[35] 王建华, 赵军, 胡晓利, 盖春梅. 2012年新编黑河流域2000年土地利用/土地覆盖数据集. 黑河: 黑河计划数据管理中心, 2014.
[36] 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、 方法和技术规范. 生物多样性, 2009, 17(6): 533- 548
[37] Magurran A E. Ecological Diversity and Its Measurement. New Jersey: Princeton University Press, 1988.
[38] 钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994: 141- 165.
[39] Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213- 251.
[40] 李新荣, 张新时. 鄂尔多斯高原荒漠化草原与草原化荒漠灌木类群生物多样性的研究. 应用生态学报, 1999, 10(6): 665- 669.
[41] 张锦春, 王继和, 赵明, 刘虎俊, 廖空太, 徐先英. 库姆塔格沙漠南缘荒漠植物群落多样性分析. 植物生态学报, 2006, 30(3): 375- 382.
[42] 张林静, 岳明, 张远东, 顾峰雪, 潘晓玲, 赵桂仿. 新疆阜康绿洲荒漠过渡带植物群落物种多样性特征. 地理科学, 2003, 23(3): 329- 334.
[43] 路明, 刘加珍, 陈永金, 路亚坤. 黑河流域下游绿洲-荒漠梯度上的植被数量特征. 西北农业学报, 2013, 22(6): 176- 182.
[44] Whitford W G. Ecology of Desert Systems. New York: Academic Press, 2002.
[45] 李文华, 赵景柱. 生态学研究回顾与展望. 北京: 气象出版社, 2004: 625- 649.
[46] 章英才, 张晋宁. 尖叶盐爪爪叶的解剖结构与分析. 宁夏农学院学报, 2004, 25(2): 43- 45, 50- 50.
[47] 赵文智, 刘鹄. 荒漠区植被对地下水埋深响应研究进展. 生态学报, 2006, 26(8): 2702- 2708.
[48] 郑丹, 李卫红, 陈亚鹏, 刘加珍. 干旱区地下水与天然植被关系研究综述. 资源科学, 2005, 27(4): 160- 167.
[49] 陈小丽, 陈亚宁, 陈亚鹏. 黑河下游荒漠河岸林植物水分利用关系研究. 中国生态农业学报, 2014, 22(8): 972- 979.
[50] 付爱红, 陈亚宁, 李卫红. 中国黑河下游荒漠河岸林植物群落水分利用策略研究. 中国科学: 地球科学, 2014, 44(4): 693- 705.
[51] 王庆锁, 冯宗炜, 罗菊春. 河北北部、内蒙古东部森林-草原交错带生物多样性研究. 植物生态学报, 2000, 24(2): 141- 146.
Diversity characteristics of plant communities in the arid desert of the Heihe basin under different moisture gradients
ZHANG Xiaolong1,2, ZHOU Jihua1,2, CAI Wentao1,2, GUAN Tianyu1,2, GAO Nannan1,2, DU Hui1, JIANG Lianhe1, LAI Liming1, YANG Dawen3, CONG Zhentao3, ZHENG Yuanrun1,*
1InstituteofBotany,ChineseAcademyofSciences,Beijing100093,China2UniversityofChineseAcademyofSciences,Beijing100049,China3DepartmentofHydraulicEngineering,TsinghuaUniversity,Beijing100084,China
The responses of species diversity to the large-scale environment represent an important issue in plant ecology. In arid regions, the influence of the unique environmental conditions of inland river basins on species diversity remains uncertain. We examined the characteristics of plant community alpha and beta diversity under different moisture gradients in plant communities of the arid desert of the Heihe basin, Northwest China. The results showed that:(1) the type of desert plant community changed with moisture gradients. The communities which were most distinguished based on important value by increasing precipitation gradients were communities dominated byHaloxylonammodendron,Nitrariasibirica,Nitrariapraevisa,Artemisiadesertorum,Kalidiumgracile,Salsolapasserina, andKalidiumcuspidatum. The communities ofPopuluseuphratica,Tamarixramosissima,Kareliniacaspia, andReaumuriasongaricawere distributed along the moisture gradient, with species listed in order of increasing distance of their communities from rivers. (2) The species diversity indexes of desert plant communities were low; the maximum Margalef, Shannon-Wiener, and Simpson values were only 0.985, 1.641, and 0.596, respectively. The structure and composition of the studied communities were simple, and the communities were relatively stable. Moreover, monotypic (oligotypic) community types were prevalent. (3) Under the gradient of regional increasing precipitation,αdiversity showed a trend towards increasing and then decreasing; there was a high-value area around the community ofSalsolapasserina(190 mm), but then values decreased rapidly. The Sørensen index reached a maximum in the communities ofKalidiumgracile(166 mm) andSalsolapasserina(190 mm), while the Cody index remained relatively small. This suggests that the species richness was highest in theSalsolapasserinacommunity (190 mm). Theβdiversity reached minimum values in theKalidiumgracile(166 mm) andSalsolapasserina(190 mm) communities. The species replacement rate was also the lowest in these communities, indicating that these communities were relatively stable. (4) Theαdiversity showed a trend towards “M” type change with increasing distance from the river, and reached two peaks at around the community ofTamarixramosissima(1km) and the community ofTamarixramosissima(4.1 km). There were two smaller Sørensen index values approximately 1—1.5 km and 4.1—4.8 km, and the Cody index value showed a trend opposite to that of the Sørensen index. Furthermore, theβdiversity values were high, which suggests that these were transitional zones. We concluded that the diversity index based on important value was more applicable in arid desert areas and that the species diversity of the desert plant community showed no linear increase with increasing moisture. These results are important for maintaining and managing the diversity of desert plant communities.
Heihe basin; desert plant community;αdiversity;βdiversity; moisture gradients
国家自然科学基金项目(91225302)
2016- 04- 08; 网络出版日期:2017- 03- 02
10.5846/stxb201604080643
*通讯作者Corresponding author.E-mail: zhengyr@ibcas.ac.cn
张晓龙,周继华,蔡文涛,管天玉,高楠楠,杜会,姜联合,来利明,杨大文,丛振涛,郑元润.水分梯度下黑河流域荒漠植物群落多样性特征.生态学报,2017,37(14):4627- 4635.
Zhang X L, Zhou J H, Cai W T, Guan T Y, Gao N N, Du H, Jiang L H, Lai L M, Yang D W, Cong Z T, Zheng Y R.Diversity characteristics of plant communities in the arid desert of the Heihe basin under different moisture gradients.Acta Ecologica Sinica,2017,37(14):4627- 4635.
