生物化学教学中加强小分子非编码RNA介绍的必要性
2017-08-09乔梁刘彦礼
乔梁+刘彦礼
摘要:随着高通量测序技术的发展,越来越多的小分子非编码RNA(Small non-coding RNA,sncRNA)被人们发现,其介导的转录调控是生物体内一种新的调控模式。目前,microRNA(miRNA)、small interferi ng RNA(siRNA)和PIWl-interacti ng RNA(piRNA)是sncRNA中研究最为透彻的三个成员。笔者在简述三者的生物合成及作用机制的基础上,建议在《生物化学》的教学中增加sncRNA知识的介绍。
关键词:小分子非编码RNA;生物化学,教学
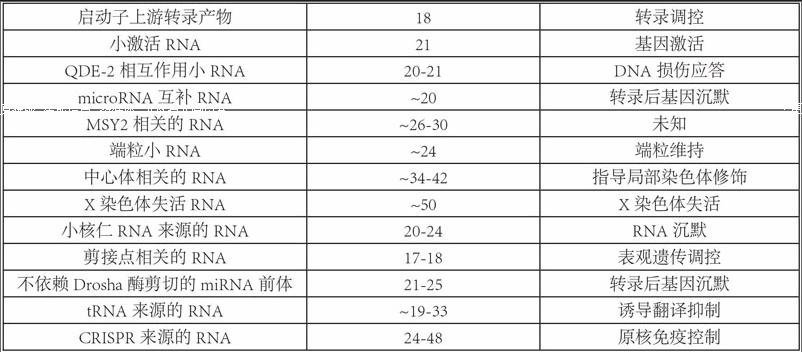
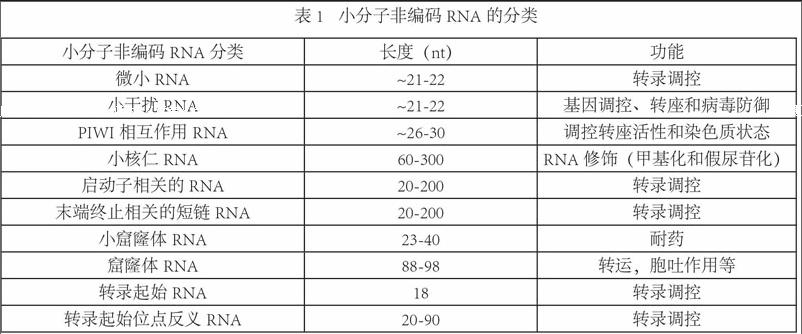
自从1993年在线虫中发现首个非编码RNA分子一lin-4并证明其调控功能后,开启了研究基因组内“暗物质”的新时代。小分子非编码RNA(sncRNA)是一類不编码蛋白质的小RNA分子,长度小于200个核苷酸(Nucleotide,nt),通常约20-30 nt。近年的研究发现,sncRNA具有重要的调控功能,参与了生物体的各种活动,如生长发育、器官形成、细胞增殖分化和凋亡,以及多种疾病的发生等。经过数年的探索,miRNA、siRNA和piRNA的功能和分子机制已被深入研究。除此之外,其他类型的sncRNA正逐渐为人所知(表1)。这些发现提示我们生物体内可能存在着一种全新的调控模式,将有助于我们深入了解生物体复杂的调控网络。
目前,sncRNA研究已成为生命科学和医学研究的热门领域,并获得了2006年的诺贝尔生理学或医学奖,但是在传统《生物化学》课程的教学中严重缺乏sncRNA知识的介绍。在此,笔者简要阐述sncRNA中研究深入的miRNA、siRNA和piRNA,从生物合成和作用机制入手,加强sncRNA知识的介绍,期望能够为《生物化学》课程的教学提供参考。
一、miRNA的生物合成和作用机制
miRNA是一类长约20-24 nt的内源性非编码单链小分子RNA,广泛存在于真核生物中,它可以通过mRNA剪切和抑制蛋白质翻译的方式负调控靶基因。目前,人类有接近1/3的基因受到miRNA的调控。进一步研究表明,同一个miRNA存在众多的靶基因,而同一个靶基因经常受到多个miRNA的共同调控,miRNA及其靶基因构建了生物体内全方位多层次的基因表达调控网络系统。
miRNA基因由RNA聚合酶Ⅱ转录,产生原始的miRNA转录物(pri-miRNA),pri-miRNA在细胞核内经RNA内切酶Ⅲ一Drosha加工,得到miRNA前体(pre-miRNA)。然后被Exportin-5蛋白转运到细胞质中,再经RNA内切酶Ⅲ一Dicer进一步加工得到约22个核苷酸的成熟双链RNA分子(miRNA:miRNA*),其中一条成熟链(miRNA)插入RNA诱导沉默复合体(RNA-induced silencingcomplex,RISC)中形成miRISC行使功能,而另外一条链(miRNA*)会被快速降解。
miRNA通过两种不同的机制下调靶基因的表达:mRNA剪切和翻译抑制。miRNA作用靶基因的3非翻译区(3UTR)内调控位点,以完全互补或不完全互补的形式结合,诱导靶基因mRNA降解或抑制蛋白翻译,导致相应蛋白质合成缺失或减少,从而在转录后水平调控基因表达,进而参与生物体内某些生理及病理过程。绝大多数的植物miRNA与其靶基因几乎完全互补,并且互补位点散布于整个mRNA而非局限于3UTR区,引发靶基因mRNA的降解。而绝大多数的动物miRNA与其靶基因互补程度较低,通过抑制翻译的方式调控靶基因表达。
二、siRNA的生物合成和作用机制
siRNA是一类长约21bP(Base pair,bP)的小分子非编码双链RNA,同样由Dicer切割完全互补的双链RNA复合物产生,广泛存在于真核生物中。早期(1999)发现外源导入细胞内的双链小RNA分子(dsRNA)形成siRNA,通过RNAi途径导致基因沉默,但是直到最近(2008)才在果蝇和哺乳动物中发现内源siRNA(Endogenous siRNA,endo-siRNA)。现今,由外源性合成的siRNA引发RNAi导致目的基因沉默,已成为基因功能研究不可或缺的方法,而且在疾病治疗中具有重要的意义。
外源性导入、病毒激活、转座子侵入及特异重复序列等因素均可诱导dsRNA产生,dsRNA转录后被Dicer剪切生成21-22 nt、3末端2 nt突出的RNA二聚体。在RISC形成过程中,siRNA二聚体会解聚形成两条单链siRNA,均可有效参与RISC的形成。同miRNA作用机制相同,siRNA可通过碱基互补配对与靶基因mRNA结合(结合位点一般都位于mRNA的中间部位),导致靶基因mRNA的裂解或降解。
endo-siRNA广泛分布于真核生物中,成簇分布于生物体内的转座子、假基因、基因内含子、外显子以及间隔区等。endo-siRNA是由基因组中的重复序列(转座子或反转座子元件等)编码,其主要功能有:第一,介导移动元件的基因沉默,导致靶基因的甲基化或靶基因mRNA的降解:第二,介导DNA的异染色质化;第三,调控基因组的重排。目前,虽然在endo-siRNA的主要特点及生物学功能上取得了一定的研究进展,仍有许多问题需要深入研究。
三、piRNA的生物合成和作用机制
piRNA是2006年发现的一类与生殖细胞特异性PIWI家族蛋白相互作用,长约26-30 nt的小分子非编码RNA。PIWI蛋白为piRNA生物发生和功能形式所必须,piRNA对生殖细胞干性的维持、配子形成、胚胎发育等多种生殖相关事件至关重要。
piRNA来源于基因组中的piRNA簇或转座子区域,有长的单链转录本切割产生。研究发现,piRNA生成有两条途径:初级生成途径和次级生成途径。与miRNA和siRNA不同的是,piRNA形成途径都不需要Dicer的参与。在初级途径中,长的单链转录本被某种核酸内切酶剪切形成5端带有尿嘧啶的初级piRNA,随后,初级piRNA完成3端的2氧甲基化修饰。研究推测果蝇中PIWI、Aub和Ag03蛋白可能就是这种核酸内切酶。
基于对果蝇和哺乳动物piRNA的分子结构特点,乒乓模型(Ping-pang model)被用来解释piRNA从转录前体物中循环增生。首先,初级piRNA互补的转录产物被PIWI蛋白识别并激活PIWI的内切酶活性,初级piRNA被切割形成一条新的piRNA(次级piRNA)。其次,次级piRNA3端的腺嘌呤与初级piRNA 5端的尿嘧啶互补。同样,次级piRNA也可通过识别和切割互补的RNA序列,产生新的piRNA,即次级生成的乒乓扩增途径。
piRNA的作用机制与miRNA和siRNA不同,miRNA和siRNA主要通过Ago亚家族蛋白相互作用,而piRNA则是通过与PIWI蛋白结合形成piRNA复合物来发挥转录阶段的基因沉默作用,可能取决于piRNA与靶基因mRNA的配对程度及参与调控的其他蛋白質因子等。
四、总结
miRNA、siRNA与piRNA的共性是它们都通过一种序列特异的方式指导效应蛋白结合到基因组位点或靶基因RNA上。同样也存在着区别:第一,发生的生物途径。miRNA是由具有“发夹”二级结构的pre-miRNA经Drosha和Dicer剪切加工形成。siRNA来源与长的dsRNA前体,经Dicer剪切形成。piRNA的生成途径依赖PIWI蛋白,不需要Dicer的参与。第二,功能机制。miRNA是在转录后水平降解或抑制蛋白翻译从而调控基因表达。siRNA主要功能是装载至Ago蛋白介导蛋白编码基因及转座子等沉默。piRNA主要功能是介导生殖细胞内转座子的沉默、生殖细胞的发育及完整等,通过表观遗传调控及转录后水平等发挥基因沉默作用。第三,分布特点。miRNA成簇分布在体细胞和生殖细胞中。endo-siRNA分布在体细胞和生殖细胞中,成簇的分布在动物体内的转座子、假基因、基因内含子、外显子以及间隔区等。piRNA主要分布在生殖细胞中,并主要成簇的分布在基因间隔区。
sncRNA在细胞分化、生物发育及疾病发生、发展过程中的作用拓展和丰富了以往人们对小分子RNA的认识。最新的高通量测序技术,将会使人们对真核生物基因表达调控网络的理解提高到一个新的水平。对sncRNA不断深入的研究,不仅在理论上丰富了我们对基因调控的认识,而且在实际应用中可以使sncRNA成为疾病诊断的新生物学标记和药物靶标,最终为疾病治疗提供新的策略和思路。因此,在《生物化学》的教学中增加sncRNA相关的知识介绍显得尤为重要。我们可以从生物合成和作用机制等方面加强教学,有利于开拓学生的视野、培养学生的创新意识和科学素养,从而为培养高素质的医学创新型人才做出积极的贡献。
