抗、感大豆品种对尖镰孢根腐病菌侵染及毒素的反应
2017-08-09黄铭慧张思奇孙丽萍李永刚
黄铭慧, 张思奇, 孙丽萍, 张 丽, 李永刚
(东北农业大学农学院, 哈尔滨 150030)
抗、感大豆品种对尖镰孢根腐病菌侵染及毒素的反应
黄铭慧, 张思奇, 孙丽萍, 张 丽, 李永刚*
(东北农业大学农学院, 哈尔滨 150030)
为了进一步明确大豆抗感品种对根腐尖镰孢的抗病机制,通过透射电镜和形态观察分析了尖镰孢(M38)侵染及产生的毒素对抗感大豆品种的影响。研究表明大豆幼苗胚根经粗毒素处理后,抗病品种‘东农56’幼苗胚根生长没有显著差别,但显著抑制了感病品种‘黑农53’胚根的伸长和侧根的生长;尖镰孢毒素高浓度和低浓度(V粗毒素∶V无菌水=1∶0和1∶15)对抗感品种的致萎能力无显著差异,高浓度处理平均萎蔫指数为100,低浓度处理为17.9~18.1左右;但中等浓度(V粗毒素∶V无菌水=1∶1,1∶5,1∶10)对于感病品种的致萎作用更大;同时,发现尖镰孢侵染感病大豆品种根内侵染量要明显多于抗病品种,且感病品种组织中菌丝的直径明显大于抗病品种,都出现很明显的质壁分离和细胞壁加厚的现象。
大豆根腐病; 尖镰孢; 毒素; 萎蔫指数; 抗病机制
大豆镰孢根腐病是大豆主要病害之一,而尖镰孢Fusariumoxysporum是大豆根腐镰孢菌的优势种群之一,是一种有较强寄生能力、毁灭性强和分布广泛的土壤习居菌,给我国大豆生产带来了严重威胁[1]。
研究者普遍认为毒素是尖镰孢主要致病因子之一,其中对镰刀菌酸(fusaric acid, FA)的研究居多。FA可以损伤寄主植物根系,致使其水分吸收受阻及生长受到抑制,并产生萎蔫[2-3]。Marre等和Bouizgarne等研究认为FA可抑制植物根部的生长[4-5]。李赤等研究香蕉枯萎病菌毒素时认为FA是引起香蕉枯萎症状的主要物质,其毒害作用随其浓度升高而增强[6],而曹永军等认为FA也是导致香蕉枯萎病菌致病力出现分化的主要原因之一[7]。经粗毒素处理的棉花和香蕉苗,均出现了胶状物或侵填体堵塞导管的维管束病理变化,产生叶片黄化、倒垂、维管组织褐变等典型枯萎病病害症状[8-9]。 而在大豆尖镰孢根腐病的研究中,台莲梅等和宋以星等研究认为尖镰孢毒素能够抑制大豆胚根生长,对大豆幼苗有致萎作用[10-11]。但大豆镰孢根腐病的抗感品种根尖镰孢毒素的致萎反应是否有差别,还未见报道。
关于尖镰孢侵入途径和扩展方面的研究中,有研究者认为尖镰孢可能是从伤口或侧根形成点侵入,在根皮层细胞间生长或者通过木质部纹孔侵入导管[12]。Lagopodi等用绿色荧光蛋白(GFP)基因标记番茄尖镰孢,发现菌丝先扩展到根毛,从根毛侵入根部并大量繁殖,随后进入维管束,形成分生孢子,导致根出现腐烂现象[13]。Ramirez 等用尖镰孢接种苜蓿,2 d病菌侵入须根,3~4 d侵入皮层,并沿着根轴向茎部生长,9~10 d到茎中心,在导管中扩展,皮层和茎部充满菌丝[14]。
尖镰孢的研究报道虽然较多,在根腐尖镰孢侵入及毒素对抗感大豆品种根系的影响仍缺少相关报道,这些对于我们进一步了解大豆品种对尖镰孢的抗病机制是十分必要的。因此,本文拟运用透射电镜及形态观察对根腐尖镰孢侵入及毒素对抗感尖镰孢根腐病的大豆品种根系等进行抗病机制方面的研究。
1 材料与方法
1.1 材料
供试菌株:尖镰孢(M38)由本课题组分离并鉴定。
供试品种:抗病品种 ‘东农56’和感病品种‘黑农53’由大豆省部共建生物学教育部重点实验室提供,经过本课题组鉴定确定其对尖镰孢(M38)的抗感反应[1]。
1.2 大豆尖镰孢(M38)毒素的提取及毒性测定
1.2.1 尖镰孢(M38)毒素的提取与纯化
大豆尖镰孢毒素的提取与纯化采用炭吸附法[15]。将菌株接种在PSA培养液(100 mL/500 mL三角瓶)中,共接3瓶,置25~28℃,培养15 d后过滤。于培养液中加5%的活性炭置于5~6℃冰箱内过夜,用滤纸过滤炭末,然后将炭末浸泡于10倍体积的热甲醇中,去除不溶物的氯仿沉淀,移出黄色不溶物,剩下甲醇-氯仿溶解液蒸发至干,加入100 mL无菌水即获得粗毒素液,备用。
1.2.2 粗毒素对大豆发芽及胚根生长的影响
‘东农56’和‘黑农53’种子经70%乙醇表面消毒后,置于清水中浸泡4~5 h,摆于铺有无菌滤纸的磁盘内(35 cm×25 cm),置于25℃恒温箱内培养3 d,取整齐一致的幼根,洗净,表面消毒,然后将幼苗根系浸入V粗毒素∶V无菌水=1∶3的粗毒素溶液中,48 h后取样,测量胚根的长度,主根和须根生长状态。‘东农56’和‘黑农53’分别设毒素处理与不处理的对照,共4个处理,每个处理5株,3次重复,对照用无菌水处理,分析尖镰孢(M38)毒素对大豆抗、感品种胚根生长的影响,以及对毒素的反应是否存在差别。参照陆仕华等[16]报道的抑制胚根生长的测定法,测定其对胚根生长的抑制率。
胚根抑制率(%)=(对照胚根平均长度-处理胚根平均长度)/对照胚根平均长度×100。
1.2.3 粗毒素液对不同品种幼苗的致萎作用
取粗毒素溶液,按体积比分别稀释成V毒素∶V无菌水为1∶0、1∶1、1∶5、1∶10、1∶15等5个梯度浓度,分别处理‘东农56’和‘黑农53’生长一致的株高为10 cm的幼苗,管口用 parafilm封口膜封住,以防水分流失,每管12 mL放1株幼苗,每个浓度处理15株,设无菌水为对照,在常温通风下培养10 d观察活体幼苗的萎蔫程度,参照兀旭辉[17]报道的方法测定并计算萎蔫指数。
萎蔫指数=∑(萎蔫级株数×萎蔫级数)/(试验总株数×最高萎蔫级数)×100。
萎蔫分级标准:0级,叶面无症状;1级,叶片出现失水状,但不萎蔫;2级,叶片出现失水状,但不萎蔫;3级,部分叶片萎垂;4级,叶片均呈暗绿色或暗褐色萎垂;5级,部分叶片枯死。
1.3 大豆尖镰孢(M38)侵染大豆根部后侧根组织结构变化
利用分生孢子悬浮液灌根法(106个孢子/mL)接种生长至约10 cm的幼苗根部,对照不接种,每处理5株,3次重复。培养5 d后取大豆植株侧根组织,样品经戊二醛-锇酸双固定,磷酸缓冲液冲洗,丙酮系列脱水,Epson812 包埋,制成50 nm 超薄切片,再经醋酸铀和柠檬酸铅染色后,透射电子显微镜H-7650拍片。对样品的侧根细胞壁、细胞及亚细胞结构等进行观察,分析尖镰孢侵染大豆根部后侧根组织结构上的变化及尖镰孢在组织中的侵染情况。
1.4 数据分析
采用SPSS 17.0软件进行统计分析,应用Duncan法进行差异显著性检验。
2 结果与分析
2.1 尖镰孢粗毒素对大豆幼苗胚根生长的影响
尖镰孢粗毒素对于大豆抗、感品种的胚根生长都具有一定的抑制作用,抑制率分别为17.39%和29.61%(表1)。用Duncan法进行差异显著性分析表明,V粗毒素∶V无菌水=1∶3粗毒素溶液处理后,抗病品种‘东农56’幼苗胚根生长与对照相比没有显著差异,但感病品种‘黑农53’ 与对照相比,25%粗毒素明显抑制了‘黑农53’胚根的伸长和侧根的生长(图1)。
表1 尖镰孢粗毒素对大豆胚根生长的影响1)
Table 1 Effects of crude toxins ofFusariumoxysporumon soybean radicle growth

品种Variety抗感类型Typeofresistance胚根长度/cmRadiclelength胚根抑制率/%Inhibitionrate‘东农56’对照‘Dongnong56’CK‘东农56’处理‘Dongnong56’treatment抗病R(1.84±0.12)a(1.52±0.10)a17.39‘黑农53’对照‘Heinong53’CK‘黑农53’处理‘Heinong53’treatment感病S(1.52±0.16)a(1.07±0.14)b29.61
1) 表中数据为平均值±标准误,同列数字后不同小写字母表示经邓肯法检验在P<0.05水平差异显著。下同。 Data in the table are mean±SE.Different letters indicate significant difference at 0.05 level by Duncan’s test. The same below.
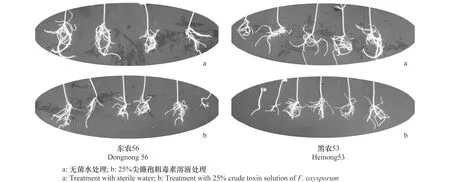
图1 尖镰孢粗毒素对大豆幼苗根部生长的影响Fig.1 Effects of the crude toxins of Fusarium oxysporum on the growth of soybean seedling roots

品种Variety粗毒素溶液浓度(V粗毒素∶V无菌水)Concentrationofcrudetoxin萎蔫指数Wiltingindex东农56Dongnong561∶0(100.0±0.0)a黑农53Heinong531∶0(100.0±0.0)a东农56Dongnong561∶1(86.2±1.3)c黑农53Heinong531∶1(91.6±1.8)b东农56Dongnong561∶5(69.1±1.1)e黑农53Heinong531∶5(80.2±2.3)d东农56Dongnong561∶10(37.7±2.0)g黑农53Heinong531∶10(44.6±1.2)f东农56Dongnong561∶15(17.9±2.4)h黑农53Heinong531∶15(18.1±0.7)h东农56Dongnong560(0.0±0.00)i黑农53Heinong530(0.0±0.00)i
2.2 尖镰孢粗毒素对大豆幼苗的致萎作用
将尖镰孢粗毒素配制成一定浓度,处理大豆抗、感品种幼苗。从表2可以看出,随着浓度的增加,粗毒素溶液对大豆幼苗的致萎作用逐渐增强,高浓度和低浓度(V粗毒素∶V无菌水=1∶0和1∶15)的致萎能力在抗、感品种之间无显著差异,萎蔫指数分别为100和17.9~18.1;但V毒素∶V无菌水1∶1、1∶5、1∶10对感病品种的致萎作用明显大于抗病品种(表2)。
2.3 大豆尖镰孢接种大豆植株后侧根组织结构的变化
利用电镜对接种与未接种尖镰孢的大豆抗、感品种植株的侧根细胞壁、细胞和亚细胞结构及尖镰孢菌丝的侵染情况进行观察。从图2~3可以看出,感病品种‘黑农53’侧根受侵染后的含菌量远高于抗病品种‘东农56’,且抗病品种中的菌丝直径明显比感病品种的细,均出现了明显的质壁分离和细胞壁加厚的现象,被侵染的侧根组织细胞壁发生形变,细胞内部结构紊乱。同时,在未接种的抗感品种中也出现在少量的菌丝,可能为内生真菌的侵染造成的,但总体上能看得出在尖镰孢侵染前后,抗感品种中菌丝侵染和扩展方面有明显差别。

图2 接种尖镰孢后‘东农56’的侧根形态Fig.2 Morphologic observation on fibrous roots of ‘Dongnong 56’ inoculated with Fusarium oxysporum

图3 接种尖镰孢后‘黑农53’的侧根形态Fig.3 Morphologic observation on fibrous roots of ‘Heinong 53’ inoculated with Fusarium oxysporum
3 讨论
尖镰孢和寄主植物之间的互作是一种十分复杂的机理,贯穿病菌与寄主之间的识别、病原菌的入侵、定殖,寄主产生的防御反应及病原菌克服防御机制等整个过程,涉及组织、细胞、生化、分子等水平及许多复杂因子的共同调控[18]。
本研究提取大豆根腐尖镰孢(M38)的粗毒素,并测定其对抗感大豆品种的致病作用,发现尖镰孢粗毒素对大豆抗、感品种的胚根生长都有明显的抑制作用,这与台莲梅等和宋以星等认为尖镰孢毒素能够抑制大豆胚根的生长,损伤大豆根部细胞,对大豆幼苗有一定的致萎作用的研究结果是一致的[10-11]。但本研究中也显示,25%的粗毒素溶液对大豆抗病品种‘东农56’的幼苗根系没有影响,而对感病品种‘黑农53’影响较大,明显抑制了胚根的伸长和侧根的生长(图1)。在致萎方面的测定中,随着粗毒素溶液浓度的增加,粗毒素溶液对大豆幼苗的致萎作用逐渐增强,当尖镰孢毒素溶液为高浓度和低浓度(V粗毒素∶V无菌水=1∶0和1∶15)时,对大豆抗感品种幼苗的致萎能力基本一致,这两个浓度在实际尖镰孢侵染过程中是达不到的,而V毒素∶V无菌水为1∶1、1∶5、1∶10时对感病品种的致萎作用明显大于抗病品种(表2),这说明尖镰孢侵染达到一定量,就可分泌足够多的毒素使抗感品种表现出不同反应。这验证了易海艳等建议用镰刀菌酸筛选抗枯萎病品种的方法是可行的[20],但仍需要做大量工作去把这项工作具体化。从本研究可以看出,毒素仅影响到根系的生长和植株出现的萎蔫,没有出现明显的根腐症状,也就是说,毒素仅是尖镰孢致病因素之一。
Lagopodi等和Ramírez-Suero等分别对番茄和苜蓿接种尖镰孢后进行了根部细胞侵染位置的观察,在根部表皮细胞、维管束、木质部、皮层和茎部均发现了菌丝大量侵入和聚集的现象[13-14]。而本研究对接种尖镰孢后经电镜样品处理的大豆植株侧根样品组织,进行侧根细胞壁、细胞、亚细胞结构及尖镰孢菌丝的侵染位置的观察,发现大豆侧根细胞间和细胞内都有尖镰孢菌体的侵入,而且使细胞内部发生严重的质壁分离现象。这与马立功等[19]的研究基本是一致的,他认为大豆幼根细胞膜对尖镰孢毒素较为敏感,随着毒素浓度的增加,对细胞膜的破坏作用也随之增强,从而导致细胞内电解质离子的异常外渗、质膜内陷、局部断裂、膜周围出现电子密集的沉淀物、细胞发生质壁分离、细胞膜破裂和电解质外流等现象。虽然这方面有研究报道,但针对大豆根腐尖镰孢抗感品种对比这方面的差异未见报道。本研究表明抗病品种与感病品种侵入菌丝的量相差较大,感病品种有大量菌丝侵入,菌丝较粗大,而抗病品种菌丝相对较少,菌丝较细,这也是抗病品种发病较轻的主要原因之一。这与上述毒素处理后,当毒素达到一定程度后,抗感品种表现的萎蔫反应出现差别的原因相吻合,因为感病品种菌丝侵染量大,大量菌丝产生的毒素量自然也就增大,导致感病品种比抗病品种更容易、更快地表现出萎蔫。
综上所述,本研究综合评价了大豆根腐尖镰孢的侵染及产生的毒素对大豆抗感品种根系的影响,为进一步揭示大豆抗、感品种对根腐尖镰孢的抗性机理提供了依据。
[1] 张丽, 耿肖兵, 王春玲, 等. 黑龙江省大豆镰孢根腐病菌鉴定及致病力分析[J]. 植物保护, 2014, 40(3): 165-168.
[2] 叶旭红, 林先贵, 王一明. 尖镰孢致病相关因子及其分子生物学研究进展[J]. 应用与环境生物学报, 2011, 17(5): 759-762.
[3] Bani M, Rispail N, Evidente A, et al. Identification of the main toxins isolated fromFusariumoxysporumf.sp.pisirace 2 and their relation with isolates pathogenicity[J]. Journal of Agricultural and Food Chemistry, 2014,62(12): 2574-2580.
[4] Bouizgarne B, Brault M, Pennarun A M, et al. Electrophysiological responses to fusaric acid of root hairs from seedlings of date palm-susceptible and-resistant toFusariumoxysporumf.sp.albedinis[J]. Journal of Phytopathology, 2004, 152: 321-324.
[5] Marre M T, Vergani P, Albergoni F G.Relationship between fusaric acid uptake and its binding to cell structures in leaves ofEgeriadensaand its toxic effects on membrane permeability and respiration [J]. Physiological and Molecular Plant Pathology, 1993, 42(2): 141-157.
[6] 李赤, 黎永坚, 于莉, 等. 香蕉枯萎病菌毒素的成分分析及其生物测定[J]. 果树学报, 2010, 27(6): 969-974.
[7] 曹永军, 程萍, 喻国辉, 等. 香蕉枯萎病菌菌株致病力分化及其原因研究[J]. 热带作物学报, 2011, 32(8): 1532-1536.
[8] 袁红旭, 商鸿生. 棉花枯萎病菌接种及粗毒素处理后棉苗维管束病理特征[J]. 植物病理学报, 2002, 32(1): 17-20.
[9] 许文耀, 兀旭辉, 杨静惠, 等. 香蕉假茎细胞对香蕉枯萎病菌小同小种及其粗毒素的病理反应[J]. 植物病理学报, 2004, 34(5): 425-430.
[10]台莲梅, 许艳丽, 闫凤云. 尖孢镰刀菌毒素对大豆胚根组织影响的超微结构研究[J].植物病理学报,2006,36(6): 512-516.
[11]宋以星,杜开书,石明旺.镰刀菌毒素诱导大豆对根腐病菌的抗性研究[J].河南科技学院学报(自然科学版),2011,39(1):38-43.
[12]王建明, 郑经武, 贺运春. 西瓜枯萎病田间症状类型及侵染来源的研究[J]. 山西农业大学学报, 1992, 12(4): 352-354.
[13]Lagopodi A L, Ram A F, Lamers G E, et al. Novel aspects of tomato root colonization and infection byFusariumoxysporumf.sp.radicis-lycopersicirevealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker [J]. Molecular Plant-Microbe Interactions, 2002, 15(2): 172-179.
[14]Ramirez-Suero M, Khanshour A, Martinez Y.A study on the susceptibility of the model legume plantMedicagotruncatulato the soil-borne pathogenFusariumoxysporum[J]. European Journal of Plant Pathology, 2010, 126(4): 517-530.
[15]台莲梅, 许艳丽, 高凤昌. 尖镰孢毒素的初步研究[J]. 黑龙江八一农垦大学学报, 2004, 16(4): 9-12.
[16]陆仕华.水稻尾孢霉毒素[J].真菌学报,1985,4(4):240-254.
[17]兀旭辉. 香蕉枯萎病菌粗毒素特性及其在抗病突变体筛选中的应用[D]. 福州: 福建农林大学, 2005.
[18]裴月令, 曾凡云, 彭军, 等. 尖镰孢与寄主互作机理研究进展[J]. 热带生物学报, 2014, 5(1): 92-100.
[19]易海艳, 查向浩, 马刘峰. 镰刀菌酸及其枯萎病菌培养液对棉花的毒性[J]. 安徽农业科学, 2011, 39(12): 7038-7039.
[20]马立功, 张匀华, 季宏平, 等. 大豆根腐主要病原菌毒素致病作用的初步研究[J]. 中国农学通报, 2010, 26(6): 243-246.
(责任编辑:田 喆)
Response of resistant and susceptible soybean varieties to toxins and invasion ofFusariumoxysporumcausing soybean root rot
Huang Minghui, Zhang Siqi, Sun Liping, Zhang Li, Li Yonggang
(CollegeofAgriculture,NortheastAgriculturalUniversity,Harbin150030,China)
In order to further clarify the resistance mechanisms of resistant and susceptible soybean varieties againstFusariumoxysporumcausing soybean root rot, the toxic and invasive effects ofF.oxysporumwere analyzed by using TEM and toxins ofF.oxysporum. The results showed that there were no significant differences between treatment and control of the resistant soybean variety ‘Dongnong 56’ with the soybean seedling roots immersed in the toxin solution ofF.oxysporum. It was obvious that radicle elongation and lateral root growth of the susceptible variety ‘Heinong 53’ were inhibited. At the same time, the wilting ability of high (original fluid of crude toxin) and low (Vcrude toxin∶Vsterile water=1∶15) concentrations of toxins on resistant and susceptible varieties was no obvious differences, with an average wilting index of 100 and 18, respectively. However, it could cause more serious wilt in susceptible varieties than resistant varieties at the moderate concentration of toxins fromF.oxysporum(Vcrude toxin∶Vsterile water=1∶1, 1∶5 and 1∶10). Observing the soybean roots after inoculation by TEM, we found that the amount and diameter of fungal mycelia of the susceptible variety ‘Heinong 53’ was thicker than those of the resistant variety ‘Dongnong 56’, and the two varieties both had obvious plasmolysis and cell wall thickening.
soybean root rot;Fusariumoxysporum; toxin; wilting index; resistance mechanisms
2016-08-10
2016-09-18
S 435.651
A
10.3969/j.issn.0529-1542.2017.04.014
* 通信作者 E-mail:neaulyg@126.com
