A型产气荚膜梭菌噬菌体裂解酶Cp51的原核表达及活性检测
2017-08-07吕梦娜龙航宇王丽梅陈冰冰林瑞庆
吕梦娜,龙航宇,王丽梅,唐 陶,陈冰冰,林瑞庆
(华南农业大学 兽医学院/广东省动物源性人兽共患病预防与控制重点实验室,广东 广州510642)
A型产气荚膜梭菌噬菌体裂解酶Cp51的原核表达及活性检测
吕梦娜,龙航宇,王丽梅,唐 陶,陈冰冰,林瑞庆
(华南农业大学 兽医学院/广东省动物源性人兽共患病预防与控制重点实验室,广东 广州510642)
【目的】原核表达A型产气荚膜梭菌噬菌体裂解酶Cp51并研究其对A型产气荚膜梭菌Clostridium perfringens的体外抗菌活性。 【方法】合成噬菌体裂解酶Cp51基因,并构建了pET-32a-Cp51原核表达载体,转化到大肠埃希菌Escherichia coli BL21(DE3),经终浓度为0.5 mmol·L–1的IPTG诱导以及镍柱纯化后,获得了可溶性的Cp51重组蛋白;用比浊法检测Cp51重组蛋白的杀菌活性。 【结果】噬菌体裂解酶Cp51重组蛋白能够有效降低7株A型产气荚膜梭菌的浊度,裂解酶质量浓度在5 μg·mL–1以上、作用30 min对A型产气荚膜梭菌的杀菌率可达到99.99%以上,而对其他种类细菌无杀菌效果。 【结论】A型产气荚膜梭菌噬菌体裂解酶Cp51重组蛋白对
A型产气荚膜梭菌有较强的体外杀菌活性和特异性,研究结果为后续裂解酶Cp51的临床应用奠定基础。
产气荚膜梭菌;噬菌体;裂解酶;蛋白表达;杀菌活性
产气荚膜梭菌Clostridium perfringens又名魏氏梭菌C. welchii,是一种重要的人畜共患病原菌[1]。产气荚膜梭菌为两端钝圆厌氧性菌,呈粗杆状,单个或成对存在,是一种能产芽孢的革兰阳性菌,在人和动物的肠道内普遍存在,也广泛分布于污水、土壤等自然环境中。根据产气荚膜梭菌产生的4种主要外毒素,可将产气荚膜梭菌分为A、B、C、D和E 型,其中A型产气荚膜梭菌是引发人类气性坏疽及家禽坏死性肠炎的主要病原[2]。目前家禽坏死性肠炎的预防和治疗仍以抗生素为主,但由于抗生素的长期和不规范使用引起细菌耐药性及药物残留等问题广泛出现,不仅使家禽的生产性能大大降低,而且通过鸡等肉制品威胁人类健康[3-4]。人类已进入了“后抗生素”时代,细菌耐药性等问题已经成为现代医学以及畜牧生产的难题[5-6]。作为预防和治疗细菌疾病的一种古老的方法,噬菌体防控在这种情况下又回到了历史舞台。噬菌体在抗生素兴起之前就已经被发现和使用,噬菌体能特异性地裂解宿主细菌,其编码的裂解酶是在基因组复制后期合成的一种酶,作用于细菌细胞壁肽聚糖上的酰胺键,使细菌破碎,释放出子代噬菌体[7]。细菌对噬菌体裂解酶不易产生耐药性,而噬菌体裂解酶的结合结构域能识别宿主细胞壁受体分子,从而从体外特异性识别细菌并将其溶解,而不需噬菌体侵染细菌时的吸附、自我复制、繁殖等系列过程,因此,噬菌体裂解酶的裂解谱比噬菌体的裂解谱宽。噬菌体裂解酶作为一种潜在的抗菌制剂具有高效、特异性强等优点[8-9]。目前国内已有许多学者利用相应的噬菌体裂解酶作用于葡萄球菌、链球菌等细菌均得到了较好的抗菌效果[10-13]。产气荚膜梭菌病的防控及噬菌体研究在国外受到重视[14],但国内目前针对产气荚膜梭菌噬菌体裂解酶的相关研究鲜见报道。
本试验原核表达了 A 型产气荚膜梭菌噬菌体裂解酶 Cp51,并初步研究了 A 型产气荚膜梭菌噬菌体裂解酶 Cp51 重组蛋白对 A 型产气荚膜梭菌的体外裂解活性,以期为进一步研究治疗坏死性肠炎的新型药物奠定基础。
1 材料与方法
1.1 供试材料
1.1.1 菌株和质粒 7株A型产气荚膜梭菌、2株金黄色葡萄球菌Staphylococcus aureus、2株猪链球菌Streptococcus suis、1株肠炎沙门氏菌Salmonella enteritidis、1株大肠埃希菌Escherichia coli和1株腐败梭菌Clostridium septicum及表达载体pET-32a(+)为华南农业大学寄生虫实验室保存;感受态细胞DH 5α、BL 21(DE3)均购自北京全式金生物技术有限公司。
1.1.2 工具酶及主要试剂 BamH I内切酶、HindⅢ内切酶、DNA Marker DL 2000、DL 15000、凝胶回收试剂盒均购自日本TaKaRa公司。Page Ruler Prestained Protein Ladder 10~170 kU购自Thermo公司。质粒抽提试剂盒购自天根生化科技有限公司。BCA蛋白浓度测定试剂盒、纯化填料、HRP羊抗鼠IgG、EasySee Western-blot Kit购自北京全式金生物技术有限公司。LB肉汤、TSC培养基、咪唑购自广州鼎国生物技术有限公司。其余常用试剂为国产或进口分析纯。
1.2 试验方法
1.2.1 A型产气荚膜梭菌噬菌体裂解酶Cp51基因的克隆及表达载体的构建 通过GeneBank获得A型产气荚膜梭菌噬菌体裂解酶Cp51蛋白序列(AGH27916.1),根据Cp51蛋白的氨基酸序列人工合成相应的Cp51基因序列,在5′端引入BamH I位点,3′端引入Hind Ⅲ位点,并在不改变氨基酸的情况下对Cp51基因序列中810位碱基由A突变为G,人工合成由深圳华大科技有限公司完成。合成的基因克隆于pMD-18T载体上,转化至DH5α感受态大肠埃希菌中,挑取阳性克隆,进行PCR、酶切、测序验证。重组质粒pMD-18T-Cp51经BamH I和HindⅢ双酶切,切胶回收Cp51基因目的片段,克隆到表达载体pET-32a(+)上,构建重组表达载体pET-32a-Cp51,阳性克隆送到深圳华大科技有限公司测序。
1.2.2 重组蛋白pET-32a-Cp51的表达和纯化 将测序正确的表达质粒转化到BL 21(DE3)感受态细胞内,挑取单克隆接种到5 mL含有100 μg·mL–1的Amp+LB液体培养基中,37 ℃条件下150 r·min–1培养过夜,按1︰100的比例扩大培养,37 ℃条件下180 r·min–1震荡培养至D600nm为0.4~0.6,加入IPTG使终浓度为0.5 mmol·L–1,37 ℃条件下180 r·min–1诱导3 h。8 000 r·min–1离心10 min,收集菌体,菌体重悬于平衡缓冲液中(10 mmol·L–1咪唑),功率200 W,超声3 s,间隔5 s,超声120次。分别取上清和沉淀,加入5×SDS-PAGE上样缓冲液10 μL,煮沸5 min,10 000 r·min–1离心10 min,进行SDSPAG 电泳,检测重组蛋白是存在于上清液中还是存在于包涵体中。
采用全式金生物技术有限公司的Ni2+–NTA蛋白质纯化柱纯化目的蛋白, 选择不同浓度的咪唑(20~200 mmol·L–1)梯度洗脱,纯化后的蛋白进行SDS-PAGE电泳分析,测定纯化效率,确定洗脱液的最佳咪唑浓度,并用BCA法测定蛋白浓度。
1.2.3 重组蛋白pET-32a-Cp51 的 Western blot检测 分别取150 mmol·L–1咪唑洗脱液40 μL,pET-32a-Cp51未诱导样品和pET-32a(+)诱导菌处理后加 5×SDS-PAGE上样缓冲液10 μL,进行SDSPAGE电泳。电泳结束后将胶切成合适大小。安装好转膜装备,放入转膜槽,接通电源,电流150 mA,转膜2 h,质量分数5%脱脂奶粉封闭1 h,小鼠抗His抗体用质量分数为5% 的脱脂奶粉稀释为体积比1︰2 000,室温条件下孵育2 h,羊抗小鼠HRPIgG用质量分数为5% 的脱脂奶粉稀释为体积比1︰2 000,室温条件下摇床孵育1~2 h。TBST洗膜3次,将ECL的2种发光液按照体积比1︰1的比例混合,发光显色,拍照保存。
1.2.4 重组蛋白pET-32a-Cp51活性检测及裂解谱的测定 将过夜培养的A型产气荚膜梭菌以10 000 r·min–1离心1 min,PBS清洗后,细菌重悬于4 mL的 PBS 中,加入1 mL蛋白纯化液,使得最终的反应体系为5 mL,蛋白的终质量浓度分别为 20、10、5和 1 μg·mL–1,同时A型产气荚膜梭菌最终的D600nm约为0.6,细菌数约为108cfu·mL–1,设置pET-32a空载组和PBS组作为对照组,37 ℃条件下180 r·min–1震荡培养,分别在0、5、10、15、20 和30 min时测量D600nm,取3次重复数据的平均值。并将作用30 min的最终反应液取100 μL涂布于TSC平板,40 ℃条件下厌氧培养24 h。
用终质量浓度为10 μg·mL–1重组蛋白pET-32a-Cp51分别作用于本试验分离和保存的7株A型产气荚膜梭菌、2株葡萄球菌、2株链球菌、1株沙门氏菌、1株大肠埃希菌和1株腐败梭菌,测定其在作用0、5、10、15、20和30 min时的D600nm。
2 结果与分析
2.1 Cp51基因原核表达载体的构建与鉴定
将阳性重组表达载体pET-32a-Cp51经BamH I和Hind Ⅲ双酶切后,得到2条片段,一条为载体条带,另一条约为1 150 bp的目的基因条带(图1),测序结果表明重组表达质粒构建正确。
2.2 pET-32a-Cp51诱导表达及纯化
将重组的质粒pET-32a-Cp51转化至BL 21(DE3)后,经IPTG诱导表达,分别取上清和沉淀进行SDS-PAGE分析,结果显示上清和沉淀中均在相对分子质量约为63 000处有明显蛋白条带(图2)。故选择从上清中纯化目的蛋白,用不同浓度的咪唑溶液洗脱收集,SDS-PAGE检测显示用150和200 mmol·L–1咪唑洗脱时,在相对分子质量63 000处可获得较多的重组蛋白,BCA试剂盒测定蛋白质量浓度分别是555和450 μg·mL–1(图3)。

图1 重组质粒pET-32a-Cp51酶切鉴定Fig. 1 Identification of recombinant plasmid pET-32a-Cp51 by enzyme digestion

图2 pET-32a-CP51表达SDS-PAGE电泳Fig. 2 SDS-PAGE pattern of expressed pET-32a-CP51 protein
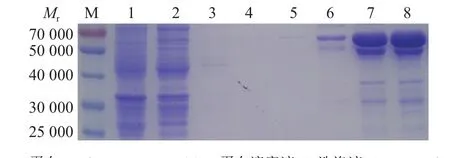
图3 pET-32a-Cp51蛋白纯化的SDS-PAGE电泳Fig. 3 SDS-PAGE pattern of purified pET-32a-Cp51 protein
2.3 pET-32a-Cp51的Western blot检测结果
pET-32a-Cp51纯化产物和pET-32a空载体表达产物经SDS-PAGE电泳后的蛋白条带转移到硝酸纤维素膜上进行免疫印迹,结果纯化的重组目的蛋白和空载体表达产物均可与小鼠抗His抗体发生反应,纯化的重组目的蛋白约在相对分子质量63 000的位置出现特异性反应条带,pET-32a空载体表达产物约在相对分子质量20 000的位置出现特异性反应条带(图4)。
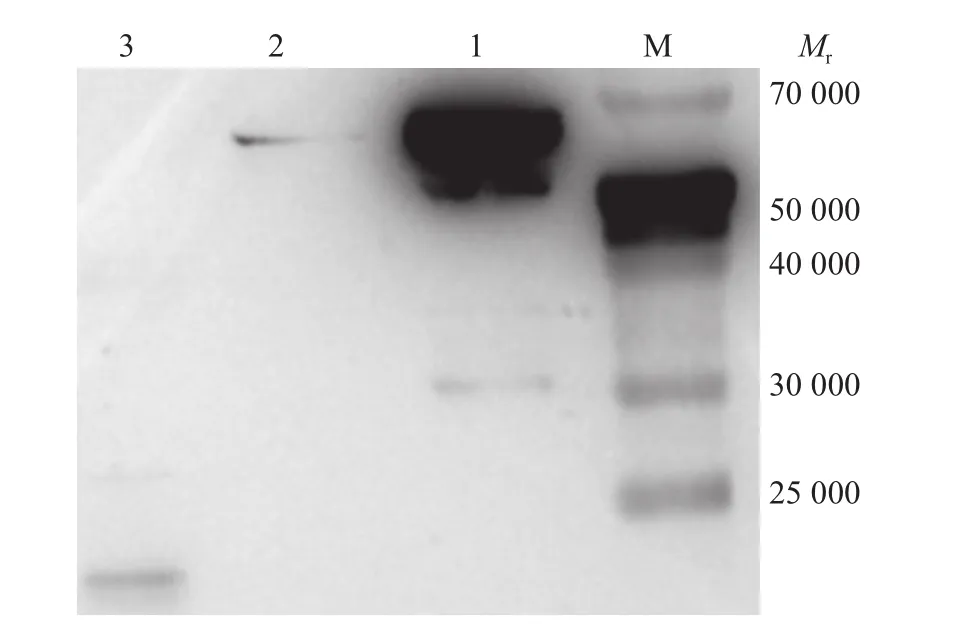
图4 pET-32a-Cp51蛋白的Western blot检测结果Fig. 4 Western blot results of pET-32a-Cp51 protein
2.4 pET-32a-Cp51活性及裂解谱检测结果
采用比浊法,测定pET-32a-Cp51的抗菌活性。结果(图5)显示,与pET-32a(+)和PBS对比,30 min内终质量浓度为20、10和5 μg·mL–1的pET-32a-Cp51噬菌体裂解酶稀释液能将A型产气荚膜梭菌的浊度从0.6降至0.1,而终质量浓度1 μg·mL–1的pET-32a-Cp51噬菌体裂解酶稀释液能将A型产气荚膜梭菌的浊度从0.6降至0.33。说明pET-32a-Cp51 作用于相近起始D600nm的A型产气荚膜梭菌液时,随着酶液作用浓度的增加,D600nm下降速度加快。平板涂布样品经40 ℃条件下培养24 h后观察,根据平板菌落计数结果,发现终质量浓度为20、10和5 μg·mL–1酶液作用于菌液30 min后,可使A型产气荚膜梭菌裂解率达99.99%、99.99%和 100%,酶液终质量浓度为1 μg·mL–1样品的裂解率可达99.91%,表明pET-32a-Cp51对A型产气荚膜梭菌具有高效裂解活性,终质量浓度5 μg·mL–1以上有较强杀菌效果。利用相同方法作用于A型产气荚膜梭菌不同分离株发现pET-32a-Cp51均有良好裂解活性(图6)。而作用于葡萄球菌、链球菌、沙门氏菌、大肠埃希菌和腐败梭菌则无裂解作用(图7)。

图5 不同质量浓度pET-32a-Cp51裂解酶对A型产气荚膜梭菌的活性Fig. 5 Activity of different concentrations of pET-32a-Cp51 lysin against type A Clostridium perfringens

图6 pET-32a-Cp51裂解酶对A型产气荚膜梭菌不同分离株的活性Fig. 6 Activity of pET-32a-Cp51 lysin against different isolates of type A Clostridium perfringens
3 讨论与结论
自人类使用抗生素以来,各种抗生素的滥用、乱用现象层出不穷,因此导致了大量的细菌耐药和抗生素残留等问题。由于抗生素的使用不当带来的耐药菌株甚至成为超级耐药菌株,不仅提高了畜禽的死亡率,而且增大了预防和治疗的投入,给养殖业带了巨大的损失[15]。随着产气荚膜梭菌耐药性加剧,开发新型抗菌药物治疗坏死性肠炎迫在眉睫[16]。裂解酶是噬菌体在裂解末期表达的一种能降解肽聚糖的酶,在革兰阳性菌上作用明显[17],还具有高效、特异性强、不易产生抗性等优势,作为新兴的抗菌剂具有较高的应用价值。本研究成功构建A型产气荚膜梭菌的噬菌体裂解酶Cp51 基因的重组表达载体pET-32a-Cp51,经IPTG 诱导表达,并进行蛋白纯化。比浊法测定结果显示A型产气荚膜梭菌的噬菌体裂解酶pET-32a-Cp51 对A型产气荚膜梭菌杀菌活性效果明显,终质量浓度5 μg·mL–1以上、作用0.5 h后的杀菌率高达99.99%,杀菌效果与Gervasi 等[17]的裂解酶试验结果相似,且能以较低的蛋白浓度达到较强的杀菌效果,此外,本试验进行了噬菌体裂解酶pET-32a-Cp51 对多种常见细菌的裂解谱试验,结果表明pET-32a-Cp51 具有较强特异性,只裂解A型产气荚膜梭菌,对其他种类细菌无作用。
近几年,人们应用噬菌体及其裂解酶进行了一系列的试验,显示其具有高效杀菌、不易产生耐药性和特异性强等优点,在治疗细菌感染病方面具有潜在应用价值[18-20]。综上所述,A型产气荚膜梭菌噬菌体裂解酶Cp51原核表达纯化后,其蛋白纯度较高,体外裂解试验表明对A型产气荚膜梭菌具有明显杀灭效果,为研究其替代抗生素作为新型抗菌药物奠定了基础。
[1]GARCIA J P, BEINGESSER J, FISHER D J, et al. The effect of Clostridium perfringens type C strain CN3685 and its isogenic beta toxin null mutant in goats[J]. Vet Microbiol, 2012, 157(3/4): 412-419.
[2]LOVLAND A, KALDHUSDAL M. Severely impaired production performance in broiler flocks with high incidence of Clostridium perfringens-associated hepatitis[J]. Avian Pathology, 2001, 30(1): 73-81.
[3]汪复, 朱德妹, 胡付品, 等. 2012年中国CHINET细菌耐药性监测[J]. 中国感染与化疗杂志, 2013, 13(5): 321-330.
[4]FILE T M, Jr, SRINIVASAN A, BARTLETT J G. Antimicrobial stewardship: Importance for patient and public health[J]. Clin Infect Dis, 2014, 59(S3): S93-S96.
[5]FISCHETTI V A. Bacteriophage endolysins: A novel anti-infective to control gram-positive pathogens[J]. Int J Med Microbiol, 2010, 300(6): 357-362.
[6]SCHRAG S J, MCGEE L, WHITNEY C G, et al. Emergence of Streptococcus pneumoniae with veryhigh-level resistance to penicillin[J]. Antimicrob Agents Chemother, 2004, 48(8): 3016-3023.
[7]SCHUCH R, PELZEK A J, RAZ A, et al. Use of a bacteriophage lysin to identify a novel target for antimicrobial development[J]. PLoS One, 2013, 8(4): e60754.
[8]SCHUCH R, NELSON D, FISCHETTI V A. A bacteriolytic agent that detects and kills Bacillus anthracis[J]. Nature, 2002, 418(6900): 884-889.
[9]LOESSNER M J. Bacteriophage endolysins: Current state of research and applications[J]. Curr Opin Microbiol, 2005, 8(4): 480-487.
[10]李跃, 宫鹏娟, 夏斐斐, 等. 金黄色葡萄球菌噬菌体裂解酶LysK的表达及其多克隆抗体的制备[J] .中国兽医学报, 2014, 34(1): 45-49.
[11]王彬 金.黄色葡萄球菌噬菌体裂解酶LysGH15外用制剂的初步研制[D]. 长春: 吉林大学, 2015.
[12]陈蔚青, 王晓枫, 王普, 等. 链球菌噬菌体裂解酶在大肠杆菌中的表达、纯化及活性检测[J]. 生物工程学报, 2009(8): 1267-1272.
[13]吴蒙, 陆海荣, 黄青山. 金黄色葡萄球菌噬菌体裂解酶Ply187的CHAP结构域的表达及抗菌活性分析[J]. 生物技术通报, 2016, 32(9): 232-238.
[14]GERVASI T, HORN N, WEGMANN U, et al. Expression and delivery of an endolysin to combat Clostridium perfringens[J]. Appl Microbiol Biotechnol, 2014, 98(6): 2495-2505.
[15]MARTIN H, WILLEY B, LOW D E, et al. Characterization of Clostridium difficile strains isolated from patients in Ontario, Canada, from 2004 to 2006[J]. J Clin Microbiol, 2008, 46(9): 2999-3004.
[16]LAWRENCE R, JEYAKUMAR E. Antimicrobial resistance: A cause for global concern[J]. BMC Proc, 2013, 7(S3): S1.
[17]BORYSOWSKI J, WEBER-DABROWSKA B, GÓRSKI A. Bacteriophage endolysins as a novel class of antibacterial agents[J]. Exp Biol Med (Maywood), 2006, 231(4): 366-377.
[18]GU J, XU W, LEI L, et al. LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection[J]. J Clin Microbiol, 2011, 49(1): 111-117.
[19]SCHMELCHER M, POWELL A M, BECKER S C, et al. Chimeric phage lysins act synergistically with lysostaphin to kill mastitis-causing Staphylococcus aureus in murine mammary glands[J]. Appl Environ Microbiol, 2012, 78(7): 2297-2305.
[20]PASTAGIA M, EULER C, CHAHALES P, et al. A novel chimeric lysin shows superiority to mupirocin for skin decolonization of methicillin-resistant and -sensitive Staphylococcus aureus strains[J]. Antimicrob Agents Chemother, 2011, 55(2): 738-744.
【责任编辑 李晓卉】
Prokaryotic expression and activity detection of bacteriophage lysin Cp51 against Clostridium perfringens type A
LÜ Mengna, LONG Hangyu, WANG Limei, TANG Tao, CHEN Bingbing, LIN Ruiqing
(College of Veterinary Medicine, South China Agricultural University/Key Laboratory of Zoonosis Prevention and Control of Guangdong Province, Guangzhou 510642, China)
【Objective】To construct a prokaryotic expression system of type A Clostridium perfringens phage lysin Cp51, and study its antibacterial activity against C. Perfringens type A in vitro. 【Method】Bacteriophage lysin Cp51 gene was synthesized. The prokaryotic expression vector pET-32a-Cp51 was constructed and transformed into Escherichia coli BL21(DE3). After induction using 0.5 mmol·L–1IPTG, the soluble recombinant protein Cp51 was successfully expressed, and was subsequently purified with Ni2+-NTA affinity chromatography. The antibacterial activity of the recombinant protein Cp51 was detected by kinetic turbidimetric assay. 【Result】The bacteria turbidities of seven strains of C. perfringens type A were effectively reduced by the recombinant protein Cp51. The bactericidal rate was above 99.99% in 30 min after treatment of recombinant protein Cp51 at the concentration of above 5 μg·mL–1. The Cp51 protein had no bactericidal effect against other types of bacteria. 【Conclusion】The recombinant protein of bacteriophage lysine Cp51 has strong bactericidal activity and specificity in vitro against type A C. perfringens, which could provide a basis for clinical application of Cp51 lysin.
Clostridium perfringens; bacteriophage; lysin; protein expression; bactericidal activity
Q939.48;S945.1
A
1001-411X(2017)05-0019-05
吕梦娜, 龙航宇, 王丽梅, 等. A型产气荚膜梭菌噬菌体裂解酶Cp51的原核表达及活性检测[J]. 华南农业大学学报, 2017, 38(5): 19-23.
2016-11-29 优先出版时间:2017-07-14
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.s.20170714.0855.008.html
吕梦娜(1991—),女,硕士研究生,E-mail: 793453116@qq.com; 通信作者: 林瑞庆(1973—),男,副研究员,博士,E-mail: rqlin@scau.edu.cn
广东省科技计划项目(2014A020208099)
