干旱胁迫对西藏4份野生披碱草属牧草幼苗生理 指标的影响
2017-07-30闫天芳苗彦军王向涛魏臻武徐雅梅蔺永和
闫天芳, 苗彦军, 王向涛, 魏臻武, 徐雅梅*, 蔺永和
(1.西藏农牧学院,西藏 林芝 860000; 2.扬州大学动物科学与技术学院, 江苏 扬州225009; 3.兰州大学草地农业科技学院, 甘肃 兰州 730000)
西藏位于我国西部,地域辽阔,平均海拔4 000 m以上,年均气温4.8℃,降水表现出明显的地域性,从东南到西北依次递减(4 000~50 mm)[1]。高寒缺氧、雨水匮乏成为高原植物生长最大的限制因素,因此探索培育能耐寒耐旱的牧草品种对于西藏生态经济的可持续发展尤为重要。披碱草属牧草(Elymus)是一种多年生冷季型禾本科牧草,主要分布于我国青藏高原的西北、西南和华北地区,能够适应平均海拔在3 000~5 000 m的高原,常作为建群种,是青藏高原草原和草甸的重要组成部分[2]。由于其品质优良、产草量高、对于高原环境的适应性强,已经被广泛应用于高寒地区牧草地和人工草地的建植,是高寒地区生态恢复改良的主要牧草之一[3-4]。
当牧草遭受干旱胁迫时,细胞内活性氧发生代谢失调,加剧了细胞膜脂过氧化作用,从而氧化细胞膜系统发生严重的损伤。牧草长期适应性演化过程中为应对这种生理特性的损伤,从而逐渐形成了一套完善的抗氧化防御系统,来提高自身的抗旱性。研究认为:丙二醛、超氧化物歧化酶、游离脯氨酸、以及植物体内可溶性糖含量等生理指标可作为植物抗逆性生理评价的重要指标。研究发现,当植物受到外界环境胁迫时,植物细胞通过积累渗透调节物质保持细胞内膨压维持植物的生理过程,以丙二醛含量来反映植物细胞膜的受伤害程度,脯氨酸含量的增加可防止细胞脱水,超氧化物歧化酶活性对于防止自由基活性氧的毒害至关重要,可溶性糖的积累有利于提高细胞膨压[5-8]。目前较为科学的研究植物抗旱性的方法是在水分胁迫下测定植物的相关生理生化指标。只以单项指标来评价,结果存在片面性,需多个与抗旱性有关的指标应用数学方法对其进行综合分析,才能更加准确地评价出植物抗旱性。本试验选取来自不同生境的野生披碱草属植物作为试验材料,进行耐旱性鉴定,系统地分析在土壤水分胁迫下植株苗期叶片生理生化变化,利用模糊隶属函数法对其耐旱性进行综合评定。通过测定相关生理生化等指标,综合评定和分析不同生境来源披碱草属植物在水分胁迫下的生理适应机制及抗旱性评价,旨在为西藏地区披碱草属植物的抗旱性选育提供理论依据。
1 材料与方法
1.1 试验材料
供试材料:老芒麦采集于拉萨市,其它三种材料分别采集于西藏的林芝县、那曲县和申扎县(表1)。
表1 材料及来源Table 1 Materials and sources
注:下文图表中出现的材料名称均以编号表示。
Note: The codes of material names will be used in following Figures and Tables.
1.2 研究方法
本实验于2016年4月份在西藏农牧学院高寒草地实验室进行。选择成熟、籽粒饱满、大小均匀、健康无病虫害的种子为试验材料。用2% NaClO对待用牧草种子进行表面消毒5 min,再用蒸馏水冲洗3次。农场取土,在105℃的烘箱中烘5 h后装入在直径20 cm、高18 cm的塑料花盆中,在有孔塑料花盆底部铺双层滤纸,这样既可以通气,同时也阻止沙土漏出。每个花盆放入2.5 kg的烘干土壤,设置3个重复。在花盆内种植牧草种子进行浇水处理,使每盆土壤处于饱和含水状态即土壤的内部和表层均含水达到饱和状态,而表面的开口孔隙及面层却处于无水状态。置于人工气候箱内培养,温度为15/25℃,12 h光照,12 h黑暗处理。在2~3片真叶时进行定苗,待长到4~5叶龄时进行干旱胁迫处理(4月6日)即停止浇水。分别在干旱处理4 d、8 d、12 d、16 d,测定植株幼苗叶片生理生化指标,每个处理3次重复。取样时间为早晨9: 00-10: 00。
1.3 测量指标与方法
1.3.1植物叶片相对含水量(relative water conter,RWC) 采用饱和称重法[9]测定RWC。称取叶片5 g(Wf),在蒸馏水中浸泡24 h称取饱和鲜重(Wt)然后在110℃下烘干称样品干重(Wd)。用RWC(%)=(Wf-Wd)(Wt-Wd)×100% 计算,3次重复。
1.3.2脯氨酸(free proline,Pro)含量 采用酸性茚三酮法[10]测定Pro含量。称取叶片各0.2~0.5 g,加入3%磺基水杨酸5 mL,在沸水浴中提取10 min,冷却后,吸取2 mL提取液加入2 mL冰醋酸和3 mL酸性茚三酮,在沸水浴中提取40 min,冷却,加入5 mL甲苯,振荡,静置数分钟,待溶液分层后吸出甲苯层,用分光光度计在520 nm波长比色,3次重复。
1.3.3超氧化物歧化酶(superxide dismutase,SOD)活性 采用氮蓝四唑法[11]测定SOD活性。称取叶片0.5 g,加磷酸缓冲液研磨成浆,10000 r·min-1下离心10 min。取上清液0.1 mL加入0.05 mol·L-1磷酸缓冲液1.5 mL,130 mmol·L-1甲硫氨酸0.3 mL,750 μmol·L-1氮蓝四唑溶液0.3 mL,100 μmol·L-1EDTA-Na2溶液0.3 mL,加入20 μmol·L-1核黄素0.3 mL,再加入0.05 mL酶液,对照管加酸缓冲液代替酶液,最后加入蒸馏水0.25 mL,共3 mL反应液,混合均匀,将对照管置暗处,其他各管立即置于4 000 lx日光灯下反应20 min后,用黑布遮住,终止反应,以遮光的对照管作为空白,在波长560 nm下比色,3次重复。
1.3.4丙二醛(malondialdeehyde,MDA)含量 采用硫代巴比妥酸法[12]测定MDA含量。称取鲜叶片1 g,加入10%三氯乙酸(TCA)缓冲液研磨得10 mI二浆,4 000 r·min-1下离心10 min,取上清液2 mL,加入2 mL硫代巴比妥酸溶液,沸水浴加热15 min,冷却后4 000 r·min-1下离心10 min,以硫代巴比妥酸溶液为参比液,分别在波长532 nm、600 nm和450 nm处测定光密度值,3次重复。
1.3.5可溶性糖(soluble sugar content,SSC)含量 采用苯酚法[13]测定SSC含量。称取鲜叶片0.1~0.3 g,加蒸馏水,塑料膜封口,于沸水中提取30 min,过滤入25 mL容量瓶中,定容,作为待测液。取0.5 mL提取液,加1.5 mL蒸馏水,加1 mL的9%苯酚溶液,5 mL浓硫酸,摇匀,显色。在485 nm波长下比色,3次重复。
1.4 苗期抗旱性综合评价
1.4.1抗旱性评价方法 采用隶属函数法进行抗旱性评价[14]。
材料各指标隶属函数值:
μ(Xj)=(Xj-Xmin)/(Xmax-Xmin)
式中,Xj表示第j个指标值,Xmin表示第j个指标的最小值,Xmax表示第j个指标中的最大值。如果某一指标与抗旱性呈负相关,可采用反隶属函数法计算抗旱隶属函数值:
μ(Xj)=(Xmax-Xj)/(Xmax-Xmin)
求出各指标平均值后对不同材料间各指标间进行算数平均数进行比较,平均值越大代表耐旱性越强。
1.4.2抗旱性评价分级标准 将试验材料的综合抗旱能力D值划分为3级。Ⅰ 级:综合评价值0.7以上,为抗旱性强;Ⅱ 级:综合评价值在0.5~0.7之间,为抗旱性中等;Ⅲ 级:综合评价值在0.5以下,为抗旱性弱[14]。
1.5 数据分析
采用Excel 2013对原始数据进行处理,SPSS 19.0软件并用邓肯氏(Duncan)法进行单因素方差分析。
2 结果与分析
2.1 干旱胁迫对叶片RWC的影响
随着干旱胁迫时间的延长,幼苗叶片相对含水量持续降低,呈下降趋势(图1)。在干旱胁迫第4 d时,供试幼苗叶片相对含水量均保持在80~90%之间,幼苗色泽光鲜,含水量充足。干旱胁迫4~8 d时失水速度缓慢,土壤中水分较充足,失水速度缓慢。以胁迫8 d时相对含水量作为对照,LS的变化量最小仅为2%,SZ的变化量最大为12.8%。干旱胁迫8~12 d时,土壤出现板结现象,幼苗出现不同程度的萎蔫,失水速度达到最快。表现为叶片发生卷曲,部分叶尖与叶片边缘开始泛黄。出现干枯现象。干旱胁迫16 d时,花盆表面土壤板结情况严重,幼苗叶片出现不同程度的干枯现象,LZ叶片倒伏于花盆边缘。
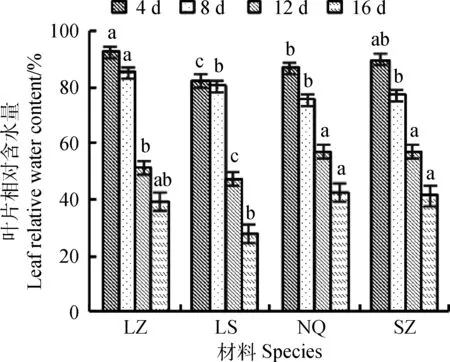
图1 干旱胁迫对叶片相对含水量影响Fig.1 Effect of drought stress on leaf relative water content注:图中不同小写字母表明差异显著(P<0.05 ),下图相同。Note:Different lowercase letters in theFigure indicate significant difference at the 0.05 level, the same as below.
2.2 干旱胁迫对MDA含量的影响
当供试幼苗遭受干旱胁迫时,随着胁迫时间的延长,MDA含量都表现出逐渐增加的趋势(图2)。胁迫8~12 d时 LZ、LS、NQ增幅达到最高,SZ在12 d后增幅达到最高。增长幅度达到了52.69%。以干旱胁迫第4 d作为对照,LZ和LS的MDA含量变化最小,分别而增加了14 nmol·g-1FW和14.24 nmol·g-1FW。SZ的MDA含量变化最大,变化量达到了23.67 nmol·g-1FW。供试幼苗MDA含量峰值与对照相比,变化差异较大,从小到大依次为:LZ 图2 干旱胁迫对叶片MDA含量的变化Fig.2 The content of MDA under different drought stress 供试幼苗叶片中SOD活性随干旱胁迫强度加剧呈先上升后下降的变化趋势(图3)。干旱胁迫4 d时各幼苗SOD活性差异性显著(P<0.05),其中LS的SOD活性最高,LZ的SOD活性最低。NQ与SZ的SOD活性最为接近,平均为276.61 U·g-1FW。胁迫8 d时SOD活性增长幅度从小到大排序依次为:LS < NQ < SZ 图3 干旱胁迫对叶片SOD含量影响Fig.3 The activity of SOD under different drought stress 随着干旱胁迫强度的加剧,Pro含量都呈明显上升的趋势(图4)。供试幼苗干旱胁迫第4 d时Pro含量接近,平均为24.66 μg·g-1FW。胁迫第8 d后Pro含量增长速度均达到最高,分别为106.8%,99.3%,115.7%,123.6%,其中LS增长速度最小为99.3%,SZ增长速度达到了123.6%。胁迫12~16 d时,Pro含量增长速度均明显变慢,胁迫16 d时,Pro含量达到峰值且差异显著(P<0.05)。其中Pro含量最高的是SZ,高达163.8 μg·g-1FW,最低的是LS,为101.03 μg·g-1FW。4份材料Pro含量由少到多依次为LS 图4 干旱胁迫对叶片Pro含量影响Fig.4 The content of pro under different drought stress 供试幼苗随着干旱胁迫时间的延长,SSC含量均表现出逐渐增加的趋势(图5)。在干旱胁迫12 d时,供试幼苗SSC含量差异显著(P<0.05)。LZ与SZ在胁迫16 d时涨幅达到最高分别为69.69%和91.96%。NQ在胁迫8 d时涨幅达到最高为61.05%,LS在胁迫4~8 d时SSC含量变化量最小仅为9.41%,供试幼苗都在胁迫16 d时可溶性糖含量达到峰值,分别为7.31 mg·g-1,4.83 mg·g-1,5.83 mg·g-1,9.26 mg·g-1,其中SSC含量SZ最高,LS最低。在胁迫期间供试幼苗SSC含量增幅达到最高点的时间各不相同,LZ增幅在12 d后达到最高,LS在胁迫4~8 d时SSC含量变化差异不显著,NQ在4 d后增幅达到最大随后增长速度趋于平缓,SZ在胁迫4~12 d时增长速度均匀,12~16 d时增长速度突然增大。方差分析表明,各供试幼苗在干旱胁迫条件下叶片中SSC含量差异性显著(P<0.05)。 图5 干旱胁迫对叶片SSC含量影响Fig.5 The content of soluble sugar under different drought stress 植物的耐旱性是由多种因素相互作用而构成的一个较为复杂的综合性状。本试验将供试的4份材料与抗旱性有关的5个指标进行抗旱指数综合分析,计算出供试材料与各指标的隶属度值,以供试材料的抗旱算数平均隶属度作为抗旱鉴定综合评价指标。综合评价值越大,抗旱性越强,反之则弱。表2为4种材料的综合评价结果。LZ、LS、NQ、SZ抗旱隶属度分别为0.552,0.544,0.560和0.573,都介于0.4~0.7之间,属于中等抗旱性。供试材料抗旱能力强弱依次为SZ > NQ > LZ >LS。 表2 不同材料抗旱指标的抗旱指数隶属及其隶属度评价Table 2 Analysis of subordinate function values of drought resistance index of different materials 叶片RWC反映植物组织的含水状况是以植物组织饱和含水量为基础来表示,被用来表示植株遭受水分胁迫后水分亏缺程度的参数[15]。本试验研究表明在干旱胁迫下,植物叶片中RWC与抗旱性呈正相关[16-19]。在胁迫第8 d后叶片供试材料相对含水量下降速度变快,其中以LZ、LS下降最快,NQ和SZ下降速度相对较慢,由此表明相对含水量下降速度越慢抗旱性越强。植物叶片RWC下降越严重,叶片萎蔫越严重,说明膜的透性变大,植物细胞受伤越严重,抗旱性随之变差。这与祁娟[17]等在披碱草与老芒麦苗期抗旱性综合评价的研究结果一致。 植物在逆境条件下或器官发生衰老时,会发生膜质过氧化作用,其中MDA是主要产物之一,其含量常被用作衡量细胞膜脂过氧化指标。MDA含量越高,植物组织受伤越重,植物抗旱能力越弱[20]。本试验研究表明:供试材料随干旱胁迫强度加剧,MDA含量呈上升趋势。供试材料在干旱胁迫下,MDA含量均存在显著差异(P<0.05),表明在干旱胁迫下植物幼苗受到了不同程度的损害。在干旱胁迫4~8 d时,供试材料叶片MDA含量与对照(以胁迫第4 d时MDA含量作为对照)相比变化不大。这表明植物在短期逆境条件下,植物器官遭受伤害程度小。在胁迫8 d后MDA含量大幅度上升,表明植物体内细胞受到严重破坏,增幅越大,植物细胞受伤害程度越大,抗逆性越差。SZ在胁迫12 d后MDA含量增幅达到最大,表明其抗旱性最弱,植物叶片MDA缓慢增加的时间维持越长,植物的抗旱性越强,反之则越弱,这与鲁存海[20],程红焱[21]研究结果一致。在干旱胁迫下,4份供试材料MDA含量均显著增加,不同材料间MDA含量增长速度不同。其中,SZ的MDA含量增幅达到最高的时间最晚,说明植物膜系统受到伤害时对于逆境的胁迫持续的时间最久,抗旱性最强,NQ次之,LZ与LW最弱。 SOD是植物体内最重要的抗氧化酶,SOD活性大小与植物抗逆性相关[22]。在逆境胁迫下,SOD活性对于防止细胞膜遭受自由基活性氧的毒害至关重要。在此期间其活性往往会升高,来保护植物细胞膜免受自由基活性氧毒害,在逆境条件下,清除自由基的能力越强,保护酶活力维持的时间越久,植物的抗逆性就越强。试验结果显示,在干旱胁迫下,4份供试材料SOD活性均呈现先增加后降低趋势。均在胁迫8~12 d时SOD活性涨幅最高,在胁迫12d时各供试幼苗中SOD活性达到最大且达到峰值,随后SOD活性开始迅速下降。其中LZ、LS下降幅度最快分别达到了62.4%,72.3%。NQ和SZ相对较慢为45.2%,52.5%。结果表明干旱胁迫可使SOD活性增强,在同等胁迫条件下,SOD达到峰值迅速下降时,下降速率越快,抗旱性越弱。胁迫后期SOD活性出现迅速下降的现象可能是由于随着胁迫强度的加剧,植物细胞多种生理功能受到破坏,发生生理代谢紊乱,使得SOD活性受到抑制。也可能是由于胁迫强度过大,活性氧和防御系统的动态平衡遭到了破坏,使酶活性降低[23]。原因有待进一步考证。 Pro既可作为渗透调节物质又能与蛋白质相互作用。植物在正常条件下, Pro含量很低。但遭受逆境胁迫时,便会大量积累来保持原生质与环境的渗透平衡。与蛋白质相互作用增加蛋白质可溶性或减少可溶性蛋白的沉淀,可作为植物抗逆性生化指标[24]。在干旱胁迫下,4份供试材料Pro含量随着胁迫程度的加剧,供试材料Pro含量都持续升高。目前,对于Pro含量的增加与植物耐旱性间的关系还存在很多异议,但多数研究者认为Pro能降低细胞的渗透势、具有保护和稳定细胞生物大分子的作用,其含量的高低与植物的抗旱性呈正相关[25-26]。本试验中,4份供试材料Pro含量随胁迫程度的加强在增大,在胁迫第8 d后供试材料Pro含量增幅达到最大。其中SZ在遭受胁迫时Pro含量明显高于其他材料,其抗旱性最高。当植物受到逆境胁迫时Pro含量越高,其抗旱性越高,本试验结果与张昌胜、郭郁频[27]等的研究结论相一致,说明逆境下Pro含量升高对植物体逆境胁迫抗性有利。 植物在干旱胁迫环境中,SSC也是植物体内的渗透调节物质,参与渗透调节,有利于植物在干旱胁迫下吸收水分,从而提高植物的抗旱性[28]。也有研究者认为SSC在维持蛋白质稳定方面可能也起到重要作用,保证细胞的正常生理功能[29]。本试验中,在干旱胁迫下,4份供试材料SSC含量随着胁迫强度的加剧呈上升趋势。其中SZ增加幅度最大,且SSC含量最高,说明SSC含量对于增强其抗旱性起到了一定作用。LZ、LS、SZ都在胁迫12 d后SSC含量出现大幅度增长,而NQ在胁迫4 d后就发生SSC含量大幅度增长现象,在胁迫8~16 d时SSC含量增长速度呈匀速增长。表明NQ材料在受到干旱胁迫时,其叶片内SSC含量对于干旱胁迫的敏感程度高于其他材料。当胁迫强度加剧时,供试材料SSC含量变化差异显著,对于抗旱性的影响较大,可作为抗旱性鉴定指标。结果表明当植物遭受干旱胁迫时,SSC含量的增高对于植物对抗干旱胁迫起到积极作用。与王川[30],张小娇[31]等研究结论一致。 植物所在生境年平均降水量对于植物的抗旱性具有一定影响[32-33]。本实验研究发现海拔高度对于披碱草属植物抗旱性呈正相关,年平均降水量对于披碱草属植物抗旱性呈负相关。由表1可看出在西藏地区海拔高度的高低又直接影响着当地的年平均降水量,随着海拔高度升高,年平均降水量降低。在降水量偏低的地区披碱草需适应本地环境而变的更具耐旱性,海拔高度与披碱草的抗旱性之间的关系,可能是由于海拔高度升高间接影响植物生境的降水量,继而影响植物的抗旱性,植物的抗旱性与海拔高度有何影响,需进行进一步研究。本实验研究表明随着海拔高度的升高披碱草属植物的抗旱性随之变强,所在生境的年平均降水量越低披碱草属植物的抗旱性越强。本实验研究材料所在生境的海拔高度排序依次为:SZ>NQ>LS>LZ;通过抗旱性评价得出抗旱性强弱顺序由强到弱依次为:SZ>NQ>LZ>LS。在降水量相对较高的林芝垂穗披碱草抗旱性高于在降水量相对较低的拉萨老芒麦,可推断出同属披碱草属的垂穗披碱草抗旱性高于老芒麦。 4种披碱草属植物幼苗在干旱胁迫条件下,RWC含量随胁迫程度的加剧而下降,MDA、Pro及可溶性糖含量随胁迫程度的加剧而上升,SOD则是先上升后下降。5个生理指标在4种披碱草属植物幼苗间均存在显著差异,可以作为披碱草属植物抗旱性的评价指标。在胁迫8 d后各测试指标的变化趋势均出现了明显的转折点,说明8 d后土壤的含水量可能是披碱草属耐旱的关键水势点。本研究采用模糊隶属函数法对西藏4种野生披碱草属植物抗旱性进行综合评价后,得出其抗旱性强弱顺序依次为:申扎垂穗披碱草>那曲垂穗披碱草>林芝垂穗披碱草>拉萨老芒麦。综合得出在申扎地区的野生垂穗披碱草在高寒缺氧、雨水匮乏的青藏高原繁育前景广阔,为选育具有抗旱、抗寒的优良垂穗披碱草提供了依据。
2.3 干旱胁迫对SOD活性的影响

2.4 干旱胁迫对Pro含量的影响

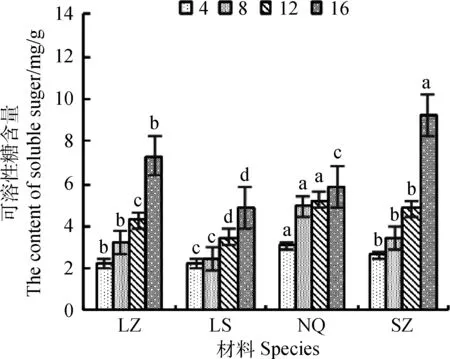
2.5 干旱胁迫对SSC含量影响
2.6 抗旱性综合评价

3 讨论
4 结论
