河北省森林草原区草本植物物种多样性和功能多样性
2017-07-21葛兆轩孙国龙袁业黄选瑞张志东
葛兆轩,孙国龙,袁业,黄选瑞,张志东*
(1.河北省林木种质资源与森林保护重点实验室,河北农业大学林学院,河北 保定071000; 2.河北省塞罕坝机械林场, 河北 围场068466)
河北省森林草原区草本植物物种多样性和功能多样性
葛兆轩1,孙国龙2,袁业1,黄选瑞2,张志东1*
(1.河北省林木种质资源与森林保护重点实验室,河北农业大学林学院,河北 保定071000; 2.河北省塞罕坝机械林场, 河北 围场068466)
理解物种多样性和功能多样性之间的关系以及分析它们对环境变化的响应,有利于揭示生物多样性的影响因素。本研究以塞罕坝地区不同植被类型(草地、灌木林和天然次生林)的草本群落为研究对象,分析了物种多样性(物种丰富度、Shannon-Wiener多样性和Pielou均匀度)和功能多样性测度指标(功能丰富度、功能均匀度和功能离散度)之间的相互关系,并采用相似性分析检验(ANOSIM)和冗余度(RDA)分析等方法研究了它们随植被类型和环境因子的变化规律。结果表明,草地中草本植物物种丰富度、Shannon-Wiener多样性指数显著高于灌木林和天然次生林中的草本群落(P<0.05),而功能丰富度指数在草地中最低(P<0.05),功能均匀度和功能离散度在灌木林中最低(P<0.05),Pielou均匀度指数在不同植被类型间差异不显著(P>0.05);功能丰富度与物种丰富度、Shannon-Wiener多样性指数显著负相关(P<0.01),功能离散度与物种均匀度显著正相关(P<0.01),而功能均匀度与所有物种多样性指数均不相关(P>0.05);影响物种多样性的主要因子有土壤含水量、土壤有机质、土壤pH值、土壤全氮含量和坡向,影响功能多样性的主要因子有土壤有机质、土层深度、土壤含水量和海拔。物种多样性和功能多样性分布主要受土壤因子的影响,分别占到总解释变异的29.1%和29.8%。本研究结果表明研究区草本植物物种多样性和功能多样性变化相对独立,物种多样性可能主要是由资源可利用程度决定,而功能多样性可能是生态位分化即环境筛选和物种间相互作用共同决定。
物种多样性;功能多样性;环境因子;植被类型;因子分解
物种多样性是生态系统功能的基础,它可以体现群落的结构、群落组织水平、发展阶段、稳定性程度和生境差异[1]。功能多样性是生物多样性的一个非常重要的组成部分,是指影响群落或生态系统中有机体性状的数值和范围[2]。阐明物种多样性和功能多样性的关系是揭示生物多样性对生态系统功能影响的关键[3]。环境因子作为一个“筛”,能够决定哪些物种或者性状可以在群落中生存和维持[4]。目前,不同植被类型物种多样性和功能多样性与环境因子间关系的研究仍然较少[5-7]。
以往有关生物多样性对生态系统功能影响的研究主要以物种多样性来进行,而忽视了生物多样性的不同组分在生态系统中的功能效应[8]。近年来,功能多样性的研究逐渐受到重视[9-11]。一般来讲,群落尺度物种多样性和功能多样性之间的关系表现为正线性相关[12]。受扰动强度和环境胁迫的影响,二者之间的关系也会表现为负相关[13]、S型曲线关系[14]和不相关[15]。很多环境因素如海拔、坡向、土壤含水量、土壤养分等会影响物种或功能多样性的分布。在亚高山草甸,北坡的物种多样性显著高于南坡,而功能多样性却呈相反的变化[16];随着土壤含水量的增加,物种多样性亦表现为增加趋势,但功能多样性变化不大[13]。由此可见,不同的生态系统物种多样性和功能多样性对不同环境梯度的响应有可能不一致。在特定的区域,阐明物种多样性和功能多样性随环境梯度的变化规律有助于更好地揭示物种共存机制和群落构建规则[17]。
塞罕坝地区是华北典型的森林-草原生态交错带,生态系统复杂多样,生物多样性丰富,具有很好的保护价值。但由于长期人为干扰以及土地沙漠化、荒漠化的影响,生物多样性呈下降趋势。本研究以塞罕坝地区不同植被类型的草本群落为研究对象,通过样地调查,主要探究以下3个问题:1) 物种多样性与功能多样性在不同植被类型间的变化是否相似?2) 物种多样性和功能多样性测度指标之间关系如何?3) 影响物种多样性和功能多样性分布的主要环境因子是否相同?
1 材料与方法
1.1 研究区概况
研究区位于河北省塞罕坝机械林场,地处内蒙古高原与冀北山地交接处(42°22′ N,116°53′ E)。地形地貌组合为高原-波状丘陵-漫滩-接坝山地[18]。气候属寒温带半干旱-半湿润季风气候。年均气温-1.40 ℃,极端最高气温32 ℃,极端最低气温-42.8 ℃;年平均降水量445 mm,蒸发量1358 mm[19]。植被由森林向草原过渡,天然次生林主要树种有华北落叶松(Larixprincipis-rupprechtii)、山杨(Populusdavidiana)、蒙古栎(Quercusmongolica)、白桦(Betulaplatyphylla);灌木种主要有山刺玫(Rosadavurica)、山杏(Armeniacasibirica)、柔毛绣线菊(Spiraeapubescens)等;草本主要由地榆(Sanguisorbaofficinalis)、金莲花(Trolliuschinensis)、披针叶苔草(Carexlanceolata)和蚊子草(Filipendulapalmata)等物种构成。
1.2 样地设置和群落调查
2014年7—9月在研究区按照分层随机取样的方法进行群落学调查,选取了32个斑块(斑块面积介于2.18~74.06 hm2)的调查数据进行物种多样性和功能多样性分析,其中包括10个草本,10个灌木林和12个天然次生林斑块。按照斑块大小设置不同数量的草本样方,样方间隔≥50 m,样方大小为1 m×1 m,共153个样方。记录每个样方的海拔、坡度、坡向、坐标以及植被类型;记录草本植物物种名、盖度、平均高度和最大高度。在每个草本样方内,选取盖度大于90%的所有物种,每个物种采集长势良好的10株个体(样方内不足10株则于样方附近采样),每株取不少于5片完好成熟的叶片,测定叶片鲜重、叶面积、叶绿素含量,所有样品置于105 ℃烘至恒重,以1/10000电子天平称重,计算比叶面积、叶干物质含量,测定方法详见功能性状测量手册[20]。在采集叶片样品的同时,从每个植株茎基部位截取长约10 cm的枝条,每个物种取5~10根,放入塑封袋中置于保温箱中保存。用水替代法测定枝条体积,然后置于105 ℃烘箱内烘至恒重,称量其干重,枝条干重与体积之比即为茎密度。将植物样品用研磨仪粉碎,采用凯氏定氮法测定叶片氮含量,钼锑抗比色法测定叶片磷含量,重铬酸钾法测定叶片有机碳含量[21]。
在每个样方中心位置采集0~10 cm、10~20 cm、20~30 cm三层土壤样品,每层取一定量土样混合后放到铝盒中保存。将取得的土样带回实验室自然阴干后,参照《土壤农化分析手册》[22]测定土壤含水量、土壤pH值、有机质含量、全磷含量、全氮含量和土壤速效钾含量。
1.3 生物多样性计算
物种重要值体现了物种在群落中的优势程度,草本物种的重要值用相对频度、相对盖度和相对高度的加和平均来表示。
物种多样性具有多种测度指标,本研究选取了3个常见的物种多样性指数,包括:物种丰富度、Shannon-Wiener指数和Pielou均匀度指数(表1)。

表1 生物多样性计算公式Table 1 The formula of biological diversity indices
选取了118个草本物种的功能性状(比叶面积、叶干物质含量、叶绿素含量、叶氮含量、叶磷含量、叶碳含量和茎密度)实测数据,进行功能多样性的计算。功能丰富度指群落中物种占有的n维功能性状凹凸包量;功能均匀度指群落中功能性状值在凹凸包量中排列的均匀性;功能离散度指群落中每个物种的n维功能性状值到所有功能性状空间重心的平均距离[23-25]。计算公式见表1。
环境数据共选取了13个因子,其中包括2个空间因子、4个地形因子和7个土壤因子(表2)。
1.4 数据分析
物种累积曲线(species accumulation curve)用于比较不同植被类型物种累积速率的变化。采用ANOSIM(analysis of similarities)方法检验物种组成在3个不同植被类型之间是否有显著差异。对物种多样性和功能多样性指数分别进行单因素方差分析(one-way ANOVA),并进行了不同植被类型物种多样性和功能多样性的多重比较(Tukey HSD)。应用皮尔逊相关方法分析物种多样性和功能多样性之间的关系。
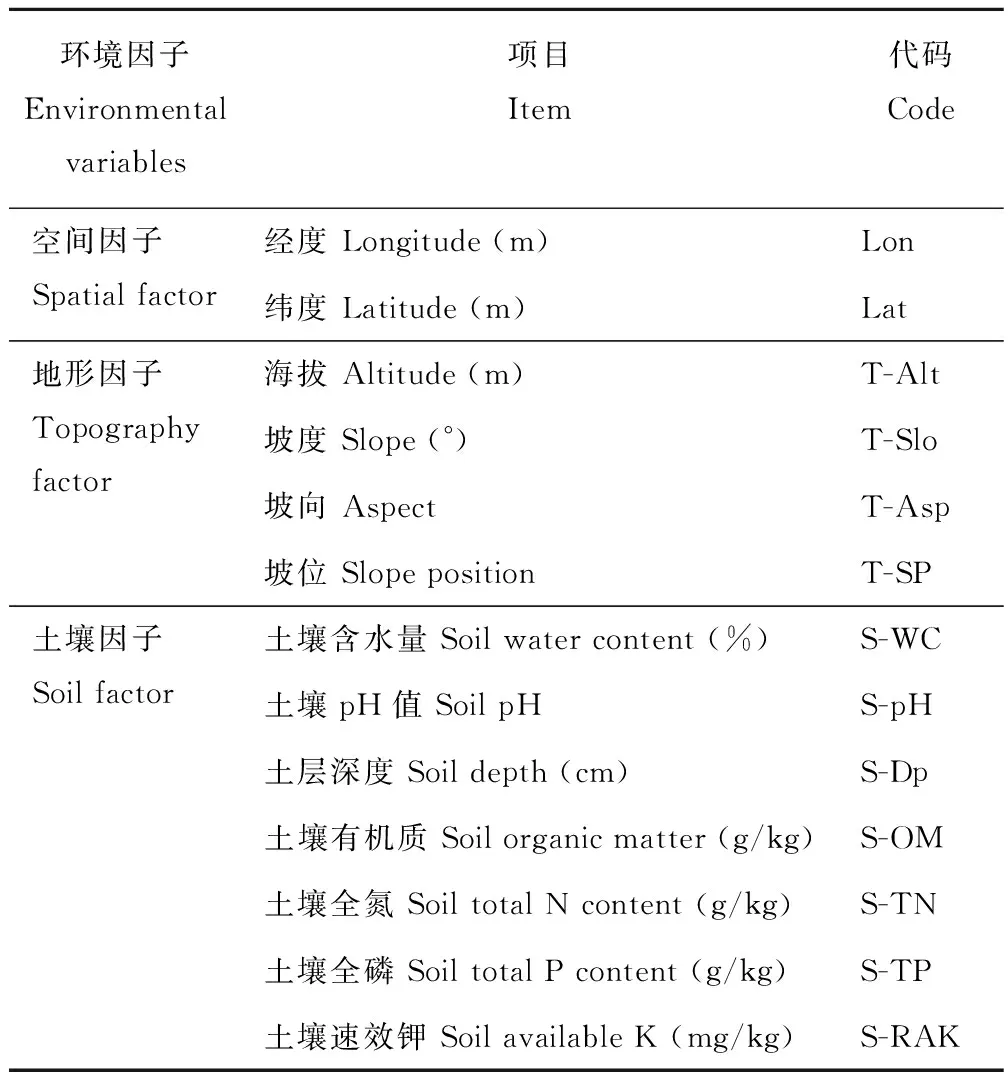
表 2 环境因子的选择与定义Table 2 Selection and definition of environmental variables
运用冗余度分析(redundancy analysis,RDA)探讨环境因子对不同植被类型物种多样性和功能多样性的影响。首先建立物种多样性-环境变量、功能多样性-环境变量矩阵,用去趋势对应分析(detrended correspondence analysis,DCA)估计排序轴梯度长度分别为0.1和0.7,表明两个数据文件均具有较好的线性反应,利用线性模型比较适宜。为了检验环境因子对物种和功能多样性的影响,首先利用Monte Carlo检验(置换次数为499)判断其重要性是否显著,当候选变量P≥0.05时,予以排除。为了更好地反映环境因子对多样性影响的解释效果,将解释变异分解为以下几部分:1)纯空间因子解释部分(Rsp);2)纯地形因子解释部分(Rto);3)纯土壤因子解释部分(Rso);4)空间因子和地形因子交互解释部分(Rspto);5)空间因子和土壤因子交互解释部分(Rspso);6)土壤因子和地形因子交互解释部分(Rsoto);7)3个因子交互解释部分(Rsptoso)。
利用R 3.3.1程序FD软件包中的dbFD函数计算物种多样性及功能多样性指数,统计分析在R 3.3.1程序Vegan软件包中完成(R development Core Team, 2011)。
2 结果与分析
2.1 物种组成

图1 基于样本的物种累积曲线Fig.1 Sample-based species accumulation curve
在32个斑块的3个植被类型中共记录了294个草本物种,分属46科,168属。其中草地物种数最高,共记录了187个物种,分属38科、118属,分别占了总物种、科和属数的63.6%、82.6%和70.2%;其次是灌木林,共记录了156个物种,分属34科、106属,分别占了总物种、科和属数的53.1%、73.9%和63.1%;天然次生林物种数最低,共记录了121个物种,分属27科、81属,分别占了总物种、科和属数的40.8%、58.6%和48.2%。
3个植被类型中草本物种数随样地数累积的结果如图1,草地植被类型具有较快的物种累积速率,天然次生林物种累积速率较慢。并且ANOSIM检验结果显示,3个植被类型物种组成差异显著(r=0.4167,P=0.001)。
2.2 不同植被类型草本植物物种多样性和功能多样性变化
不同植被类型间物种丰富度、Shannon-Wiener多样性指数(H)差异显著(P<0.05),草地物种丰富度、Shannon-Wiener多样性指数(H)显著高于灌木林和次生林植被类型。草地和次生林植被类型间功能丰富度(FRic)差异显著(P<0.05),草地功能丰富度(FRic)显著低于次生林和灌木林植被类型,次生林和灌木林植被类型间功能均匀度(FEve)和功能离散度(FDis)差异显著(P<0.05),并且灌木林低于次生林和草地植被类型(图2)。

图2 不同植被类型物种多样性和功能多样性差异性分析Fig.2 Difference analysis of species diversity and functional diversity among different vegetation types 不同字母表示显著差异(P<0.05) The different letters indicate significant differences at P<0.05 level; GL:草地 Grassland; SF: 灌木林 Shrub forest; NS: 天然次生林 Natural secondary forest.下同 The same below.
2.3 物种多样性与功能多样性的关系
FRic与S、H极显著负相关(P<0.01),而与J关系不大(P>0.05);FDis与J呈极显著正相关(P<0.01),而与S和H不相关(P>0.05);FEve与所有物种多样性指数均不相关(P>0.05)(表3)。

表3 物种多样性与功能多样性相关性Table 3 The relationship between species diversity and functional diversity
“**”,P<0.01 ;“-”,负相关 Negative correlation.
2.4 不同植被类型环境因子对比分析
通过对不同植被类型的环境因子进行单因素方差分析,土壤全磷含量、全氮含量和土壤含水量差异显著(P<0.05),只对这3个因子进行了多重比较(图3)。草地土壤全氮含量、土壤含水量显著高于灌木林和天然次生林,草地中全磷含量显著高于灌木林,而与天然次生林差异不显著(P>0.05)。

图3 不同植被类型环境因子对比分析Fig.3 Variations of environmental variables among different vegetation types
2.5 物种多样性和功能多样性与环境因子相关性
土壤含水量、土壤pH值、坡向和土壤全氮含量与物种多样性各指数正相关,而随着土层深度和速效钾含量的增加,物种丰富度、Shannon-Wiener多样性降低(图4A);对于功能多样性来说,FRic主要受到土壤有机质和土层深度的影响而随土壤含水量的增加而减小,FEve与土层深度正相关性,FDis主要受海拔、土壤含水量和土层深度的影响,表现为正相关(图4B)。

图4 物种多样性(A)和功能多样性(B)与环境因子RDA排序图Fig.4 Redundancy analysis ordination diagram of the species diversity (A), functional diversity (B) and environmental factors
2.6 因子分解
对影响物种多样性的环境因子进行定量分解可知,空间因子、地形因子和土壤因子共同解释效果达到了53.3%,其中纯土壤因子的解释效果最显著,占到了总解释变异部分的29.1%,而纯空间因子和纯地形因子的解释效果相对较弱,分别只占到9.3%和10.1%,各环境因子之间相互耦合作用相对较小(图5A)。
对于功能多样性(图5B),3类环境因子共同解释效果达到60.5%,纯空间因子所起到的解释效果相对较弱,占总变异解释部分的6.5%,纯地形因子所起到的解释效果占16.3%,土壤因子的解释能力为29.8%。空间和地形因子的耦合作用相对较大,占总变异解释部分的5.0%,3类环境因子共同作用最小,占总变异解释部分的0.2%。
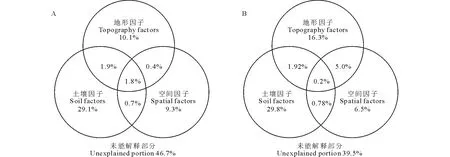
图5 影响物种多样性(A)和功能多样性(B)的环境因子的分解变异Fig.5 Variance partitioning of environmental factors for explaining species diversity (A) and functional diversity (B)
3 讨论
3.1 物种多样性变化
一般认为,物种多样性是由资源可利用程度决定的[26],也有的学者将这一观点称之为生产力假说或能量假说[27],即高的环境能量形成高的生产力,进而承载高的物种多样性。如随着土壤含水量[16]、土壤氮含量[28]的增加,物种多样性呈增加趋势,而随生境干燥程度的增加而降低[13]。本研究中,草地具有较高的土壤含水量、土壤全氮含量和土壤全磷含量(图3)。相对于灌木林和天然次生林,草地中可以说具有良好的资源供给能力,因而导致了更多的物种共存。这可从草地中草本物种丰富度和Shannon-Wiener多样性指数较高,灌木林次之,而天然次生林最低(图2)的结果中得到很好的证明。通过冗余度分析表明物种多样性各指数与土壤含水量、土壤全氮含量表现出较强正相关性(图4A),表明研究区天然次生林和灌木林中相对较低的土壤含水量和全氮含量是影响草本植物生长和分布的限制性因素,这在其他学者的研究中[28-29]也得到了相似的结论。在塞罕坝地区,内陆河岔较多,多数水源补充入就近的沼泽或滩地。据塞罕坝机械林场资料记载,该区有沼泽地和滩地面积约1326.7 hm2。在本研究调查的10个草地斑块中,大多数落在了湿地中,导致了草地中具有较高的土壤水分和养分含量,草本物种多样性较高。
通过对所选环境因子进行定量分解可知,影响物种多样性分布的未能解释部分占到了46.7% (图5A),表明该地区草本物种多样性除了受土壤水分、养分等的影响外,还可能受种间竞争(乔草、灌草间)、人为干扰(商业采伐、放牧等)、光环境异质性等的影响[30],更多的机理性研究仍有待进一步探讨。
3.2 功能多样性变化
在一个群落中,物种功能性状的值、范围和分布程度决定了功能多样性的大小[2]。环境筛选会使适应特定生境的物种共存,物种的功能性状出现趋同现象,这在一定程度上会降低功能多样性[8,14]。但是,在环境胁迫的条件下,性状生态位空间也可能会导致强烈的分化,这样具有不同功能性状的物种可以高效互补的利用有限的生态资源[13]。与物种丰富度相反,本研究中,功能丰富度在草地中最低,而在天然次生林和灌木林中相对较高(图2)。这一结果很可能是因为,在塞罕坝机械林场,由于过去长期的以追求木材效益最大化的经营目标,大面积的立地条件较好的天然次生林和灌木林皆被伐用于种植用材林,现存的天然次生林多处于坡度较陡、土壤瘠薄的生境中。由于土壤水分、养分等资源的限制,导致物种生态位分化程度要显著高于生境条件较好的草地中,进而引起了性状趋异,功能多样性增加。Bello等[13]在研究功能多样性在干旱和湿润地区的差异上,也发现了类似的规律。在本研究中,除了土壤因子、地形因子和空间因子外,其他未能解释的部分仍然高达39.5%(图5B)。有研究表明植物群落内光环境异质性的变化、种间和种内作用、外部干扰等均会影响植物功能性状的变化和个体生态策略的改变,进而影响功能多样性[13,31]。
3.3 物种多样性和功能多样性
本研究表明功能多样性的测度指标在不同植被类型间的变化趋势与物种多样性的变化模式并不相同(图1),不同测度指标之间存在正相关、负相关和不相关的情况(表3),证实了物种间的功能差异并不能由物种多样性简单代替,与其并不成比例关系[32]。相似的结论也在其他的研究中得到了验证。如在森林和采伐迹地生境,由于生态构建过程的不同,物种多样性和功能多样性间的关系存在显著差异[33]。也有研究指出物种多样性与功能多样性呈正相关[8,12],随着干扰和环境胁迫的增加,物种多样性与功能多样性也可由正相关变为不相关[34]或负相关[13,16,35]。由前文分析可知,物种多样性主要是由资源可利用程度决定,而功能多样性是由环境筛选和物种间相互作用即生态位分化所决定,不同的生态构建过程影响了不同生境中物种多样性和功能多样性的关系差异[33]。本研究物种多样性相关指标沿草地、灌木林和次生林植被类型显著降低,而功能多样性测度指标在草地中最低,而天然次生林和灌木林相对较高的结论,正是以上生态学假说综合作用的结果。
4 结论
本研究揭示了草地、灌木林和天然次生林下草本群落物种多样性和功能多样性测度指标随环境梯度(如土壤含水量、土壤有机质等)的响应差异。物种丰富度、Shannon-Wiener多样性指数沿草地、灌木林和次生林植被类型呈显著降低趋势,而功能丰富度则在草地植被类型中最低,天然次生林和灌木林相对较高。相关分析表明,功能多样性和物种多样性测度指标之间存在正相关、负相关和不相关的情况。以上结果反映了不同植被类型草本群落构建机制的不同。在研究区,土壤养分差异是影响物种多样性的主要原因,土壤水分、养分胁迫导致生态位分化,决定功能多样性。本研究结果进一步证实了物种多样性主要是由资源可利用程度决定,而功能多样性是由环境筛选和物种间相互作用即生态位分化共同决定,该研究结论可为塞罕坝地区生物多样性保护和管理提供科学依据。
References:
[1] Ma K P, Huang J H, Yu S L,etal. Plant community diversity in Dongling Mountain, Beijing, China; Ⅱ. Species richness, evenness and species diversities. Acta Ecologica Sinica, 1995, 15(3): 268-277. 马克平, 黄建辉, 于顺利, 等. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277.
[2] Petchey O L, Gaston K J. Functional diversity: back to basics and looking forward. Ecology Letters, 2006, 9(6): 741-758.
[3] Zhang Q G, Zhang D Y. Biodiversity and ecosystem functioning: recent advances and trends. Biodiversity Science, 2003, 11(5): 351-363. 张全国, 张大勇. 生物多样性与生态系统功能:最新的进展与动向. 生物多样性, 2003, 11(5): 351-363.
[4] Bu W S. The Relationships Between Biodiversity and Ecosystem Functioning in Natural Tropical Forests of Hainan Island, China[D]. Beijing: Chinese Academy of Forestry, 2013. 卜文圣. 海南岛热带天然林生物多样性与生态系统功能关系的研究[D]. 北京: 中国林业科学研究院, 2013.
[5] Suding K N, Goldstein L J. Testing the Holy Grail framework: using functional traits to predict ecosystem change. New Phytologist, 2008, 180(3): 559-562.
[6] Ledo A, Burslem D F R P, Condés S,etal. Micro-scale habitat associations of woody plants in a neotropical cloud forest. Journal of Vegetation Science, 2013, 24(6): 1086-1097.
[7] Jiang Y, Zang R G, Letcher S G,etal. Associations between plant composition/diversity and the abiotic environment across six vegetation types in a biodiversity hotspot of Hainan Island, China. Plant and Soil, 2016, 403(1): 21-35.
[8] Díaz S, Cabido M. Vive la différence: plant functional diversity matters to ecosystem processes. Trends in Ecology & Evolution, 2001, 16(11): 646-655.
[9] Stevens R D, Cox S B, Strauss R E,etal. Patterns of functional diversity across an extensive environmental gradient: vertebrate consumers, hidden treatments and latitudinal trends. Ecology Letters, 2003, 6(12): 1099-1108.
[10] Sun G J, Zhang R, Zhou L. Trends and advances in researches on plant functional diversity and functional groups. Acta Ecologica Sinica, 2003, 23(7): 1430-1435. 孙国钧, 张荣, 周立. 植物功能多样性与功能群研究进展. 生态学报, 2003, 23(7): 1430-1435.
[11] Jiang X L, Zhang W G. Functional diversity and its research method. Acta Ecologica Sinica, 2010, 30(10): 2766-2773. 江小雷, 张卫国. 功能多样性及其研究方法. 生态学报, 2010, 30(10): 2766-2773.
[12] Petchey O L, Gaston K J. Functional diversity (FD), species richness and community composition. Ecology Letters, 2002, 5(3): 402-411.
[13] Bello F D, Jan L, Sebastià M T. Variations in species and functional plant diversity along climatic and grazing gradients. Ecography, 2006, 29(6): 801-810.
[14] Sasaki T, Okubo S, Okayasu T,etal. Two-phase functional redundancy in plant communities along a grazing gradient in Mongolian rangelands. Ecology, 2009, 90(9): 2598-2608.
[15] Li X G, Zhu Z H, Zhou X S,etal. Effects of clipping, fertilizing and watering on the relationship between species diversity, functional diversity and primary productivity in alpine meadow of China. Chinese Journal of Plant Ecology, 2011, 35(11): 1136-1147. 李晓刚, 朱志红, 周晓松, 等. 刈割、施肥和浇水对高寒草甸物种多样性、功能多样性与初级生产力关系的影响. 植物生态学报, 2011, 35(11): 1136-1147.
[16] Li X E. Patterns of species diversity and functional diversity along a south- to north-facing slope in a subalpine meadow. Community Ecology, 2011, 12(2): 179-187.
[17] Mason N W H, Bello F D, Doležal J,etal. Niche overlap reveals the effects of competition, disturbance and contrasting assembly processes in experimental grassland communities. Journal of Ecology, 2011, 99(3): 788-796.
[18] Qin P Y, Yang H J, Jiang F L,etal. Quantitative classification of natural plant communities in the Saihanba National Nature Reserve, Hebei, Province, China. Chinese Journal of Applie Ecology, 2016, 27(5): 1383-1392. 秦朋遥, 杨会娟, 蒋凤玲, 等. 河北省塞罕坝保护区天然植物群落数量分类. 应用生态学报, 2016, 27(5): 1383-1392.
[19] Zhang S Z, Li M, Zhang S B,etal. Factors affecting natural regeneration ofLarixprincipis-rupprechtiiplantations in Saihanba of Hebei, China. Acta Ecologica Sinica, 2015, 35(16): 5403-5411. 张树梓, 李梅, 张树彬, 等. 塞罕坝华北落叶松人工林天然更新影响因子. 生态学报, 2015, 35(16): 5403-5411.
[20] Pérez-Harguindeguy N, Díaz S, Garnier E,etal. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany, 2013, 61(3): 167-234.
[21] Cornelissen J H C, Lavorel S, Garnier E,etal. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335-380.
[22] Lu R K. Soil and Agricultural Chemistry Analysis[M]. Beijing: Science and Technology of China Agriculture Press, 2000. 鲁如坤. 土壤农业化学分析方法[M]. 北京: 中国农业科技出版社, 2000.
[23] Mason N W H, Mouillot D, Lee W G,etal. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118.
[24] Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301.
[25] Laliberté E, Legendre P. A distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305.
[26] Morin P J. Biodiversity’s ups and downs. Nature, 2000, 406: 463-464.
[27] Currie D J, Francis A P. Regional versus climatic effect on taxon richness in angiosperms: reply to Qian and Ricklefs. American Naturalist, 2004, 163(5): 780-785.
[28] Perroni-Ventura Y, Montaa C, García-Oliva F. Relationship between soil nutrient availability and plant species richness in a tropical semi-arid environment. Journal of Vegetation Science, 2006, 17(6): 719-728.
[29] Bai Y F, Li L H, Wang Q B,etal. Changes in plant species diversity and productivity along gradients of precipitation and elevation in the Xilin River Basin, Inner Mongolia. Chinese Journal of Plant Ecology, 2000, 24(6): 667-673. 白永飞, 李凌浩, 王其兵, 等. 锡林河流域羊草草原植物种群和功能群的长期变异性及其对群落稳定性的影响. 植物生态学报, 2000, 24(6): 667-673.
[30] La Q, Zhaxi Ciren, Zhu W D,etal. Plant species-richness and association with environmental factors in the riparian zone of the Yarlung Zangbo River of Tibet, China. Biodiversity Science, 2014, 22(3): 337-347. 拉琼, 扎西次仁, 朱卫东, 等. 雅鲁藏布江河岸植物物种丰富度分布格局及其环境解释. 生物多样性, 2014, 22(3): 337-347.
[31] Markesteijn L, Poorter L, Bongers F. Light-dependent leaf trait variation in 43 tropical dry forest tree species. American Journal of Botany, 2007, 94(4): 515-525.
[32] Devictor V, Mouillot D, Meynard C,etal. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: the need for integrative conservation strategies in a changing world. Ecology Letters, 2010, 13(8): 1030-1040.
[33] Mayfield M M, Boni M F, Daily G C,etal. Species and functional diversity of native and human-dominated plant communities. Ecology, 2005, 86(9): 2365-2372.
[34] Kong B B, Wei X H, Du J L,etal. Effects of clipping and fertilization on the temporal dynamics of species diversity and functional diversity and their relationships in an alpine meadow. Chinese Journal of Plant Ecology, 2016, 40(3): 187-199. 孔彬彬, 卫欣华, 杜家丽, 等. 刈割和施肥对高寒草甸物种多样性和功能多样性时间动态及其关系的影响. 植物生态学报, 2016, 40(3): 187-199.
[35] Zang Y M, Zhu Z H, Li Y N,etal. Effects of species diversity and functional diversity on primary productivity of alpine meadow. Chinese Journal of Ecology, 2009, 28(6): 999-1005. 臧岳铭, 朱志红, 李英年, 等. 高寒矮嵩草草甸物种多样性与功能多样性对初级生产力的影响. 生态学杂志, 2009, 28(6): 999-1005.
Herbaceous plant species diversity and functional diversity in the forest-steppe zone of Hebei, China
GE Zhao-Xuan1, SUN Guo-Long2, YUAN Ye1, HUANG Xuan-Rui2, ZHANG Zhi-Dong1*
1.HebeiProvinceKeyLaboratoryofForestTreesGermplasmResourcesandForestProtection,CollegeofForestry,HebeiAgriculturalUniversity,Baoding071000,China; 2.SaihanbaMechanizedForestryCentreofHebeiProvince,Weichang068466,China
Understanding the relationships between environmental/soil factors and plant species diversity and functional diversity is useful for identifying which factors have the strongest effects on plant communities. In this study, we first studied the relationships between different components of herbaceous plant species diversity (species richness, Shannon-Wiener diversity, and Pielou evenness) and functional diversity indices (functional richness, functional evenness, and functional dispersion). Then, we used methods of analysis of similarities (ANOSIM) and redundancy analysis to explore their variation across three vegetation types (grassland, shrub, and natural secondary forest) and different environmental gradients. The results showed that species richness and Shannon-Wiener diversity were significantly higher in grassland than in shrub and natural secondary forest (P<0.05); however, the lowest functional richness was in grassland (P<0.05) and the lowest functional evenness and functional dispersion were in shrub forests (P<0.05). There was no significant difference in Pielou evenness among the three vegetation types (P>0.05). Functional richness was negatively related to species richness and Shannon-Wiener diversity (P<0.01). Functional dispersion was significantly positively correlated with species evenness (P<0.01), whereas functional evenness had a non-significant correlation with all species diversity indices (P>0.05). The main factors affecting species diversity were soil water content, soil organic matter, soil pH, soil total nitrogen content, and aspect. The main factors correlated with functional diversity were soil organic matter, soil depth, soil water content, and altitude. Consequently, soil factors were the vital determinants of plant species diversity and functional diversity in the study area, and accounted for 29.1% and 29.8% of the total variance explained by environmental factors, respectively. The results showed that herbaceous plant species diversity and functional diversity varied independently in the study area. Species diversity was mainly determined by resource availability, whereas functional diversity was mainly related to niche differentiation (the effects of both environmental filters and species interactions).
species diversity; functional diversity; environmental factors; vegetation types; variation partitioning
10.11686/cyxb2017015
2017-01-18;改回日期:2017-03-15
国家自然科学基金项目(31370636)和林业公益性行业科研专项(20150430304)资助。
葛兆轩(1990-),女,河北唐山人,在读硕士。E-mail: zhxuan_ge@126.com
*通信作者Corresponding author. E-mail: zhzhido@163.com
http://cyxb.lzu.edu.cn
葛兆轩, 孙国龙, 袁业, 黄选瑞, 张志东. 河北省森林草原区草本植物物种多样性和功能多样性. 草业学报, 2017, 26(7): 35-44.
GE Zhao-Xuan, SUN Guo-Long, YUAN Ye, HUANG Xuan-Rui, ZHANG Zhi-Dong. Herbaceous plant species diversity and functional diversity in the forest-steppe zone of Hebei, China. Acta Prataculturae Sinica, 2017, 26(7): 35-44.
