互花米草引种及遗传多样性研究进展
2017-07-03路峰,王昕
路 峰,王 昕
(1.山东黄河三角洲国家自然保护区黄河口管理站,山东 东营 257091;2.北京师范大学环境学院,北京 100875)
1 米草属起源及分类
互花米草 (Spartina alterniflora) 为禾本科(Gramineae)米草属(Spartina)的一种多年生六倍体植物[1]。目前有记录的米草属植物一共有15种[2],如表1所示。根据其染色体数目可以分为两大类[3](详见图1)。其中六倍体植物 (2n=6x=60,62)包括S.foliosa、S.maritima及其姐妹种 S.alterniflora, 四倍体植物 (2n=4x=40)包括S.argentinensis,S.arundinacea,S.bakeri,S.ciliata,S.cynosuroides,S.gracilis,S.patens,S.pectinata and S.spartinae[4]。
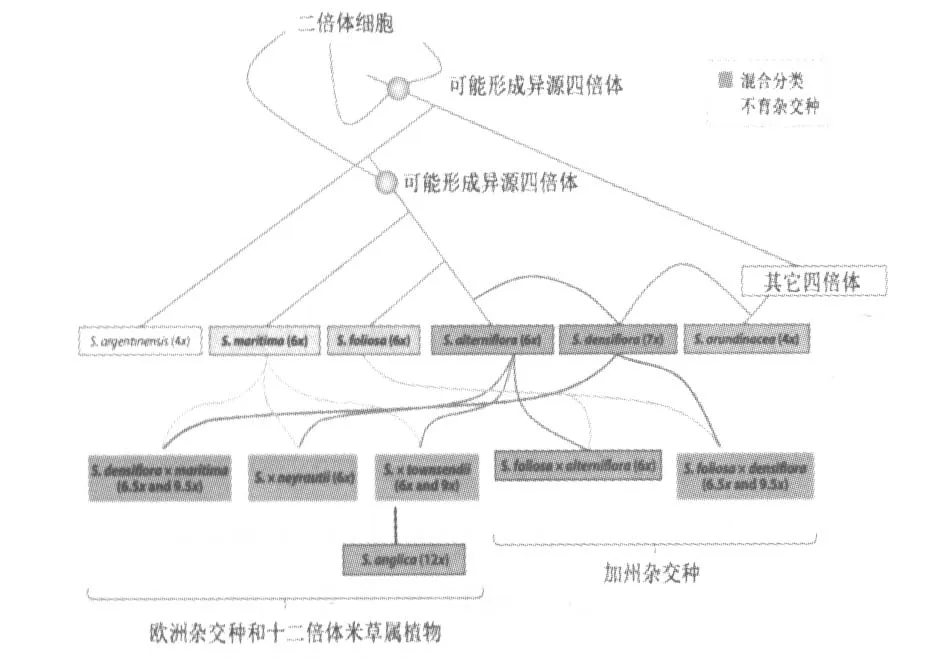
图1 米草属植物的杂交情况[3]

表1 米草属植物种类[2]
2 互花米草在国外的引种情况
互花米草原产于美洲大西洋西海岸及墨西哥湾,分布于北起加拿大魁北克和纽芬兰,南至美国佛罗里达和德克萨斯的沿海地区[5]。
2.1 在欧洲的引种情况
互花米草于19世纪初引入英国南安普敦(Southampton Water)地区[6]和法国西南部靠近昂达伊(Hendaye)的区域[7],然而互花米草在当地的引种并不成功,最终结果是与当地的欧米草杂交产生后代[8]。1870年在英国海斯镇(Hythe)出现了不育的杂交种S.×townsendii;1892年人们在法国昂达伊附近发现了一种新的不育杂交种S.×neyrautii。这两个杂交种的都是以互花米草为母本,S.×townsendii在形态学上介于互花米草和欧米草之间,S.×townsendii则更接近互花米草[7]。Spartina×townsendii在南安普敦生长旺盛,并在十九世纪广泛传播到世界各地[9];而Spartina×neyrautii则分布范围较小,也没有像Spartina×townsendii那样大规模传播开来[7]。
2.2 在美国西海岸的引种情况
互花米草在美国西海岸主要分布于加利福尼亚州(California)的旧金山湾(San Francisco Bay)和华盛顿州(Washington)的威拉帕湾(Willapa Bay)。
在20世纪70年代初期,作为湿地生境恢复计划的一部分,美国陆军工程兵团将互花米草引入旧金山湾[10],并与当地的叶米草混植。与叶米草相比,互花米草更高大,具有密集的分蘖,根系能够深入土壤深层,适应性更强。互花米草引入后与当地叶米草产生杂交种S.alterniflora×S.foliosa。该杂交种在株高、茎宽[11]、根系侧向扩张力、种子活力、花粉活力和耐盐性[12]都与亲本有一定差异。经研究该杂交种产生的花粉量是当地叶米草的400倍,当地90%的米草属花粉是由该杂交种产生的。在1998-2004年间,该杂交种占当地米草属播种繁殖的80%,叶米草占18%,互花米草不足2%,由此可见是该杂交种而并非互花米草导致互花米草引种后当地生态环境发生的一系列变化[12]。
互花米草于1894年被无意引进到华盛顿州的威拉帕湾。当时互花米草的种子和根系混杂在东方牡蛎(eastern oysters)中,通过铁路从美国东海岸运输到西海岸,最早只在当地的7个牡蛎养殖区域有零星分布[13]。随后的50年中互花米草的蔓延极其缓慢,然而自20世纪40年代互花米草在威拉帕湾首度开花之后[14],其扩散迅速加快,在2003年互花米草的生长高峰期,威拉帕湾的互花米草约3500公顷。
3 互花米草在国内的引种情况
3.1 引种目的与历史
互花米草的根系坚韧、直立,能够减缓水流的冲刷,降低水流速度,对潮水起到显著的消浪作用,其根和根状茎可以沉淀潮水中的矿物质,促进矿物质和有机质的沉淀淤积[15]。其促淤造陆作用明显,这是我国引种互花米草的主要目的[2]。
我国于1979年从原产地美国进口了5kg互花米草种子和60株互花米草个体。这60株个体来自3个地区即北卡罗来纳州的莫尔黑德城(Morehead City,North Carolina)、乔治亚州的奥尔塔马霍河河口(Altamaha Estuary,Georgia)和佛罗里达州的坦帕湾(Tampa Bay,Florida),分别代表3种不同的生态型。种子则采于北卡罗来纳州。随后在南京大学的田间和实验室条件下对互花米草进行栽培和播种,并将其引种到福建罗源的试验地[16]。人们根据其生理和形态特征区分不同来源的互花米草种群,并引种至从天津到广东的东部沿海各地区[17][18]。
3.2 互花米草在我国的分布范围
互花米草引入我国已有近40年的时间,由于其抗胁迫能力较强,生长迅速,在保滩护岸、控制污染等方面取得了一定的生态和经济效益[19][20]。目前其分布范围已扩大到北起辽宁,南达广西,成为盐沼湿地的优势类群,在我国东部海岸跨越了近22个纬度,使我国成为亚洲成功引种米草的第一个国家,其扩散规模远大于世界上其他地区[21]。
4 互花米草遗传多样性研究进展
4.1 国外研究进展
遗传多样性又称基因多样性,是生物多样性的重要组成部分[22]。它是指不同物种间或者同种的种群内、种群间在核苷酸、基因、染色体或整个基因组水平上的差异,主要来自基因流、突变、遗传漂变和自然选择[23]。
目前国外互花米草遗传多样性以Michael et al[24]研究的较为全面。他在2007年收集了互花米草在北美洲作为本地种(美国东海岸和墨西哥湾)和入侵种(美国西海岸)共有598个样本(具体采样地如图2所示),根据叶绿体DNA序列将样本分为4种单倍型即 A、B、C、D 如图 3所示。
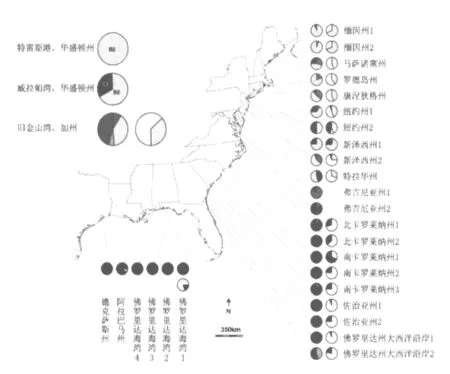
图2 美国互花米草采样地分布[24]
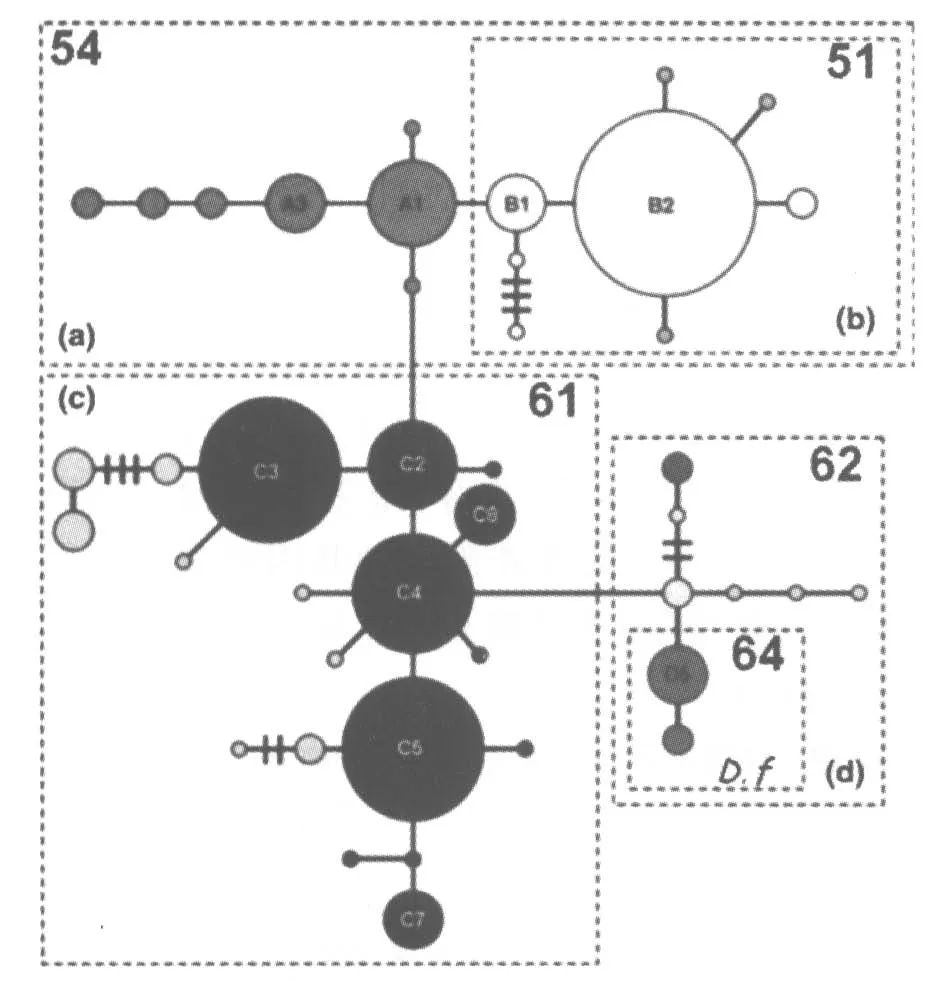
图3 美国互花米草单倍型分类[24]
4.1.1 美国东海岸和墨西哥湾
该区域互花米草的基因流和基因变异存在一定的地理模式,可大致划为“北”和“南”两种生态型。由于该区域单倍型群体之间的分布范围是重叠的,因而互花米草整体多样性并没有沿纬度梯度向北发生显著降低,但对某一单倍型来说其分布却不均匀。单倍型C几乎遍布所有的采样区,而其他类型的互花米草分布范围较小。在单倍型A和B占优势切萨皮克湾北部(Chesapeake Bay),单倍型C处于相对弱势状态,而单倍型D在低纬度地区间歇出现。在沿大西洋海岸出现的三个主要的单倍型群中,单倍型B的地理分布范围最小,只在特拉华和缅因州之间的采样点存在,与单倍型A相比其地理分布范围更广泛。单倍型A和B相对丰度的差异则表明B2是特拉华州和缅因州最常见的单倍型。单倍型C的出现频率的沿着纬度逐渐变化,在低纬度地区出现频率峰值,最多可有5组C单倍型共存;随着纬度升高其多样性频率逐渐下降,以C3和C4为主。其中稀有单倍型的出现也促进了单倍型C的向高纬度地区延伸分布[24]。
4.1.2 美国西海岸
旧金山湾的互花米草其遗传多样性水平相对较高,A,B,D和Df可在该地区共存。其中Df单倍型的个体是互花米草与叶米草的杂交种 S.alterniflora×S.foliosa。在威拉帕湾发现了两种单倍型B2和C3的互花米草,其分布较广泛,其中B2的出现频率是C3的两倍。在所有的样点中,只有新泽西州、纽约州和威拉帕湾出现B2和C3共存的情况,这表明威拉帕湾的互花米草可能最初起源于大西洋中部沿岸。但也不排除威拉帕湾的互花米草分别从中大西洋(盛产单倍型B2)和南大西洋(盛产单倍型C3)传入的可能性。特雷斯港(Grays Bay)互花米草的单倍型多样性水平最低,在采样地只发现一种单倍型B2。这一发现证明一个假说:特雷斯港互花米草来源于其邻近的威拉帕湾,通过潮汐交换将种子带入特雷斯港[24]。
4.2 国内研究进展
近年来随着互花米草在中国沿海地区的迅速蔓延,中国学者关于互花米草遗传多样性等方面的研究也越来越多(表2)。目前关于遗传多样性的研究手段有形态学标记、细胞学标记、生化标记和分子标记[25]。其中前三者在研究植物的遗传多样性时存在着很大的不确定性和局限性[26],而分子标记是以个体间遗传物质内核苷酸序列变异为基础的遗传标记,是DNA水平遗传多态性的直接的反映,是研究遗传多样性最理想的方法,已被广泛应用到植物遗传多样性等方面的研究[27]。
目前分子标记分为两种方法,一种是以southern杂交为基础的标记,如限制性片段长度多态性(Restriction Fragment Length Polymorphism,RFLP)、随机扩增多态性 DNA(Random Amplified Polymorphic DNA,RAPD)、简单重复序列区间多态性(Inter simple sequence repeat,ISSR)、简单序列重复长度多态性(微卫星 DNA,Simple sequence repeat,SSR)、扩增片段长度多态性 (Amplified fragment length polymorphism,AFLP)等;另一种是以特定序列为核心的标记,包括叶绿体基因(cpDNA)、核基因(nrDNA)及线粒体基因(mtDNA)的序列测定[28]。现有的文章主要是通过AFLP、叶绿体和线粒体基因测序的标记方法研究互花米草的遗传多样性。
4.2.1 AFLP标记
AFLP具有可靠性好、重复性高、可信度强等特点,非常适合绘制指纹图谱及构建分子遗传连锁图谱等研究[29]。
Deng et al[30]最早通过该方法分析中国江苏省射阳、东台、大丰3个地区的互花米草遗传多样性。由于上述3个研究样地距离较近,种内和种间的遗传多样性都较低,作者认为种群内的遗传多样性和植株高度对外界环境较强的适应性是其迅速扩展的主要原因。Xia et al[31]通过AFLP标记分析中国东部沿海7个样地的互花米草遗传多样性较低,并将上述样地的互花米草分为3种生态型即佛罗里达生态型(Florida ecotype)、乔治亚生态型(Georgia ecotype)和北卡罗来纳生态型(North Carolina ecotype),作者认为这3种相对独立生态型的互花米草的引入以及人类干扰可能是其适应性较强进而造成入侵的主要原因;在亚热带地区应该控制最具繁殖力和入侵性的佛罗里达生态型互花米草的入侵,而在温度较高的地区则应控制最具竞争性的乔治亚生态型互花米草的入侵。
4.2.2 SSR标记
SSR标记是基于 PCR技术的分子标记,它具有在基因组中分布均匀、数量众多、重复性好、操作简单、多态性信息丰富、廉价等优点,被认为是检测种群内遗传多样性和种群间遗传分化的最有效标记[32]。
沈栋伟[33]利用SSR标记对中国沿海和美国南卡罗莱纳州的16个互花米草种群共477个样品进行分析,发现各个种群间存在地理隔离。不同样点互花米草种群遗传多样性指数相差比较大。美国南卡罗来纳州和上海崇明的种群遗传多样性最高,福建罗源、江苏盐城和江苏启东次之,上海金山、福建厦门、山东潍坊和山东日照的种群遗传多样性最低。
4.2.3 叶绿体和线粒体基因测序
植物叶绿体基因组属于母系遗传,其结构简单、稳定,极少或不发生重组和种内变异,但某些基因间隔区序列进化速度较快,为植物的遗传进化分析提供了大量的信息[34]。植物线粒体基因组在核苷酸序列上的进化相对较慢,而基因排列的进化又相当快,因而植物线粒体基因组测序在植物遗传多样性研究中的应用并不多[35]。
吴娟子等[36]测定了中国沿海互花米草8个种群共80个样本的叶绿体trnT-trnF序列,研究结果表明互花米草在福建罗源的单倍型多样性高于浙江、上海、江苏、山东和天津种群;此外中国互花米草种群受瓶颈效应影响明显,种群间遗传变异较低,种群内多样性相对较高。宫璐等[37]对目前中国沿海10个省(市)共20个总群的互花米草通过扩增互花米草其叶绿体trnT-trnL和线粒体nad1两个基因片段,发现中国沿海地区互花米草遗传多样性低,中国南部、中部和北部3大种群没有发生明显遗传分化,推测可能是表型可塑性在3大种群的表型变异上起了更大作用。Guo etal[38]将中国与美国的互花米草通过叶绿体DNA测序进行遗传多样性比较,认为中国沿海互花米草大部分起源于大西洋,而部分香港地区互花米草个体可能来源于大西洋和墨西哥湾。
除了分析中国东部沿海互花米草的遗传多样性外,中国学者还关注其遗传多样性对入侵能力的影响。Wang etal[39]通过对崇明东滩的22丛6个基因型的互花米草设置3个水平的同质园试验,发现不同基因型平均株高、直径、节茎、叶鞘厚度、穗分枝长存在显著差异,互花米草的基因型多样性效应随时间逐渐增强,基因型多样性显著提高互花米草的入侵能力。

表2 中国学者关于互花米草基因型多样性的研究
5 结语与建议
本文论述了互花米草的引种历程、分布范围及遗传多样性,鉴于互花米草在中国表现出一定的入侵性,笔者认为未来的研究应该着重从如下几个方面进行:
5.1 尽快开展对互花米草的风险评估工作
从互花米草的生长、繁殖、生存条件、引进用途、控制危害的费用等方面进行综合评价,确定风险大小和风险管理措施,也为制定外来物种引进政策提供科学依据。
5.2 深入对互花米草入侵预测的研究
目前相关的研究较少,今后应加强对互花米草入侵的生态学规律、动态监测、扩散机制等方面的研究,明确互花米草下一步可能入侵的区域,并采取相关的管理防治措施,从而达到生态预警的作用。
5.3 加强对互花米草生物防治的研究
目前国内主要通过刈割、水淹、掩埋等物理方法来治理互花米草,但出于成本和后果的考虑,从长远角度来看还应选择生物防治对互花米草进行控制。前人的研究显示,在长江河口的芦苇对互花米草的再生长起到一定的抑制作用[40],在广东珠海淇澳岛引进无瓣海桑来控制互花米草也有明显的效果[41],笔者认为应加强此方面的研究,从而促进我国被入侵区域的生态系统修复。
5.4 加强对互花米草正效应的研究。互花米草在饲养禽畜、水产饵料等方面具有一定的功效,因而应充分发掘其经济价值,使其变废为宝。
[1]J Ferreira de Carvalho, J Poulain, C Da Silva, P Wincker,S Michon-Coudouel, A Dheilly, D Naquin, J Boutte,A Salmon and M Ainouche. Transcriptome de novo assemblyfrom next -generation sequencing and comparativeanalyses in the hexaploid salt marsh species Spartinamaritima and Spartina alterniflora [J]. Heredity, 2013,110(2): 181-193.
[2]苑泽宁.互花米草结构与生殖生态学研究.南开大学博士论文.2008.
[3]Ainouche M L,Fortune P M,Salmon A,Parisod C,Grandbastien M A,Fukunaga K,Ricou M,Misset M T.Hybridization,polyploidy and invasion:lessons from Spartina (Poaceae)[J].Biological Invasions,2009,11:1159-73.
[4]Donald R.Strong and Debra R.Ayres.Ecological and Evolutionary,Misadventures of Spartina [J].Annual Review of Ecology,Evolution and Systematics,2013,44:389-410.
[5]Daehler C C,Strong D R.Status prediction and prevention of introduced cordgrass Spartina spp.invasions in Pacific estuaries,USA[J].Biological Conservation,1996,78:51-58.
[6]Ayres D R and Strong D R.Origin and genetic diversity of Spartina anglica (Poaceae)using nuclear DNA markers[J].American Journal of Botany,2001,88:1863–67.
[7]Baumel A,Ainouche M L,Misset M T,Gourret J P,Bayer R J.Genetic evidence for hybridization betweenthe native Spartinamaritima and the introduced Spartina alterniflora (Poaceae)in South-West France:Spartina×neyrautii re-examined[J].Plant Systematics and Evolution,2003,237:87-97.
[8]Marchant C J and Goodman P J. Biological flora of theBritish Isles, Spartina alterniflora Loisel [J]. Journal ofEcology, 1969, 57: 291-95.
[9]Renny-Byfield S,Ainouche M,Leitch I J,Lim K Y,Le Comber SC,Leitch A R.Flow cytometry and GISH reveal mixed ploidy populations and Spartina nonaploids with genomes of S.alterniflora and S.maritima origin[J].Annals of Botany,2010,105:527-33.
[10]Strong D R,Ayres D A.Spartina introductions and consequences in saltmarshes:arrive,survive,thrive,and sometimes hybridize.In Human Impacts on Salt Marshes:A Global Perspective,ed.B R Silliman,M D Bertness,E D Grosholz [M].Berkeley:University of California Press,2009:3-22.
[11]Ayres D R,Smith D L,Zaremba K,Klohr S,Strong D R.2004.Spread of exotic cordgrasses and hybrids(Spartina sp.)in the tidal marshes of San Francisco Bay,California,USA [J].Biological Invasions,2004,(6):221-31.
[12]Ayres D R,Zaremba K,Sloop CM,Strong D R.Sexual reproduction of cordgrass hybrids(Spartina foliosa×alterniflora)invading tidalmarshes in San Francisco Bay[J].Diversity and Distributions,2008,(14):187-95.
[13]Civille JC,Sayce K,Smith SD,Strong D R.2005.Reconstructing a century of Spartina alterniflora invasion with historical records and contemporary remote sensing[J].Ecoscience,2005,(12):330-38.
[14]Sayce K.Spartina in Willapa:a case history.In Spartina Workshop Record,ed.TFMumford,P Peyton,JR Sayce,SHarbell[M].Seattle:Wash.Sea Grant Program,1990:27-30.
[15]Bouma T J,De Vries M B,Herman PM J.Comparing ecosystem engineering efficiency of two plant species with contrasting growth strategies[J].Ecology,2010,91(9):2696-2704.
[16]AnSQ,GuBH,ZhouCF,WangZS,DengZF,ZhiYB,LiHL,Chen L,Yu D H,Liu Y H.Spartina invasion in China:implications for invasive species management and future research[J].Weed Research,2007,47:183-191.
[17]Chung C H.Thirty years of ecological engineering with Spartina plantations in China[J].Ecological Engineering,1993,(2):261-289.
[18]Chung C H.Creation of Spartina plantations for reclaiming Dongtai,China,tidal flats and offshore sands[J].Ecological Engineering,2004,23:135-150.
[19]朱晓佳,钦佩.外来种互花米草及米草生态工程 [J].海洋科学,2003,27(12):l-19.
[20]曹大正,王银生,张冬然,顾立军,唐廷贵.互花米草在吹填筑挡工程上的试验与应用 [J].中国工程科学,2005,7(7):14-23.
[21]邓自发,安树青,智颖飙,周长芳,陈琳,赵聪蛟,方淑波,李红丽.外来种互花米草入侵模式与爆发机制 [J].生态学报,2006,(8):2678-2686.
[22]Millar C I,Libby W J.Strategies for consvering clinal,eoctypic and disjunet population diversity in widespread species.In:Falk D A,Holsiner KE.Genetics and Conservation of Rare Plants [M].New York:Oxford University Press,1991:149-170.
[23]程蓓蓓.中国红豆杉属分子谱系地理学与遗传多样性研究[D].中国林业科学研究院博士论文,2016.
[24]Michael J.Blum.,K.Jun Bando,M.Katz,Donald R.Strong.Geographic structure,genetic diversity and source tracking of Spartina alterniflora [J].Joural of Biogeography,2007,34:2055-2069.
[25]余亚莹.国内外香稻资源遗传多样性研究 [D].中国农业科学院硕士论文,2015.
[26]方向民.珍稀植物太行花的遗传多样性研究 [D].河南农业大学硕士论文,2010.
[27]冯涛.新疆野苹果部分表型性状遗多样性研究 [D].山东农业大学硕士论文,2007.
[28]狄晓艳.油松遗传多样性与光合生理生态特性研究[D].山西大学博士论文,2014.
[29]于海萍.牡丹SSR分子标记的开发及其在亲缘关系分析中的应用[D].北京林业大学硕士论文,2013.
[30]Deng Z F,An S Q,Zhou C F,Wang Z S,Zhi Y B,Wang Y J,Shi SH,Chen L,Zhao C J.Genetic structure and habitat selection of the tall form Spartina alterniflora Loisel.In China [J].Hydrobiologia,2007,583:195-204.
[31]Xia L,Zhao H,Yang W,An S Q.Genetic Diversity,Ecotype Hybrid,and Mixture of Invasive Spartina alterniflora Loisel in Coastal China [J].CLEAN-Soil,Air,Water,2015,43(12):1672-1681.
[32]Ellegren H.Microsatellites:simple sequences with complex evolution[J].Nature reviews genetics,2004,5(6):435-445.
[33]沈栋伟.互花米草基因型多样性及其与入侵能力的关系[D].华东师范大学硕士论文.2007.
[34]Green B R.Chloroplast genomes of photosynthetic eukaryotes[J].Plant Journal,2011,66(1):34-44.
[35]Adams K L,Palmer J D.Evolution of mitochondrial gene content:gene loss and transfer to the nucleus[J].Molecular Phylogenetics and Evolution,2003,29,380-395.
[36]吴娟子,王强,钟小仙,陈建群.根据cpDNA trnT-trnF序列变异分析中国互花米草种群的遗传结构[J].草业学报,2012,21(4):134-140.
[37]宫璐,李俊生,柳晓燕,赵相健,邓贞贞,赵彩云.中国沿海互花米草遗传多样性及其遗传结构 [J].草业科学,2014,31(7):1290-1297.
[38]Guo W,Qiao S,Wang Y,Shi S,Tan F,Huang Y.Genetic diversity,population structure,and genetic relatedness of native and non-native populations of Spartina alterniflora(Poaceae,Chloridoideae)[J].Hydrobiologia,2015,745(1):313-327.
[39]Wang X Y,Shen DW,Jiao J,Xu N N,Yu S,Zhou X F,Shi M M,Chen X Y.Genotypic diversity enhances invasive ability of Spartina alterniflora[J].Molecular ecology,2012,21(10):2542-2551.
[40]Li H P,Zhang L Q.An experimental study on physical controls of an exotic plant Spartina alterniflora in Shanghai,China [J].Ecological Engineering,2008,32(1):11-21.
[41]唐国玲,沈禄恒,翁伟花,章家恩,廖宝文,刘金苓,滕兴顺.无瓣海桑对互花米草的生态控制效果 [J].华南农业大学学报,2007,28(1):10-13.
