饲粮高铁对肉仔鸡十二指肠黏膜铁转运载体基因表达及组织微量元素含量的影响
2017-06-24邹亚学王秋悦牛一兵唐家明张丽阳罗绪刚李素芬
邹亚学 王秋悦* 牛一兵 贺 英 唐家明 吕 林 张丽阳 罗绪刚 李素芬**
(1.河北科技师范学院,秦皇岛066000;2.中国农业科学院北京畜牧兽医研究所,北京100093)
饲粮高铁对肉仔鸡十二指肠黏膜铁转运载体基因表达及组织微量元素含量的影响
邹亚学1王秋悦1*牛一兵1贺 英1唐家明1吕 林2**张丽阳2罗绪刚2李素芬1**
(1.河北科技师范学院,秦皇岛066000;2.中国农业科学院北京畜牧兽医研究所,北京100093)
本试验旨在研究饲粮铁含量对肉仔鸡组织重要微量元素铁、锰、铜、锌含量及十二指肠黏膜主要铁转运载体基因表达的影响,探讨铁对肉仔鸡微量元素吸收和代谢的影响及其机制。将336只1日龄商品代罗斯308肉公雏按照体重随机分成4个组,每组6个重复,每个重复14只鸡。对照组饲喂不额外添加铁的基础饲粮(实测铁含量为78 mg/kg),铁添加组分别饲喂以七水硫酸亚铁(FeSO4·7H2O)形式添加100、250或500 mg/kg铁的试验饲粮(实测铁含量分别为166、308和579 mg/kg)。试验期21 d。各组试鸡分别于7、14和21日龄屠宰分析肝脏、心脏、胰腺、十二指肠黏膜和胫骨灰中铁、锰、铜、锌含量及十二指肠黏膜中二价金属转运蛋白(DMT1)和膜铁转运蛋白(FPN1)mRNA表达水平。结果表明:1)500 mg/kg铁添加组1~7日龄和8~14日龄的平均日增重显著低于其他3组(P<0.10),250和500 mg/kg铁添加组1~7日龄的平均日采食量显著低于其他2组(P<0.10)。2)饲粮铁含量对肉仔鸡7、14、21日龄的血浆总铁结合力以及全血血红蛋白浓度(7日龄除外)和红细胞压积均无显著影响(P>0.10),但显著影响7、14和21日龄血浆铁含量和铁饱和度(P<0.10),二者均随饲粮铁含量增加而升高。3)7和14日龄心脏及7、14和21日龄肝脏、十二指肠黏膜、胰腺和胫骨灰铁含量均随饲粮铁含量的增加而升高,7、14和21日龄十二指肠黏膜、胰腺和胫骨灰锰含量均随饲粮铁含量的增加而降低;饲粮添加铁显著降低7日龄胰腺锌含量(P<0.10),但对其他日龄胰腺和各日龄其他所测组织锌含量以及各日龄所测各组织铜含量均无显著影响(P>0.10)。4)饲粮铁含量显著影响7、14和21日龄十二指肠黏膜DMT1和FPN1 mRNA表达水平(P<0.10),各日龄DMT1和FPN1 mRNA表达水平均随饲粮铁含量的增加而降低。以上结果提示,高铁饲粮可能通过调控十二指肠黏膜DMT1和FPN1基因的表达降低锰和锌在肠道的吸收,进而减少锰和锌在组织中的沉积。
铁;铁转运载体基因表达;组织微量元素含量;肉仔鸡
微量元素是动物机体必需的营养成分,作为体内酶或特定蛋白的组成成分在抗氧化和骨骼发育、供能、氧气运输和储存、DNA合成及蛋白质代谢等方面发挥着非常重要的作用。然而,由于某些微量元素在肠道吸收和代谢过程上的协同或拮抗作用,往往会导致一种元素的过量影响另外一种或多种元素的吸收和代谢。已有研究发现,存在于十二指肠黏膜上的二价金属转运蛋白(divalent metal transporter 1,DMT1)不仅可以转运铁,还可以转运锰、铜、锌等二价金属[1-2]。存在于基底膜上膜铁转运蛋白1(ferroportin 1,FPN1)除同时转运铁和锰跨过基底膜进入血液循环外[3],可能也参与锌、铜、钴和镉的转运[4]。断奶仔猪饲粮中铁含量过高时,十二指肠黏膜DMT1 mRNA表达水平降低,肝脏和十二指肠黏膜铁含量线性增高而锰含量线性降低[5]。断奶犊牛饲粮中铁含量过高时,十二指肠黏膜FPN1 mRNA表达水平降低,DMT1 mRNA表达水平有降低的趋势,肝脏铁含量提高,十二指肠黏膜和心脏铁含量有提高趋势,但仅有十二指肠黏膜锰含量显著降低,肝脏和心脏锰含量并未降低[6]。在肉仔鸡配合饲料加工过程中,大量铁的掺入常使饲粮含铁量远远超过其对铁的需要量,但在现有文献中尚未见到饲粮高铁对肉仔鸡肠道中铁转运载体基因表达及组织中微量元素含量影响的研究报道。因此,本试验通过观察饲粮铁含量对肉仔鸡组织重要微量元素铁、锰、铜、锌含量及十二指肠黏膜DMT1及FPN1 mRNA表达的影响,探讨铁对肉仔鸡微量元素吸收和代谢的影响及其机制,为提高肉仔鸡对微量元素的利用率提供试验依据。
1 材料与方法
1.1 饲粮配制
参照我国《鸡饲养标准》[7]中肉仔鸡营养需要量配制1~21日龄肉仔鸡玉米-豆粕型基础饲粮(表1,实测铁含量为78 mg/kg),并以七水硫酸亚铁(FeSO4·7H2O)为铁源,按照100、250和 500 mg/kg铁的添加量替代基础饲粮中等量的玉米淀粉,配制3种试验饲粮(实测铁含量分别为166、308和579 mg/kg)。
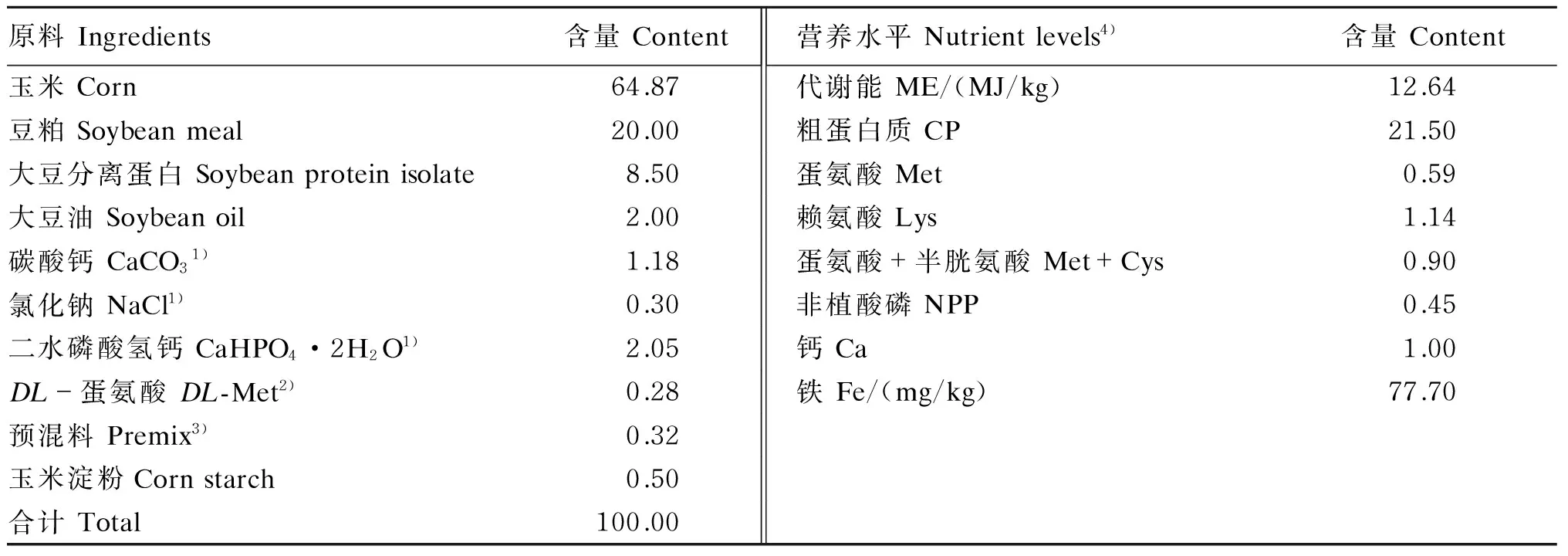
表1 1~21日龄肉仔鸡基础饲粮组成及营养水平(饲喂基础)Table 1 Composition and nutrient levels of the basal diet for broilers during 1 to 21 days of age (as-fed basis) %
1)试剂级Reagent grade。
2)饲料级Feed grade。
3)每千克饲粮含有Contained the following per kilogram of the diet:VA 12 500 IU,VD33 750 IU,VK32.5 mg,VE 20 IU,VB12.5 mg,VB28 mg,VB120.015 mg,VB62.5 mg,烟酸 nicotinic acid 32.5 mg,D-泛酸D-pantothenic acid 12.5 mg,生物素 biotin 0.125 mg,叶酸 folic acid 1.25 mg,胆碱 choline 700 mg,金霉素 chlortetracycline 50 mg,Cu (as copper sulfate) 8 mg,Zn (as zinc sulfate) 60 mg,Mn (as manganese sulfate) 100 mg,Se (as sodium selenite) 0.15 mg,I (as potassium iodide) 0.35 mg。
4)粗蛋白质、钙和铁为实测值,其他为计算值。CP, Ca and Fe were measured values, while the others were calculated values.
1.2 试验动物与饲养管理
将336只1日龄商品代罗斯308肉公雏按照体重随机分成4个组,每组6个重复,每个重复14只鸡。对照组饲喂不额外添加铁的基础饲粮,铁添加组分别饲喂3种试验饲料。试鸡以重复为单位饲养于不锈钢肉鸡笼中,自由采食和饮自来水(未检测到铁)。试鸡饲养管理按《罗斯肉仔鸡饲养管理手册》进行。每日观察并记录鸡只健康、死亡状况;每周末以重复为单位称试鸡空腹体重,并统计鸡只耗料量,计算平均日采食量、平均日增重、料重比及死亡率。试验期21 d。
1.3 样品采集与制备
禁食8 h后,分别于8、15日龄从每个重复中选取4只与平均体重相近的试鸡,21日龄从每个重复中选取2只与平均体重相近的试鸡,心脏穿刺采集乙二胺四乙酸(EDTA)抗凝血,4 ℃保存,用于测定血红蛋白浓度及红细胞压积;然后采集肝素钠抗凝血,3 000 r/min离心10 min后得血浆,于-20 ℃保存,用于测定血浆铁含量及总铁结合力(TIBC)。
将采过血的试鸡屠宰,用生理盐水冲洗十二指肠2次,然后在距幽门1 cm处剪开十二指肠,用载玻片刮取上部4~5 cm的黏膜,液氮速冻后于-80 ℃冻存,用于铁转运载体基因mRNA表达水平的测定;然后刮取下部10 cm左右的黏膜,取左侧肝脏、心脏、胰脏和左腿胫骨,-20 ℃冻存,用于测定铁、锰、铜和锌含量。
上述样品分析时均是将每个重复中屠宰鸡的样品等量合并为1个样品进行分析[8]。
1.4 样品分析
铁、锰、铜和锌含量:参照马新燕[9]采用的方法,将自来水、饲粮、十二指肠黏膜、肝脏、心脏、胰脏和胫骨灰用混酸(HNO3和HClO4按20∶1混合)湿消化后,用电感耦合等离子体发射光谱仪(Model IRIS Intrepid Ⅱ,Thermal Jarrell Ash,Waltham,MA)测定其中的铁、锰、铜和锌含量。
血液指标:采用全自动血液分析仪(HC-3000)测定全血红细胞压积及血红蛋白浓度。参照马新燕[9]的方法,采用比色法测定血浆铁含量和总铁结合力,试剂盒(No. A039和No. A040)购自南京建成生物工程研究所,计算血浆铁含量与总铁结合力的百分比即血浆铁饱和度(TS)。
DMT1和FPN1 mRNA表达水平测定:采用实时荧光定量PCR法,参照白世平[10]所述方法测定DMT1和FPN1 mRNA表达水平。主要操作步骤为:用Trizol试剂(No. 15596-026,Invitrogen)提取总RNA,用P330-31型核酸分析仪(Implen)测定细胞总RNA的浓度及其在260和280 nm处的吸光度(OD)值,计算OD260 nm和OD280 nm的比值,采用1%甲醛变性琼脂糖凝胶电泳检测总RNA的完整性。参照SuperScriptTMⅢ First-Strand Synthesis System for RT-PCR试剂盒(No. 205311,Qiagen)说明书所述步骤合成cDNA模板。采用SYBR Green染料试剂盒(No. 4367659,ABI)在ABI 7500 PCR仪上对DMT1和FPN1基因进行实时荧光定量PCR分析,以β-肌动蛋白(β-actin)基因作为内参基因。实时荧光定量PCR所用引物序列见表2,由上海英潍捷基公司合成。每个样品重复测定3次。
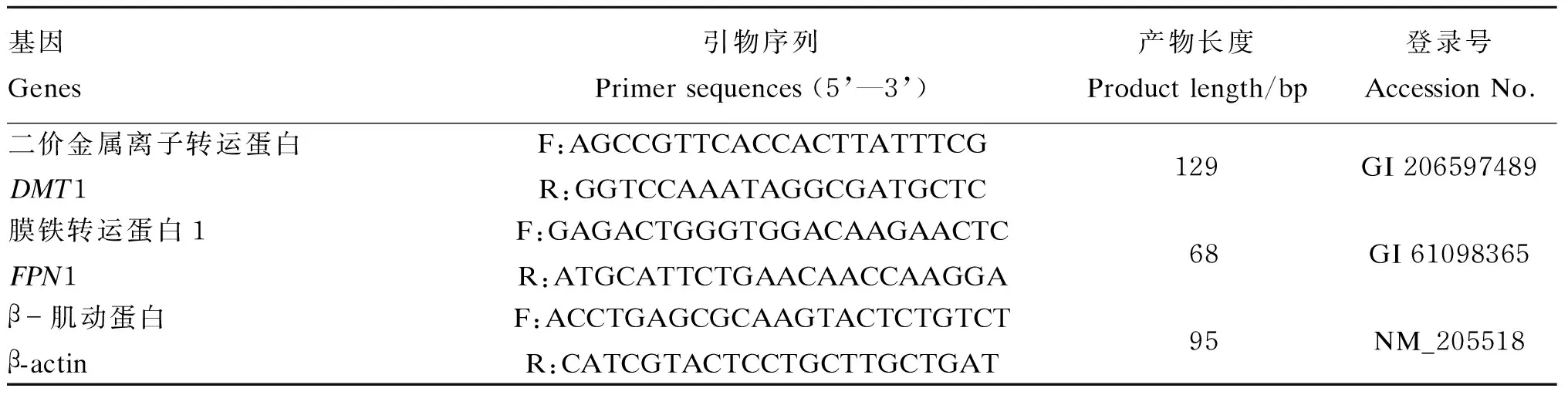
表2 实时荧光定量PCR引物序列Table 2 Primer sequences for real-time fluorescent quantitative PCR
1.5 数据统计与分析
采用SAS 8.0的一般线性模型(GLM)对所有试验数据进行方差分析。以重复作为统计分析的试验单元,若方差分析结果显示差异显著,则以最小显著差异(LSD)法比较各平均值之间的差异显著性。以国际文献报道中通常采用的P<0.10作为检验差异显著性标准[11-12]。
2 结果与分析
2.1 饲粮铁含量对1~21日龄肉仔鸡生长性能的影响
饲粮铁含量对1~21日龄肉仔鸡生长性能的影响结果列于表3。在整个试验期内,仅250 mg/kg铁添加组有1只鸡在14~21日龄阶段死亡,饲粮铁含量与死亡率之间没有直接相关性,故未将死亡率列入表3。饲粮铁含量除对肉仔鸡1~7日龄的平均日增重和平均日采食量及8~14日龄的平均日增重有显著影响(P<0.10)外,对其他生长阶段的各生长性能指标均无显著影响(P>0.10)。500 mg/kg铁添加组1~7日龄和8~14日
龄的平均日增重显著低于其他3组(P<0.10),其他3组间无显著差异(P>0.10)。250和500 mg/kg铁添加组1~7日龄的平均日采食量显著低于其他2组(P<0.10),100 mg/kg铁添加组与对照组间无显著差异(P>0.10)。

表3 饲粮铁含量对1~21日龄肉仔鸡生长性能的影响Table 3 Effects of dietary Fe content on growth performance of broilers during 1 to 21 days of age (n=6)
同列同一阶段数据肩标不同字母表示差异显著(P<0.10)。表4至表8同。
Values of the same stage with different letter superscripts in the same column differ significantly (P<0.10). The same as Table 4 to Table 8.
2.2 饲粮铁含量对1~21日龄肉仔鸡血液指标的影响
饲粮铁含量对1~21日龄肉仔鸡血液指标的影响结果列于表4。饲粮铁含量对7、14、21日龄血浆总铁结合力无显著影响(P>0.10),但显著影响血浆铁含量和铁饱和度(P<0.01),二者均随饲粮铁含量增加而升高。饲粮铁含量对7、14、21日龄全血红细胞压积及14、21日龄全血血红蛋白浓度均无显著影响(P>0.10),但显著影响7日龄全血血红蛋白浓度(P<0.01)。
2.3 饲粮铁含量对1~21日龄肉仔鸡组织铁、锰、铜和锌含量的影响
饲粮铁含量对1~21日龄肉仔鸡组织铁、锰、铜和锌含量的影响结果见表5至表8。饲粮铁含量除对21日龄心脏铁含量无显著影响(P>0.10)外,对7和14日龄心脏及各日龄其他所测组织铁含量均有显著影响(P<0.10),这些组织中铁含量均随饲粮铁含量增加而升高。饲粮铁含量对各日龄肝脏和心脏锰含量无显著影响(P>0.10),但对各日龄十二指肠黏膜、胰腺和胫骨灰锰含量有显著影响(P<0.10),这些组织中锰含量均随饲粮铁含量的增加而降低。饲粮铁含量对7日龄胰腺锌含量有显著影响(P<0.10),其随饲粮铁含量的增加而降低,饲粮铁含量对14和21日龄胰腺以及各
日龄其他所测组织锌含量无显著影响(P>0.10)。饲粮铁含量对各日龄所测各组织铜含量均无显著影响(P>0.10)。
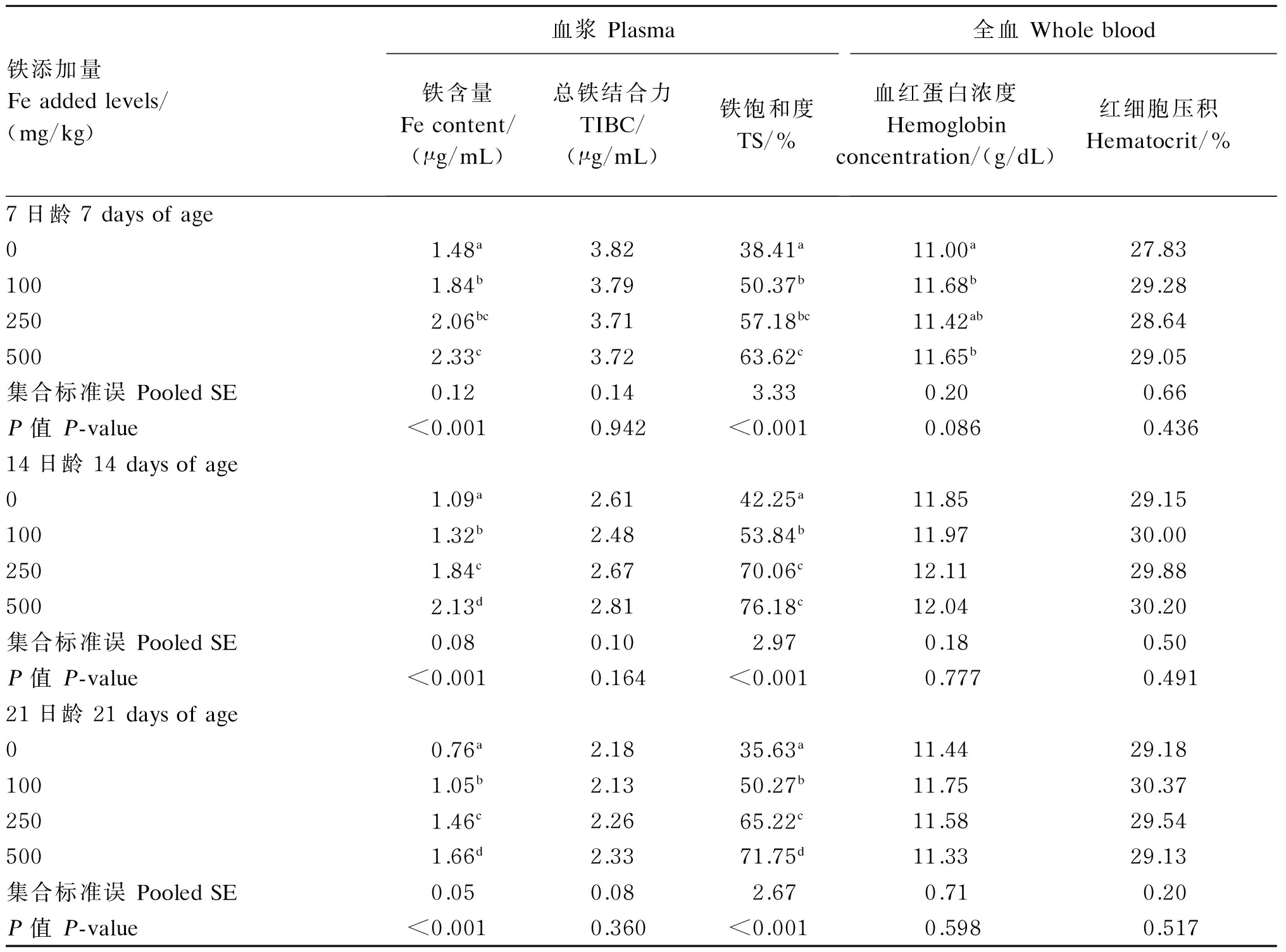
表4 饲粮铁含量对1~21日龄肉仔鸡血液指标的影响Table 4 Effects of dietary Fe content on blood parameters of broilers during 1 to 21 days of age
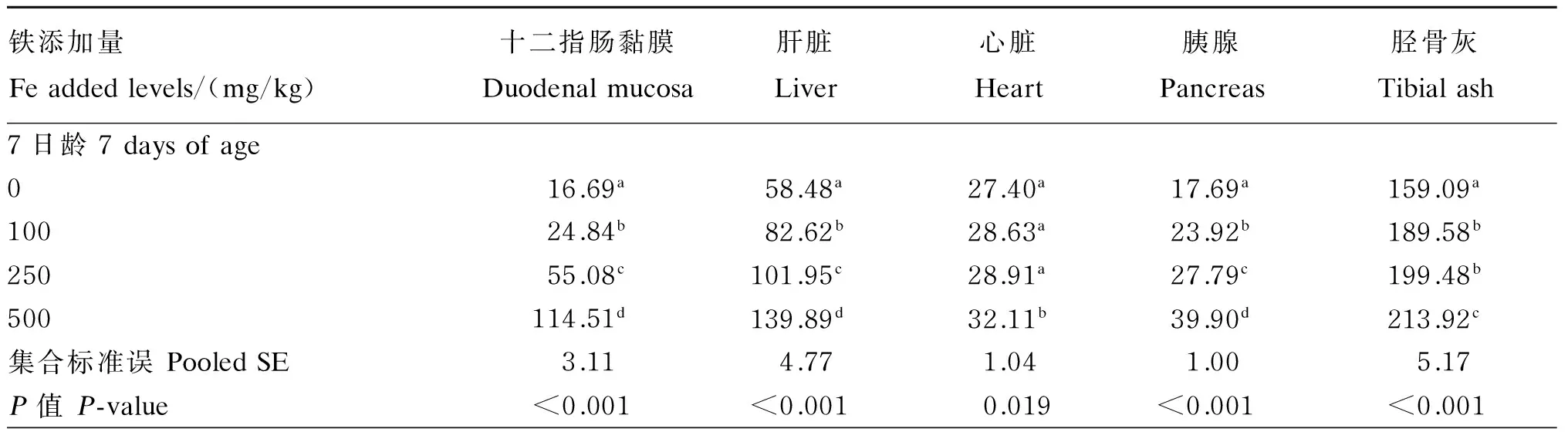
表5 饲粮铁含量对1~21日龄肉仔鸡组织铁含量的影响(鲜重基础)Table 5 Effects of dietary Fe content on Fe content in tissues of broilers during 1 to 21 days of age (fresh weight basis) μg/g

续表5铁添加量Feaddedlevels/(mg/kg)十二指肠黏膜Duodenalmucosa肝脏Liver心脏Heart胰腺Pancreas胫骨灰Tibialash14日龄14daysofage016.02a70.05a34.76a17.69a192.98a10027.10b78.32a36.28ab23.92b225.36b25065.52c90.37b38.29bc27.79c235.99bc500148.41d106.50c40.81c39.90d239.58c集合标准误PooledSE3.134.301.300.605.44P值P-value<0.001<0.0010.012<0.001<0.00121日龄21daysofage012.42a70.05a36.9513.25a192.09a10021.72b78.32a37.3316.03b222.21b25033.86c90.37ab37.2216.44b230.01bc50093.26d106.50b38.9518.45c238.08c集合标准误PooledSE2.924.731.380.716.54P值P-value<0.001<0.0010.736<0.001<0.001

表6 饲粮铁含量对1~21日龄肉仔鸡组织锰含量的影响(鲜重基础)Table 6 Effects of dietary Fe content on Mn content in tissues of broilers during 1 to 21 days of age (fresh weight basis) μg/g
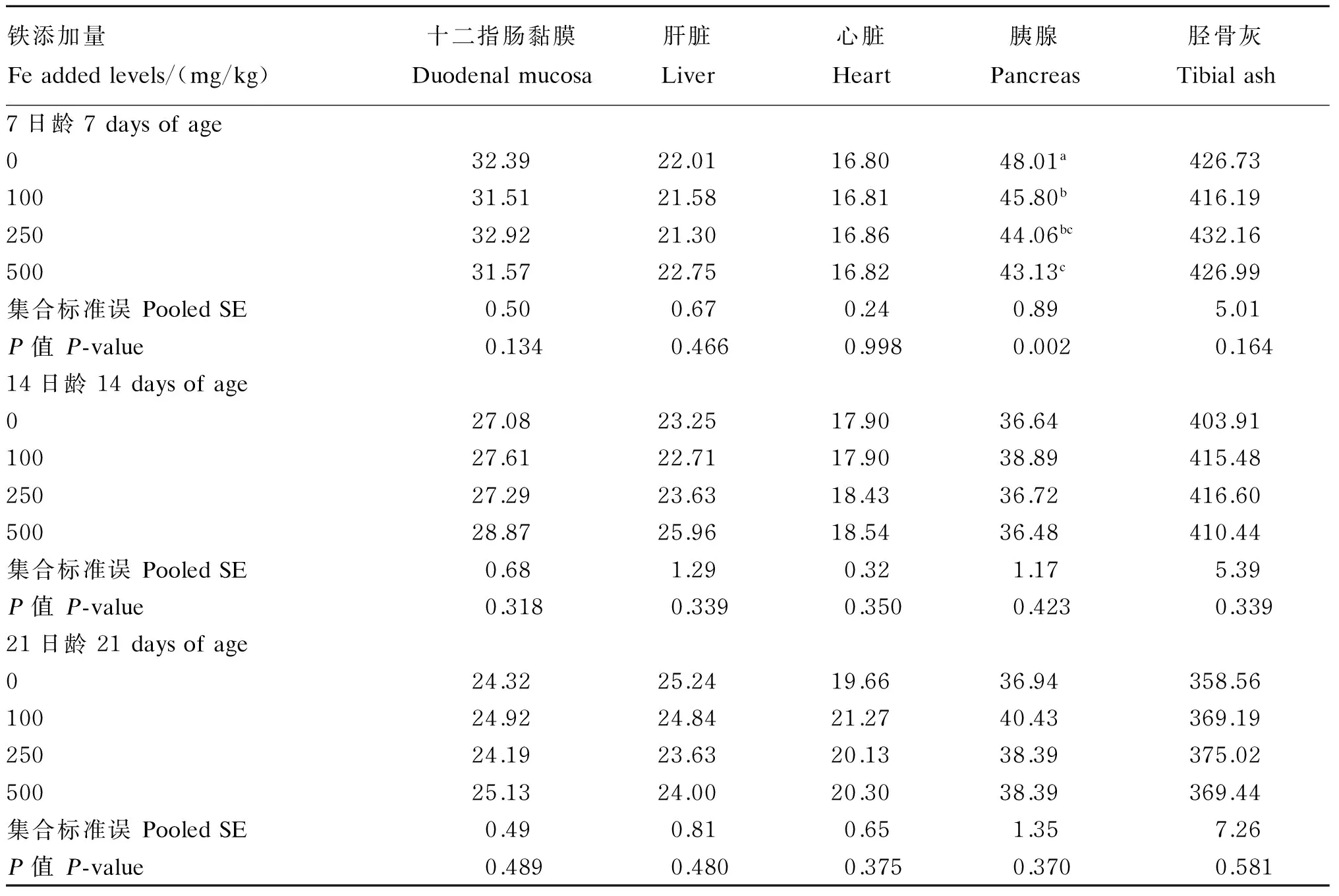
表7 饲粮铁含量对1~21日龄肉仔鸡组织锌含量的影响(鲜重基础)Table 7 Effects of dietary Fe content on Zn content in tissues of broilers during 1 to 21 days of age (fresh weight basis) μg/g

表8 饲粮铁含量对1~21日龄肉仔鸡组织铜含量的影响(鲜重基础)Table 8 Effects of dietary Fe content on Cu content in tissues of broilers during 1 to 21 days of age (fresh weight basis) μg/g
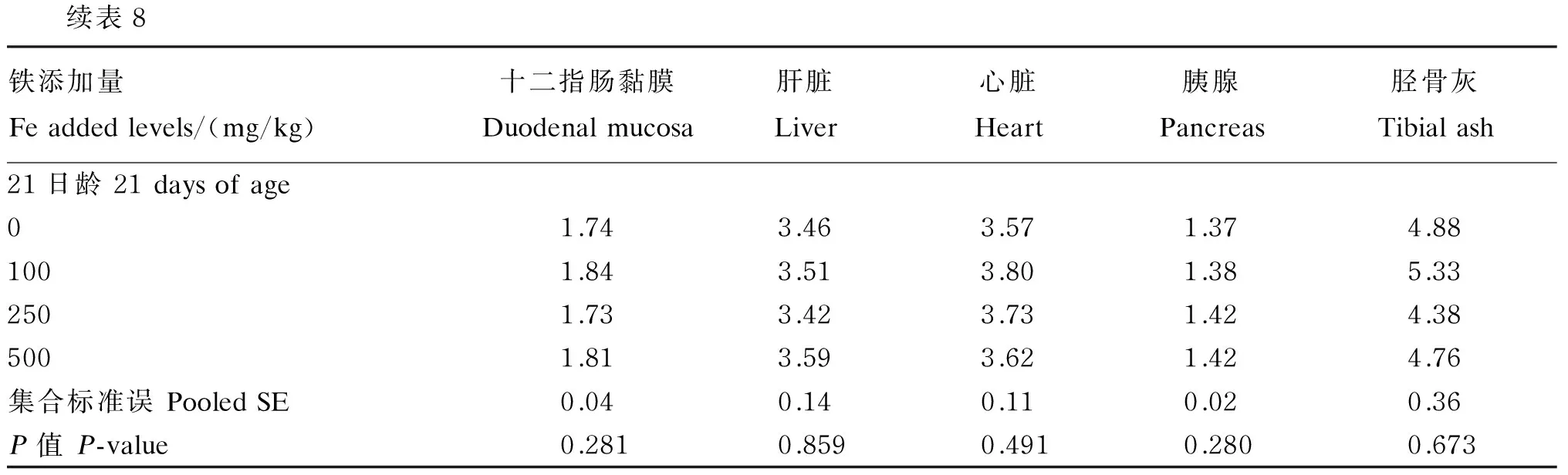
续表8铁添加量Feaddedlevels/(mg/kg)十二指肠黏膜Duodenalmucosa肝脏Liver心脏Heart胰腺Pancreas胫骨灰Tibialash21日龄21daysofage01.743.463.571.374.881001.843.513.801.385.332501.733.423.731.424.385001.813.593.621.424.76集合标准误PooledSE0.040.140.110.020.36P值P-value0.2810.8590.4910.2800.673
2.4 饲粮铁含量对1~21日龄肉仔鸡十二指肠黏膜DMT1和FPN1 mRNA表达水平的影响
饲粮铁含量对1~21日龄肉仔鸡十二指肠黏膜DMT1和FPN1 mRNA表达水平的影响结果见表9。饲粮铁含量显著影响各日龄十二指肠黏膜DMT1和FPN1 mRNA表达水平(P<0.10),各日龄DMT1和FPN1 mRNA表达水平均随饲粮铁含量的增加升高而降低。
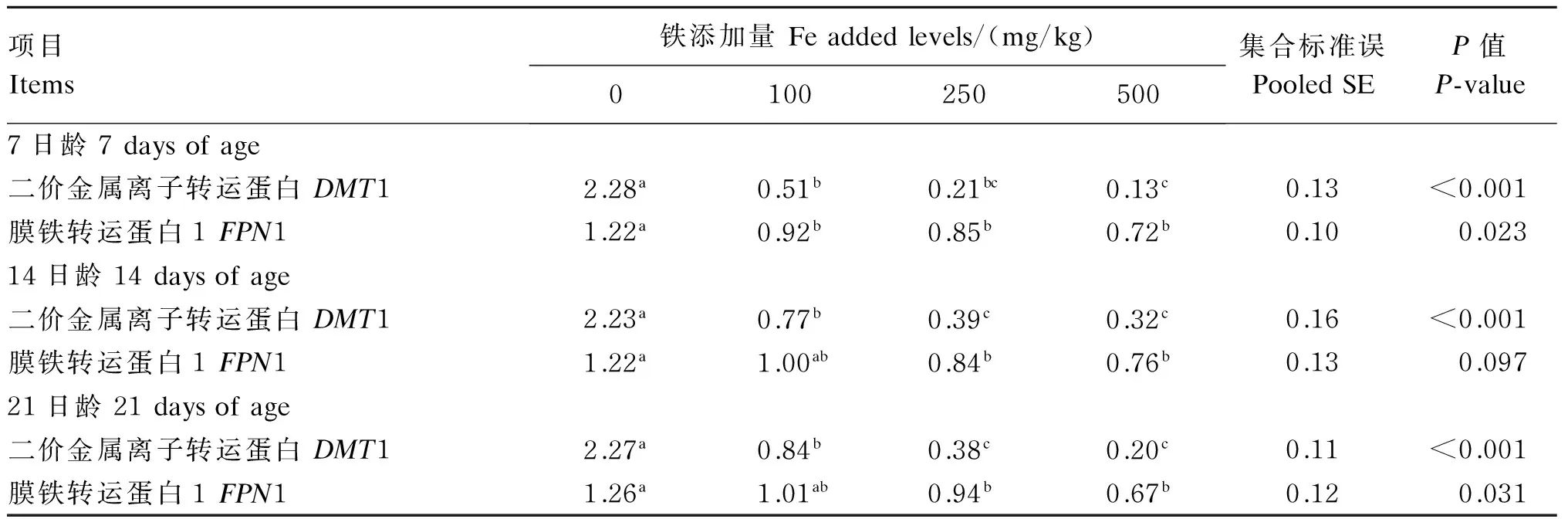
表9 饲粮铁含量对1~21日龄肉仔鸡十二指肠黏膜DMT1和FPN1 mRNA表达水平的影响Table 9 Effects of dietary Fe content on mRNA expression levels of DMT1 and FPN1 in duodenal mucosa of broilers during 1 to 21 days of age
同行数据肩标不同字母表示差异显著(P<0.10)。
Values with different letter superscripts in the same row differ significantly (P<0.10).
3 讨 论
3.1 饲粮铁含量对1~21日龄肉仔鸡生长性能的影响
Cao等[13]报道,肉仔鸡玉米-豆粕型饲粮(铁含量188 mg/kg)中添加400、600和800 mg/kg铁,1~7日龄、8~14日龄及15~21日龄阶段3个铁添加组肉仔鸡的平均日采食量和平均日增重均降低,但3个铁添加组间无显著差异。马新燕[9]报道,1~21日龄肉仔鸡采食玉米-豆粕型饲粮(铁含量67 mg/kg)中添加100 mg/kg铁的饲粮后,平均日采食量和平均日增重均未见降低。本试验中,100 mg/kg铁添加组肉仔鸡的平均日增重和平均日采食量与对照组无显著差异,但250和500 mg/kg铁添加组在1~7日龄的平均日采食量均较对照组显著降低,500 mg/kg铁添加组在1~7日龄和8~14日龄的平均日增重显著降低,说明此时机体已经呈现铁过量的负面影响;在以后日龄阶段250和500 mg/kg铁添加组表现出平均日采食量和平均日增重降低的趋势,可能与鸡只对饲粮高铁的逐渐适应有关。
3.2 饲粮铁含量对1~21日龄肉仔鸡血液指标的影响
动物体内的铁有60%~70%用于合成血红蛋白,因此血红蛋白浓度是评价动物体内铁营养状况的常用指标[9,14]。尽管动物缺铁会导致血红蛋白浓度降低[15-17],但在不缺铁饲粮中添加100~500 mg/kg铁并不增加血红蛋白浓度[5,9,18]。本试验中不加添加铁的对照组全血中血红蛋白浓度仅在7日龄显著低于100和500 mg/kg铁添加组,其他日龄阶段与各铁添加组均未表现出显著差异,可能与本试验中所用饲粮含铁量(78 mg/kg)较高、此时鸡只仅临界缺铁有关。与不加添加铁的对照组相比,铁添加组全血血红蛋白浓度和红细胞压积并未显著升高,说明体内铁已经满足正常生理需要,这也在血浆铁饱和度结果中得到了验证。作为综合反映机体铁状况的指标,血浆铁饱和度低于16%说明体内铁缺乏,超过60%则说明铁过量[19],本试验中的250 mg/kg铁添加组各日龄阶段血浆铁饱和度均在60%上下,说明机体处于临界过量状态,而500 mg/kg铁添加组各日龄阶段血浆铁饱和度均在60%以上,说明鸡只处于铁过量状态。
3.3 饲粮铁含量对1~21日龄肉仔鸡组织铁、锰、铜和锌含量的影响
肝脏、肾脏、十二指肠黏膜和胫骨灰铁含量随饲粮铁含量增加而线性增加[5,9,13,18,20-21],说明这些组织是储存铁的场所。而机体吸收的锰主要储存于含线粒体丰富的组织中,胰脏、肾脏、肝脏、心脏以及骨骼中锰沉积量随饲粮锰含量的增加而呈线性增加[22-23],说明这些组织器官是储存锰的主要场所。断奶仔猪饲粮(铁含量20 mg/kg)中添加100和500 mg/kg铁,肝脏和十二指肠铁含量线性升高而锰含量线性降低[5]。断奶犊牛饲粮(铁含量67 mg/kg)中添加750 mg/kg铁显著提高肝脏铁含量,十二指肠和心脏铁含量也有提高趋势,仅有十二指肠锰含量显著降低,肝脏和心脏锰含量并未降低[6]。关于饲粮铁含量对肉仔鸡铜、锌和锰代谢影响的研究报道很少。姜俊芳等[24]报道,在含铁231~296 mg/kg的饲粮中添加30或60 mg/kg铁,锰的表观存留率随饲粮铁含量的增加而下降。本试验中各日龄肉仔鸡肝脏、胰腺、十二指肠黏膜和胫骨灰铁含量均随饲粮铁含量的增加呈线性增加,而所测富集锰的组织中除肝脏和心脏外,胰腺、十二指肠黏膜和胫骨灰锰含量均随饲粮铁含量的增加呈线性降低,这种负相关强烈提示饲粮高铁可能通过降低肠道中DMT1的表达而降低锰的吸收量,因而导致组织锰含量降低,本试验得出的十二指肠黏膜DMT1和FPN1 mRNA表达水平随饲粮铁含量的增加而降低的结果证实了这一推论。
关于饲粮中铁对铜、锌吸收的影响,有研究报道饲粮添加铁会降低大鼠对锌[25]和铜[26]的吸收,但也有研究报道饲粮添加铁不影响肉仔鸡[20]、大鼠[25]和断奶仔猪[5]对铜的吸收。本试验所测组织中,各铁添加组7日龄胰腺锌含量均低于对照组,说明饲粮添加铁可能降低了肉仔鸡对锌的吸收,从而导致敏感组织胰腺锌含量降低;所测组织中铜含量均未受饲粮铁含量的影响,说明在本试验中饲粮添加铁对肠道内铜的吸收未产生显著影响,结果的差异可能与饲粮组成和铁添加量不同有关。
3.4 饲粮铁含量对1~21日龄肉仔鸡十二指肠黏膜DMT1和FPN1 mRNA表达水平的影响
Dupic等[27]报道,断奶大鼠饲喂无铁饲粮或添加2%羧酸铁的高铁饲粮2周,无铁饲粮组十二指肠中与铁吸收转运相关基因十二指肠细胞色素b(Dcytb)、DMT1、FPN1和转铁蛋白受体1(TfR1)的mRNA表达水平均提高,而高铁饲粮组上述基因的mRNA表达水平则明显降低。Hansen等[6]在断奶犊牛饲粮中添加750 mg/kg铁降低了十二指肠黏膜FPN1 mRNA的表达水平,DMT1 mRNA的表达水平也有降低的趋势。Hansen等[5]在21日龄断奶仔猪饲粮中添加100和500 mg/kg,2个铁添加组十二指肠黏膜DMT1 mRNA表达水平均显著降低,但2个铁添加组间无显著差异。8日龄肉仔鸡饲喂不加铁玉米-豆粕型饲粮(铁含量51 mg/kg)或添加柠檬酸铁至铁含量达到141 mg/kg饲粮6周后,铁添加组十二指肠中DMT1、FPN1和Dcytb的mRNA表达水平均显著低于不铁添加组[17]。与上述报道一致,本试验中,添加铁1周后十二指肠黏膜中DMT1和FPN1 mRNA的表达水平均持续降低,说明肉仔鸡通过调控十二指肠黏膜铁转运载体来减少过量铁的吸收,进而也减少了对二价金属锰和锌的吸收,这与本试验中所测组织中锰和锌含量降低的结果相一致。
4 结 论
① 在1~21日龄肉仔鸡玉米-豆粕型基础饲粮(含铁量78 mg/kg)中添加250和500 mg/kg铁会呈现铁临界过量和过量状态。
② 饲粮高铁会降低1~21日龄肉仔鸡胰腺、十二指肠黏膜和胫骨灰锰含量以及7日龄胰腺锌含量。
③ 饲粮高铁会降低1~21日龄肉仔鸡十二指肠黏膜中DMT1和FPN1 mRNA的表达水平。
[1] THOMSON A B R,OLATUNBOSUN D,VALVERG L S.Interrelation of intestinal transport system for manganese and iron[J].Journal of Laboratory Clinical Medicine,1971,78(4):642-655.
[2] ROSSANDER-HULTÉN L,BRUNE M,SANDSTRÖM B,et al.Competitive inhibition of iron absorption by manganese and zinc in humans[J].The American Journal of Clinical Nutrition,1991,54(1):152-156.
[3] ABBOUD S,HAILE D J.A novel mammalian iron-regulated protein involved in intracellular iron metabolism[J].Journal of Biological Chemistry,2000,275(26):19906-19912.
[4] TROADEC M B,WARD D M,LO E,et al.Induction of FPN1 transcription by MTF-1 reveals a role for ferroportin in transition metal efflux[J].Blood,2010,116(22):4657-4664.
[5] HANSEN S L,TRAKOOLJUL N,LIU H C,et al.Iron transporters are differentially regulated by dietary iron,and modifications are associated with changes in manganese metabolism in young pigs[J].The Journal of Nutrition,2009,139(8):1474-1479.
[6] HANSEN S L,ASHWELL M S,MOESER A J,et al.High dietary iron reduces transporters involved in iron and manganese metabolism and increases intestinal permeability in calves[J].Journal of Dairy Science,2010,93(2):656-665.
[7] 中华人民共和国农业部.NY/T 33—2004 鸡饲养标准[S].北京:中国农业出版社,2004:43.
[8] LI S F,LU L,HAO S F,et al.Dietary manganese modulates expression of the manganese-containing superoxide dismutase gene in chickens[J].Journal of Nutrition,2011,141(2):189-194.
[9] 马新燕.肉仔鸡对有机蛋白铁相对生物学利用率及其饲粮铁适宜水平的研究[D].硕士学位论文.北京:中国农业科学院,2012:25-32.
[10] 白世平.不同形态锰在肉仔鸡小肠中的吸收机理研究[D].博士学位论文.北京:中国农业科学院,2008:84-89.
[11] LI S F,LUO X G,LU L,et al.Bioavailability of organic manganese sources in broilers fed high dietary calcium[J].Animal Feed Science and Technology,2005,123/124:703-715.
[12] LIU S B,XIE J J,LU L,et al.Estimation of standardized phosphorus retention for inorganic phosphate sources in broilers[J].Journal of Animal Science,2013,91(8):3766-3771.
[13] CAO J,LUO X G,HENRY P R,et al.Effect of dietary iron concentration,age,and length of iron feeding on feed intake and tissue iron concentration of broiler chicks for use as a bioassay of supplemental iron sources[J].Poultry Science,1996,75(4):495-504.
[14] MILLER E R,PARSONS M J,ULLREY D E,et al.Bioavailability of iron from ferric choline citrate and a ferric copper cobalt choline citrate complex for young pigs[J].Journal of Animal Science,1981,52(4):783-787.
[15] STRUBE Y N J,BEARD J L,ROSS A C.Iron deficiency and marginal vitamin a deficiency affect growth,hematological indices and the regulation of iron metabolism genes in rats[J].The Journal of Nutrition,2002,132(12):3607-3615.
[16] RINCKER M J,HILL G M,LINK J E,et al.Effects of dietary iron supplementation on growth performance,hematological status,and whole-body mineral concentrations of nursery pigs[J].Journal of Animal Science,2004,82(11):3189-3197.
[17] TAKO E,RUTZKE M A,GLAHN R P.Using the domestic chicken (Gallusgallus) as aninvivomodel for iron bioavailability[J].Poultry Science,2010,89(2):514-521.
[18] HANSEN S L,TRAKOOLJUL N,SPEARS J W,et al.Age and dietary iron affect expression of genes involved in iron acquisition and homeostasis in young pigs[J].The Journal of Nutrition,2010,140(2):271-277.
[19] BAINTON D F,FINCH C A.The diagnosis of iron deficiency anemia[J].The American Journal of Medicine,1964,37(1):62-70.
[20] 蒲俊华.不同铁源生物学效价及其对仔鸡组织铁铜锌锰含量影响的研究[D].硕士学位论文.扬州:扬州大学,2006:11-13.
[21] 马春燕.22-42日龄肉鸡玉米-豆粕型饲粮铁适宜水平的研究[D].硕士学位论文.北京:中国农业科学院,2014:20-21.
[22] HENRY P R,AMMERMAN C B,LITTELL R C.Relative bioavailability of manganese from a manganese-methionine complex and inorganic sources for ruminants[J].Journal of Dairy Science,1992,75(12):3473-3478.
[23] BLACK J R,AMMERMAN C B,HENRY P R,et al.Biological availability of manganese sources and effects of high dietary manganese on tissue mineral composition of broiler-type chicks[J].Poultry Science,1984,63(4):1999-2006.
[24] 姜俊芳,张春善,贾春燕,等.铁与维生素A及其互作效应对肉仔鸡的生产性能、铁、铜、锰、锌表观存留率的影响[J].动物营养学报,2003,15(1):31-37.
[25] TROOST F J,BRUMMER R J M,DAINTY J R,et al.Iron supplements inhibit zinc but not copper absorptioninvivoin ileostomy subjects[J].The American Journal of Clinical Nutrition,2008,78(5):1018-1023.
[26] JOHNSON M A,MURPHY C L.Adverse effects of high dietary iron and ascorbic acid on copper status in copper-deficient and copper-adequate rats[J].The American Journal of Clinical Nutrition,1988,47(1):96-101.
[27] DUPIC F,FRUCHON S,BENSAID M,et al.Duodenal mRNA expression of iron related genes in response to iron loading and iron deficiency in four strains of mice[J].Gut,2002,51(5):648-653.
*Contributed equally
**Corresponding authors: LYU Lin, associate professor, E-mail: lulin1225@163.com; LI Sufen, professor, E-mail: lisufen64@163.com
(责任编辑 菅景颖)
Effects of High Dietary Iron on Iron Transporter Gene Expressions in Duodenal Mucosa and Tissue Microelement Contents of Broilers
ZOU Yaxue1WANG Qiuyue1*NIU Yibing1HE Ying1TANG Jiaming1LYU Lin2**ZHANG Liyang2LUO Xugang2LI Sufen1**
(1.HebeiScienceandTechnologyNormalCollege,Qinhuangdao066000,China; 2.InstituteofAnimalSciences,ChineseAcademyofAgriculturalSciences,Beijing100093,China)
This experiment was conducted to investigate the effects of dietary iron (Fe) content on tissue some important microelements such as Fe, manganese (Mn), copper (Cu) and zinc (Zn) contents and Fe transporter gene expressions in duodenal mucosa of broilers, and to explore the effects of Fe on absorption and metabolism of microelements of broilers and its mechanism. A total of 336 one-day-old Rose-308 male chicks were randomly divided into 4 groups with 6 replicates per group and 14 chicks per replicate. Chicks in control group were fed a basal diet without adding exogenous Fe (measured value of Fe content was 78 mg/kg), while those in Fe added groups were fed experimental diets added with 100, 250 and 500 mg/kg Fe (FeSO4·7H2O as Fe source) based on the basal diet (measured value of Fe content was 166, 308 and 579 mg/kg, respectively), respectively. The experiment lasted for 21 days. Chicks were slaughtered on 7, 14 and 21 days of age to analyze the contents of Fe, Mn, Cu and Zn in liver, heart, pancreas, duodenal mucosa and tibia ash and the mRNA expression levels of divalent metal transporter 1 (DMT1) and ferroportin 1 (FPN1) in duodenal mucosa. The results showed as follows: 1) chicks in the 500 mg/kg Fe added group had significantly lower average daily gain (ADG) during 1 to 7 days of age and 8 to 14 days of age than that in the other three groups (P<0.10), and chicks in the 250 and 500 mg/kg Fe added groups had significantly lower average daily feed intake (ADFI) during 1 to 7 days of age than that in the other two groups (P<0.10). 2) Plasma total iron binding capacity, hemoglobin concentration (except 7 days of age) and hematocrit in whole blood at 7, 14 and 21 days of age were not significantly affected by dietary Fe content (P>0.10). Plasma Fe content and transferrin saturation at 7, 14 and 21 days of age were significantly affected by dietary Fe content (P<0.10), and they were increased as dietary Fe content increasing. 3) The content of Fe in heart at 7 and 14 days of age and in liver, duodenal mucosa, pancreas and tibia ash at 7, 14 and 21 days of age was increased as dietary Fe content increasing, while the content of Mn in duodenal mucosa, pancreas and tibia ash was decreased. Zn content in pancreas at 7 days of age was significantly decreased by dietary added Fe (P<0.10), but the contents of Zn in pancreas at 14 and 21 days of age and Cu in each analyzed tissue at 7, 14 and 21 days of age were not affected by dietary added Fe (P>0.10). 4) The mRNA expression levels ofDMT1 andFPN1 in duodenal mucosa at 7, 14 and 21 days of age were significantly affected by dietary Fe content (P<0.10), and they were decreased as dietary Fe content increasing. These results suggest that high Fe diets may decrease the absorption of Mn and Zn through the down-regulations ofDMT1 andFPN1 gene expressions in duodenal mucosa, and then result in the lower depositions of Mn and Zn in tissues of broilers.[ChineseJournalofAnimalNutrition, 2017, 29(6):1976-1987]
iron; iron transporter gene expression; tissue microelement contents; broilers
10.3969/j.issn.1006-267x.2017.06.019
2016-11-21
国家自然科学基金项目(31272465)
邹亚学(1972—),男,河北迁安人,副教授,博士,主要从事分子生物学研究。E-mail: zouyaxue@163.com
S816
A
1006-267X(2017)06-1976-12
*同等贡献作者
**通信作者:吕 林,副研究员,硕士生导师,E-mail: lulin1225@163.com;李素芬,教授,硕士生导师,E-mail: lisufen64@163.com
