RORα高表达对二烯丙基二硫抑制人胃癌MGC803细胞上皮-间质转化的影响
2017-06-22赵晓红
夏 红,苏 坚,2,赵晓红,3,刘 芳,苏 波,凌 晖,曾 希,苏 琦
1.湖南省胃癌研究中心,湖南省高校肿瘤细胞与分子病理学重点实验室,南华大学肿瘤研究所,湖南 衡阳 421001;2.南华大学附属第二医院病理科,湖南 衡阳 421001;3.海南省妇幼保健院妇产科,海南 海口 570206
RORα高表达对二烯丙基二硫抑制人胃癌MGC803细胞上皮-间质转化的影响
夏 红1,苏 坚1,2,赵晓红1,3,刘 芳1,苏 波1,凌 晖1,曾 希1,苏 琦1
1.湖南省胃癌研究中心,湖南省高校肿瘤细胞与分子病理学重点实验室,南华大学肿瘤研究所,湖南 衡阳 421001;2.南华大学附属第二医院病理科,湖南 衡阳 421001;3.海南省妇幼保健院妇产科,海南 海口 570206
背景与目的:二烯丙基二硫(diallyl disulfide,DADS)具有抗肿瘤的作用。该研究在DADS上调人胃癌MGC803细胞RORα的基础上,观察DADS与RORα高表达对MGC803细胞上皮-间质转化(epithelialmesenchymal transformation,EMT)的影响。方法:相差显微镜观察MGC803细胞形态的影响。采用反转录聚合酶链反应(reverse transcription polymerase chain reaction,RT-PCR)、蛋白[质]印迹法(Western blot)、免疫荧光与免疫组织化学检测EMT相关分子表达。裸鼠实验检测对移植瘤生长的影响。结果:相差显微镜显示,DADS组与RORα高表达组细胞大小较MGC803细胞一致,呈圆形或椭圆形,梭形细胞少见,异型性降低。RORα高表达加DADS后,上述改变更为明显。Western blot检测结果显示,DADS组与RORα高表达组Snail蛋白表达明显下调,DADS+RORα高表达组更明显(P<0.05)。RT-PCR与Western blot检测显示,DADS与RORα高表达可明显下调Vimentin和上调E-cadherin mRNA与蛋白表达,DADS+RORα高表达组更为显著(P<0.05)。免疫荧光显示,DADS组与RORα高表达组细胞Snail与Vimentin表达较对照组明显减弱和E-cadherin表达显著增强,DADS+RORα高表达组更为显著,与Western blot结果一致。裸鼠实验显示,DADS组、RORα高表达组与RORα高表达+DADS组移植瘤较对照组生长减慢(P<0.05),并且,移植瘤体重较对照组明显降低,抑瘤率分别为15.07%、26.55%与49.27%(P<0.05)。免疫组织化学检测结果显示,DADS组、RORα高表达组与RORα高表达+DADS组较对照组的Ki-67、CD34与Vimentin表达均明显降低,E-cadherin表达明显增强。结论:RORα高表达可增强DADS体内外抑制人胃癌细胞EMT的作用。
二烯丙基二硫;RORα;人胃癌MGC803细胞;上皮-间质转化;Snail;Vimentin;E-cadherin
胃癌是最常见的恶性肿瘤之一,发生率与死亡率分别为全球第四位与第三位。据2015年最新统计,胃癌在我国的发生率与死亡率位于第二位,每年约新发67.9万和死亡49.8万人。由于患者就诊时大多已发生侵袭转移,5年生存率低于10%[1-3]。因此,研究胃癌侵袭转移机制,寻找靶点具有重要的意义。
最近,我们运用蛋白质组学技术鉴定二烯丙基二硫(diallyl disulfide,DADS)处理人胃癌MGC803细胞的差异蛋白,结果发现维甲酸相关孤核受体α(retinoid acid receptor related orphan receptor α,RORα)蛋白表达明显上调[4]。我们发现,RORα在胃癌组织中低表达,与胃癌发生和分化程度有关[5]。RORα是核受体超家族成员之一,可调节多种正常组织细胞的发育和分化、生物代谢、机体稳态维持及高级神经功能等,并且,RORα在肿瘤组织中表达下调与肿瘤发生密切相关,可能是肿瘤治疗的靶点[6-7]。DADS是大蒜中的一种脂溶性的有效成分,对多种肿瘤均有明显的抑制作用,是一种很有开发潜力的抗肿瘤药物[8]。我们前期工作表明,DADS可通过Rac1-Pak1/Rock1通路下调LIMK1、MMP-9和上调TIMP-3,抑制人胃癌细胞上皮-间质转化(epithelial-mesenchymal transformation,EMT)与迁移侵袭[9]。本研究进一步探讨DADS与RORα高表达对胃癌细胞EMT的影响及其相关机制。
1 材料和方法
1.1 细胞培养
人胃癌MGC803细胞由本实验室保存,高表达RORα人胃癌MGC803细胞由本实验室构建[10],置于含10%小牛血清的RPMI-1640培养基中,于37 ℃,CO2体积分数为5%、饱和湿度的培养箱内传代培养。取对数生长期的细胞用于实验。
1.2 主要试剂
DADS购自美国Fluka公司,RNA提取试剂盒购自美国OMEGA公司,逆转录试剂盒与BCA蛋白定量试剂盒购自美国Promega公司,RORα、E-cadherin、Vimentin与β-actin抗体购自美国Abcam公司,Snail、Ki-67与CD34抗体与ECL发光试剂盒购自美国Santa Cruz公司,新生牛血清购自杭州四季青生物工程材料有限公司。引物用Primer Premier 5.0软件设计,由生工生物工程(上海)股份有限公司合成。羊抗兔IgGHR和羊抗小鼠IgG-HRP购自江苏凯基生物技术股份有限公司,羊抗小鼠IgG(H+L)购自美国Protech公司,DAPI和正常山羊血清购自武汉博士德生物工程有限公司,MaxVisionTM试剂盒购自福州迈新生物技术开发有限公司。
1.3 相差显微镜观察
人胃癌MGC803细胞与RORα高表达MGC803细胞以及经DADS处理的两种细胞培养24 h后,置于倒置相差显微镜下观察细胞形态学变化。
1.4 采用反转录聚合酶链反应(reverse transcription polymerase chain reaction,RTPCR)
采用总RNA试剂盒提取细胞总RNA,在AMV酶作用下逆转录合成cDNA。设计并合成PCR引物序列。Vimentin顺义链为5’-ACACCCTGCAATCTTTCAGACA-3’,反义链为5’-AGAAATCCTGCTCTCCTCGCCT-3’,产物长度635 bp;E-cadherin顺义链为5’-CTCCCAATACATCTCCCTTCA C-3’,反义链为5’-CGCCTCCTTCTTCATCATAGTAA-3’,产物长度423 bp;β-actin顺义链为5’-TCTACA AT G A G C T G C G T G T G G-3’,反义链:5’-GGAACCGCTCATTGCCAATG-3’,产物长度498 bp。PCR反应条件:94 ℃,5 min;94 ℃,40 s,各基因Tm,45 s,72 ℃ 80 s,28个循环;72 ℃ 10 min。5 μL的PCR产物经1%的琼脂糖电泳,嗅化乙啶染色,通过IS1000图像分析软件读取条带灰度值,相对值以目的基因与β-actin灰度值之比表示。
1.5 蛋白[质]印迹法(Western blot)检测
收集细胞,提取细胞总蛋白,BCA法测定蛋白浓度,每组取等量样本进行SDS-PAGE凝胶电泳,电泳后转膜,封闭1 h,加一抗,4 ℃过夜,TBST洗膜,加二抗温育1 h,洗膜,ECL发光,X片曝光、显影、定影。
1.6 细胞免疫荧光实验
在6孔板中滴加培养基,将消毒的盖玻片放入6孔板。将对数生长期的MGC803细胞制成悬液,每孔接种5×105个细胞,细胞贴壁融合至80%时,取出盖玻片,PBS洗3次,每次5 min,采用4%多聚甲醛固定,常温静置15 min,PBS洗3次,每次5 min;将0.5%Triton覆盖细胞后,常温静置30 min,PBS洗3次,每次5 min,吸干PBS液;山羊血清封闭,于37 ℃温育1 h;吸净封闭液,加入一抗,湿盒内4℃温育过夜;第2天移置37 ℃下复温1 h,PBS洗3次,每次5 min;暗室中加FITC的二抗,37 ℃下避光湿盒中温育1 h,PBS洗3次,每次5 min。染核:0.4 μg/μL DAPI覆盖细胞,避光静置2~5 min,PBS洗3次,每次5 min,甘油封片在荧光显微镜下观察。
1.7 裸鼠成瘤实验
裸鼠购自北京维通利华实验动物技术有限公司,为4周龄雄性,分为MGC803细胞组、MGC803+DADS组、RORα高表达组和RORα高表达+DADS组,每组5只。将处于对数生长期的细胞1×107个/mL,分别取0.2 mL细胞悬液接种于各组裸鼠的腋下。观察裸鼠进食、饮水、精神及活动等情况。每周测量移植瘤大小,肿瘤体积:V=ab2/2,以裸鼠移植瘤体积的平均值,绘制生长曲线。第10周麻醉处死裸鼠,完整剥离肿瘤,称移植瘤重量,瘤重抑瘤率=(1-实验组瘤重/对照组瘤重)×100%。移植瘤组织固定于4%的中性甲醛溶液中。
1.8 免疫组织化学检测
采用MaxVisionTM法,分别滴加一抗室温60 min,4 ℃过夜,用PBS洗3次,每次3 min,滴加即用型MaxVisionTM试剂,室温下15 min,PBS洗3次,每次5 min。DAB显色,自来水冲洗,苏木素复染,脱水、透明、封固、镜检。
1.9 统计学处理
2 结 果
2.1 相差显微镜观察
采用相差显微镜对细胞进行观察。镜下可见MGC803细胞大小不一,大部分呈长梭形,纤维母细胞样,细胞膜可见突起,核质比例增大,异型性明显。30 mg/L DADS组细胞大小较一致,大部分呈圆形或椭圆形,梭形细胞少见,细胞膜突起减少,核质比值下降,异型性降低。RORα高表达细胞大小较一致,呈圆形或椭圆形,细胞膜突起少见,核质比例减少,异型性显著降低。RORα高表达加DADS后,上述改变更为明显。表明DADS与RORα高表达可从形态上抑制MGC803细胞EMT,RORα高表达可增强DADS的抑制作用(图1)。

图1 RORα高表达对MGC803细胞形态的影响Fig. 1 The morphological effect of overexpression of RORα in MGC803 cells
2.2 DADS与RORα高表达抑制MGC803细胞EMT
Western blot检测结果显示,DADS与RORα高表达可明显下调MGC803细胞Snail蛋白表达(P<0.05)。并且,RT-PCR与Western blot检测结果显示,DADS与RORα高表达可下调Vimentin和上调E-cadherin mRNA与蛋白表达(P<0.05)。上述改变RORα/MGC803+DADS作用更为明显(P<0.05)。表明DADS与RORα高表达可通过下调Snail与Vimentin和上调E-cadherin抑制MGC803细胞EMT,RORα高表达可增强DADS的抑制作用(图2~4)。
2.3 细胞免疫荧光检测DADS与RORα高表达对EMT相关蛋白表达的影响
免疫荧光检测结果显示,Snail蛋白定位细胞核,Vimentin与E-cadherin蛋白主要定位于细胞质。DADS与RORα高表达组细胞Snail与Vimentin阳性表达较对照组明显减弱,而E-cadherin阳性表达显著增强,与Western blot检测结果一致(图5)。
2.4 DADS与RORα高表达对裸鼠移植瘤生长的影响
生长曲线图显示,随着时间的延长,移植瘤体积逐渐增大,但是,DADS较对照组移植瘤生长明显减慢;而RORα高表达组生长速度减慢程度更加显著,特别是DADS处理后,生长速度减慢最为显著(P<0.05,图6)。图7显示,DADS组、RORα高表达组与RORα高表达+DADS移植瘤平均体重分别为4.51±0.29、3.90±0.33和2.80±0.27,较对照组的5.31±0.37明显降低,抑瘤率分别为15.07%、26.55%和49.27%,差异有统计学意义(P<0.05)。该结果表明DADS与RORα可在体内抑制MGC803细胞移植瘤形成,RORα高表达具有增强DADS的作用。

图2 DADS与RORα高表达对Snail蛋白表达的影响Fig. 2 The effect of Snail expression by DADS and overexpression of RORα
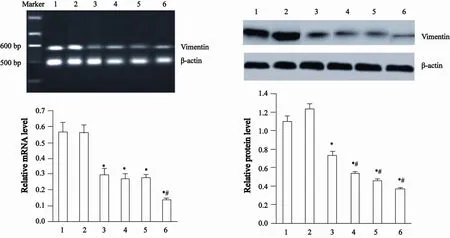
图3 DADS与RORα高表达对Vimentin表达的影响Fig. 3 The e ff ect of Vimentin expression by DADS and overexpression of RORα

图4 DADS与RORα高表达对E-cadherin表达的影响Fig. 4 The e ff ect of E-cadherin expression by DADS and overexpression of RORα
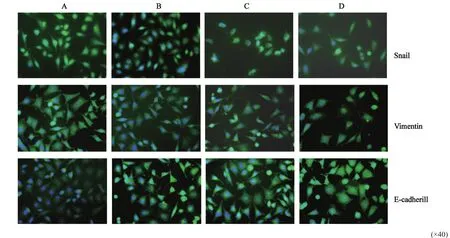
图5 DADS与RORα高表达对EMT相关蛋白表达的影响Fig. 5 The e ff ect of EMT associated proteins expression by DADS and overexpression of RORα

图6 DADS与RORα高表达对裸鼠移植瘤生长的影响Fig. 6 The effect of growth in mouse xenograft models by DADS and overexpression of RORα
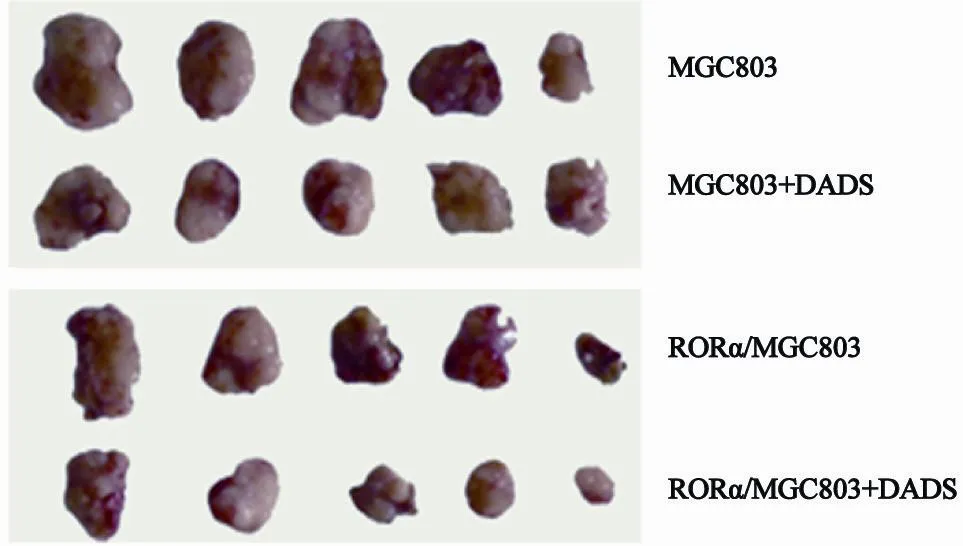
图7 DADS与RORα高表达抑制裸鼠移植瘤形成的作用Fig. 7 DADS and overexpression of RORα inhibit the formation in transplantation tumor
2.5 RORα高表达对裸鼠移植瘤组织EMT相关蛋白表达的影响
免疫组织化学检测结果显示,DADS组、RORα高表达组与RORα高表达+DADS组移植瘤组织的Ki-67、CD34与Vimentin阳性表达分别较对照组均明显减弱,而E-cadherin阳性表达明显增强(图8)。

图8 裸鼠移植瘤组织EMT相关蛋白的表达Fig. 8 The expression of EMT associated proteins in transplantation tumor tissue
3 讨 论
研究表明,RORα在胃癌、结肠癌、食管癌、胰腺癌、肝癌、乳腺癌、子宫颈癌、卵巢癌、前列腺癌、膀胱癌、头颈部癌及白血病等多种肿瘤中表达下调[5-7]。恢复RORα表达可体内外抑制乳腺癌细胞增殖与侵袭,提示RORα可能是肿瘤治疗靶点[7]。研究发现,RORα在肝癌组织表达明显下调,与血清AFP、病理分级、肿瘤复发、血管侵袭和预后密切相关。并且,RORα高表达可减少肝癌细胞需氧糖酵解和下调生物合成途径,上调p21,抑制PDK2表达和磷酸化[11-12]。但是,RORα抑制肿瘤细胞迁移侵袭与转移是否与EMT有关尚不清楚。
肿瘤细胞获得迁移和侵袭能力是肿瘤转移起始的重要步骤,而EMT是肿瘤细胞获得迁移和侵袭能力的关键。上皮源性肿瘤细胞发生EMT后,除形态发生改变外,具有较高的迁移与侵袭、抗凋亡和降解细胞外基质的能力,上皮标志物E-cadherin、ZO-1等表达下调,间质标志物Vimentin、α-SMA、N-cadherin等上调以及Snail、Slug和Twist等转录因子活性增强。此时,肿瘤细胞间的连接变得疏松,细胞骨架蛋白发生重组,黏附能力下降,迁移能力增强,更易于离开原有位置发生侵袭与转移。因此,肿瘤细胞出现EMT改变是启动肿瘤侵袭与转移的关键所在,阻止EMT发生已经成为抑制恶性肿瘤转移的新的治疗策略[13-14]。
本研究在在DADS上调人胃癌MGC803细胞RORα的基础上,探讨DADS与RORα高表达对胃癌细胞EMT的影响。相差显微镜显示,DADS组与RORα高表达组细胞大小较MGC803细胞一致,呈圆形或椭圆形,梭形细胞少见,异型性降低,DADS处理RORα高表达细胞后,上述改变更为明显。基于Snail、E-cadherin与Vimentin是肿瘤EMT与进展的关键因子[15-16],Ki-67是检测肿瘤增殖能力的重要指标[17],CD34是血管形成的标志[18]。RT-PCR与Western blot检测结果显示,DADS与RORα高表达可明显下调Snail与Vimentin和上调E-cadherin表达,DADS+RORα高表达组更为显著。免疫荧光检测结果与Western blot检测结果一致。裸鼠实验显示,DADS组、RORα高表达组与RORα高表达+DADS组移植瘤较对照组生长减慢,移植瘤体重明显降低,抑瘤率分别为15.07%、26.55%与49.27%。免疫组织化学检测结果显示,DADS组、RORα高表达组与RORα高表达+DADS组较对照组的Ki-67、CD34与Vimentin阳性表达均明显降低和E-cadherin阳性表达明显增强。上述证明DADS与RORα高表达通过下调Snail与Vimentin和上调E-cadherin表达可体内外抑制人胃癌细胞EMT,RORα高表达可增强DADS的作用。
研究表明,RORα可通过Wnt/β-catenin、Wnt5a/PKC、Hypoxia/Angiogenesis、NF-κB和p53等多种途径调控肿瘤细胞增殖与迁移侵袭[7]。Lee等[19]的研究结果显示,RORα可通过丝氨酸35残基磷酸化,竞争结合β-catenin,抑制Wnt/β-catenin靶基因cyclin D1、c-myc和Axin,从而调控细胞增殖与肿瘤进展。此外,RORα可依赖PGE2/PKCα途径磷酸化减弱结肠癌细胞Wnt靶基因表达[20]。近来,我们发现,RORα高表达可抑制人胃癌细胞Wnt/β-catenin通路靶基因表达和增殖与迁移侵袭[7,21]。然而,DADS上调RORα抑制人胃癌细胞EMT是否通过Wnt/β-catenin通路调控的分子机制尚待深入研究。
[1] TORRE L A, BRAY F, SIEGEL R L, et al. Global cancer statistics, 2012[J]. CA Cancer J Clin, 2015, 65(2): 87-108.
[2] ORDITURA M, GALIZIA G, SFORZA V, et al. Treatment of gastric cancer[J]. World J Gastroenterol, 2014, 20(7): 1635-1649.
[3] CHEN W, ZHENG R, BAADE P D, et al. Cancer statistics in China, 2015 [J]. CA Cancer J Clin, 2016, 66(2): 115-132.
[4] SU B, SU J, HE H, et al. Identification of potential targets for diallyl disulfide in human gastric cancer MGC-803 cells using proteomics approaches[J]. Oncol Rep, 2015, 33(5): 2484-2494.
[5] 石 莺, 黄建军, 苏 坚, 等. RORα蛋白在胃癌中的表达及临床病理意义[J]. 临床与实验病理学杂志, 2012, 28(3): 270-273.
[6] 赵晓红, 苏 琦. 维甲酸相关孤核受体α与Wnt信号途径及肿瘤的关系[J]. 国际病理科学与临床杂志, 2011, 31(3): 234-237.
[7] DU J, XU R. RORα, a potential tumor suppressor and therapeutic target of breast cancer[J]. Int J Mol Sci, 2012, 13(12):15755-17566.
[8] YI L, SU Q. Molecular mechanisms for the anti-cancer effects of diallyl disulfide[J]. Food Chem Toxicol, 2013, 57:362-370.
[9] SU B, SU J, ZENG Y, et al. Diallyl disulfide suppresses epithelial-mesenchymal transition, invasion and proliferation by downregulation of LIMK1 in gastric cancer[J]. Oncotarget, 2016, 7(9): 10498-10512.
[10] 赵晓红, 向姝霖, 刘 芳, 等. RORα高表达对人胃癌MGC803细胞增殖与迁移侵袭的影响[J]. 肿瘤防治研究, 2016, 43(11): 926-932.
[11] FU R D, QIU C H, CHEN H A, et al. Retinoic acid receptorrelated receptor alpha (RORalpha) is a prognostic marker for hepatocellular carcinoma[J]. Tumour Biol, 2014, 35(8): 7603-7610.
[12] BYUN J K, CHOI Y K, KANG Y N, et al. Retinoic acidrelated orphan receptor alpha reprograms glucose metabolism in glutamine-deficient hepatoma cells[J]. Hepatology, 2015, 61(3): 953-964.
[13] GOMES L R, TERRA L F, SOGAYAR M C, et al. Epithelialmesenchymal transition: implications in cancer progression and metastasis[J]. Curr Pharm Biotechnol, 2011, 12(11): 1881-1890.
[14] MENG F, WU G. The rejuvenated scenario of epithelial-mesenchymal transition (EMT) and cancer metastasis[J]. Cancer Metastasis Rev, 2012, 31(3-4): 455-467.
[15] ABOUHASHEM N S, IBRAHIM D A, MOHAMED A M. Prognostic implications of epithelial to mesenchymal transitionrelated proteins (E-cadherin, Snail) and hypoxia inducible factor 1α in endometrioid endometrial carcinoma[J]. Ann Diagn Pathol, 2016, 22: 1-11.
[16] LAZAROVA D L, BORDONARO M. Vimentin, colon cancer progression and resistance to butyrate and other HDACis[J]. J Cell Mol Med, 2016, 20(6): 989-993.
[17] LEE W S, PARK Y L, KIM N, et al. Myeloid cell leukemia-1 regulates the cell growth and predicts prognosis in gastric cancer[J]. Int J Oncol, 2015, 46(5): 2154-2162.
[18] LEE O, CHOI M R, CHRISTOV K, et al. Progesterone receptor antagonism inhibits progestogen-related carcinogenesis and suppresses tumor cell proliferation[J]. Cancer Lett, 2016, 376(2): 310-317.
[19] LEE J M, KIM I S, KIM H, et al. RORalpha attenuates Wnt/beta-catenin signaling by PKCalpha-dependent phosphorylation in colon cancer[J]. Mol Cell, 2010, 37(2): 183-195.
[20] SHIN D, KIM I S, LEE J M, et al. The hidden switches underlying RORα-mediated circuits that critically regulate uncontrolled cell proliferation[J]. J Mol Cell Biol, 2014, 6(4): 338-348.
[21] 苏 坚, 赵晓红, 刘 芳, 等. RORα高表达抑制人胃癌细胞Wnt/β-catenin通路靶基因[J]. 中国细胞生物学学报, 2016, 38(11): 1358-1365.
Effect of RORα overexpression on inhibition of epithelial-mesenchymal transformation by DADS in human gastric MGC803 cells
XIA Hong1, SU Jian1,2, ZHAO Xiaohong1,3, LIU Fang1, SU Bo1, LING Hui1, ZHEN Xi1, SU Qi1
(1. Center for Gastric Cancer Research of Hunan Province, Key Laboratory of Cancer Cellular and Molecular Pathology of Hunan Provincial University, Cancer Research Institute, University of South China, Hengyang 421001, Hunan Province, China; 2. Department of Pathology, the Second Affiliated Hospital, University of South China, Hengyang 421001, Hunan Province, China; 3. Department of Gynaecology and Obstetrics, Hainan Maternal and Child Health Hospital, Haikou 570206, Hainan Province, China)
SU Qi E-mail: suqi1945@163.com
Background and purpose: Diallyl disulfide (DADS) could inhibit the growth of cancer cells. This study aimed to investigate the effect of DADS and overexpression of RORα on EMT in human gastric cancerMGC803 cells with upregulation of RORα by DADS. Methods: The morphological effect on MGC803 cells was observed by phase-contrast microscope. The correlative molecules with EMT were detected by RT-PCR, Western blot, immunof l uorescence and immunohistochemistry. The inf l uence on xenograft tumor growth in nude mice was observed in MGC803 cells. Results: Phase-contrast microscope showed that MGC803 cells were of identical size, round or oval with decreased spindle cells and lower level of heteromorphism in DADS group and RORα/MGC803 group. The above-mentioned alterations were more obvious in RORα/MGC803+DADS group. Western blot exhibited obviously the downregulation of Snail protein in DADS group and RORα/MGC803 group (P<0.05). RT-PCR and Western blot disclosed that the expression of Vimentin was downregulated notably and E-cadherin was upregulated in DADS group, RORα/MGC803 group, and RORα/MGC803+DADS group more obviously (P<0.05). Immunofluorescence revealed that the positive expression of Snail and Vimentin protein was attenuate, while E-cadherin was strengthened in DADS group, RORα/MGC803 group and RORα/MGC803+DADS group compared with MGC803 cells. Moreover, the xenograft tumor growth was markedly decreased, and body weight of transplanted tumor was visibly reduced with the inhibition ratio of 15.07%, 26.55 % and 49.27%, respectively (P<0.05). The positive expression of Ki-67, CD34 and Vimentin were obviously decreased, while the positive expression of E-cadherin was increased. Conclusion: Overexpression of RORα can remarkably enhance inhibition of EMT in MGC803 cells by DADS in vivo and in vivo.
Diallyl disulfide; RORα; Human gastric cancer MGC803 cells; Epithelial-mesenchymal transformation; Snail; Vimentin; E-cadherin
10.19401/j.cnki.1007-3639.2017.05.007
R735.2
A
1007-3639(2017)05-0359-09
2016-12-30
2017-02-10)
国家自然科学基金(81374013)。
苏 琦 E-mail:suqi1945@163.com
