耦合Pre-Bötzinger复合体中神经元的反相簇放电模式及同步研究*
2017-06-07刘晶曹秦禹王子剑赵勇段利霞
刘晶曹秦禹王子剑赵勇段利霞†
(1.北方工业大学理学院,北京 100144)(2.河南理工大学数学与信息科学学院,焦作 454000)
耦合Pre-Bötzinger复合体中神经元的反相簇放电模式及同步研究*
刘晶1曹秦禹1王子剑1赵勇2段利霞1†
(1.北方工业大学理学院,北京 100144)(2.河南理工大学数学与信息科学学院,焦作 454000)
本文以pre-Bötzinger复合体中两个耦合兴奋性神经元为研究对象,分别研究了耦合神经元反相簇放电类型及其同步转迁.当钠电导参数在一定范围内变化时,耦合神经元分别表现为“sup-Hopf/fold cycle”型和“fold/fold cycle”型反相簇放电.通过计算耦合神经元的簇放电内的峰相位差,进而判定反相簇放电中的峰同步行为,并且研究了反相放电的类型与同步的关系.
簇放电,相位差,反相同步,快慢分析,前包钦复合体
引言
簇放电是神经系统中对信息处理具有重要作用的神经放电模式,近年来簇放电的类型及其同步行为引起研究者们的广泛关注.Ivanchenko等给出簇放电映射神经元的混沌相位同步并发现相位同步并非只与耦合强度相关[1-2].Rulkov给出了两个耦合神经元簇同步的产生机制,并研究了电耦合神经元的同相簇同步和反相簇同步[3].研究者对于同步的产生机制做了大量而有意的工作[4-8].
目前,研究者大多认为位于哺乳动物延髓外侧区的pre-Bötzinger复合体与呼吸节律的产生密切相关.呼吸运动之所以能自主地有节奏地进行,是因为pre-Bötzinger复合体中的一些细胞群,其中包括一些吸气神经元,它们可自主地按一定的频率发出冲动,即神经元放电活动.这些神经元集群能够与其它呼吸组织互相联系,共同调节和控制呼吸运动.在pre-Bötzinger复合体中,兴奋性突触耦合可以促成神经元的同步活动[9-10].
Best等人于2005年研究了pre-Bötzinger复合体兴奋性中间神经元模型,发现了两种不同性质的同步放电模式,并将其定义为对称与非对称的簇放电和对称与非对称的峰放电[10].Caiteri和Rubin研究了在pre-Bötzinger复合体中神经元的动力学本质和网络拓扑结构的交互作用对神经元簇放电的影响,他们研究了单个神经元的放电模式及其在网络中的位置对神经元同步的影响,并进一步比较了不同的网络拓扑结构对神经元的同步的影响[9].Rubin和Terman等利用简化的pre-Bötzinger复合体神经元模型,研究了两个神经元同步的条件,得到了稳定的同步周期解,并给出了稳定性的充分条件;他们还进一步给出了破坏同步的条件[11].Dunmyre和Rubin利用简化的三细胞网络模型,研究了神经元内在动力学性质对神经元的簇同步的影响,得到了同步解存在的充分必要条件;此外,他们还给出了三神经元网络的数值结果,其中,三个神经元分别是静息态神经元、峰放电神经元和一个添加的神经元;结果表明,当加入静息态神经元时,网络可以达到最佳同步放电模式;而加入簇放电的神经元时反而会破坏神经元的簇同步[12].对 pre-Bötzinger复合体中耦合神经元的簇同步也有广泛的研究[13-14].
同一参数条件下,pre-Bötzinger复合体神经元会产生两种不同的簇放电模式:同相簇放电和反相簇放电[10],这里反相簇同步是指簇中的峰是反相同步的.对于这类反相簇同步研究及其与簇放电模式之间的关系,目前的研究较少.
本文的结构如下:第一节给出了Butera模型的数学表示;第二节给出了反相簇放电的类型及快慢动力学分析,并计算了不同类型簇放电中峰放电的相位差;最后是本文的结论.
1 模型描述
Butera等人在1999年[9]提出的pre-Bötzinger复合体中兴奋性神经元模型描述如下:
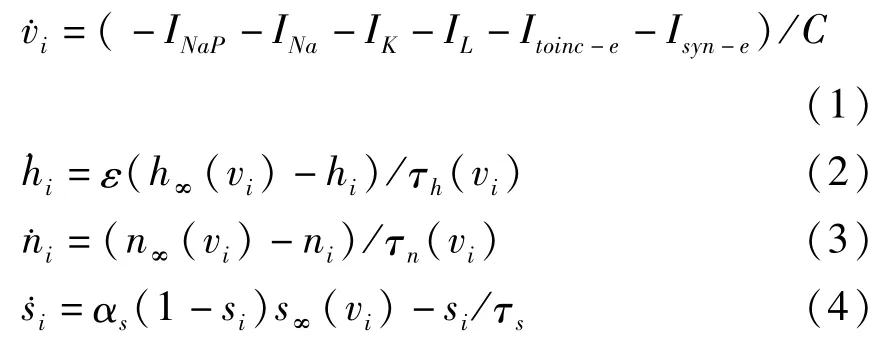
其中,i∈{1,2}.C是膜电容,v是膜电位,INaP,INa,IK和IL分别代表持续钠电流、钠电流、钾电流和漏电流.Itonic-e表示细胞受到兴奋性刺激所产生的电流,Isyn-e表示耦合神经元网络连接而产生的突触电流.各离子电流动力学模型描述为:INaP=gNaPmp,∞(vi)hi(vi-ENa),,IL=gL(vi-EL),Isyn-e= gsyn-esj(vi-Esyn-e),Itonic-e=gtonic-e(vi-Esyn-e),其中i,j∈{1,2}且i≠j.在本文模型中,对于x∈{mp,m,h,n,s},函数x∞(v)可表示为x∞(v)={1+exp[(v-θx)/σx]}-1.对于x∈{h,n},函数τx(v)可表示为τx(v)=τx/cos h[(v-θx)/2σx].其中所用到的参数的值和单位见表1.
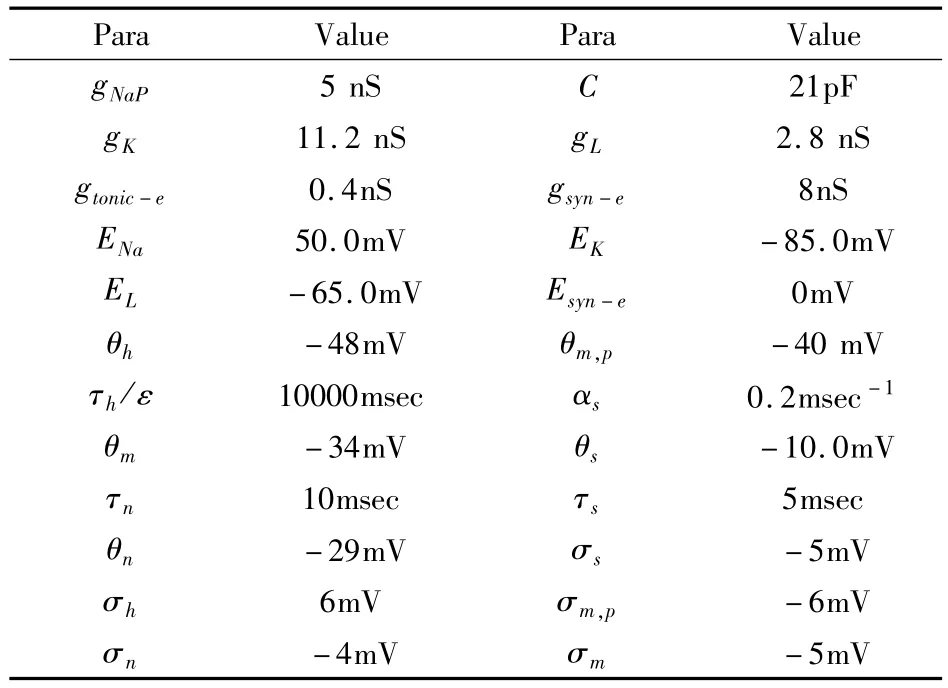
表1 模型中的参数值Table 1 Parameters of the model
2 耦合神经元的簇放电模式及簇放电同步
耦合pre-Bötzinger复合体神经元在同一参数集下,但初始条件不同时会表现出两类不同的簇放电模式,即同相簇和反相簇[15-16].这里,同相簇是指簇中的峰是同相的,反相簇是指簇中的峰是反相的.当耦合神经元表现为同相簇放电模式时,两个膜电位v1和v2随时间t的变化完全一致,因此,本文只考虑反相簇的情形.
2.1 耦合神经元的簇放电模式
当钠离子电导gNa变化时,耦合pre-Bötzinger复合体神经元会产生不同类型的反相簇放电模式,如图1所示.根据Izhikevich[17]对簇放电的基于快慢动力学的分类方法可知,当gNa的值逐渐增大时,反相簇放电的模式逐渐发生变化.下面我们具体分析反相簇放电的类型及相应的同步情况.
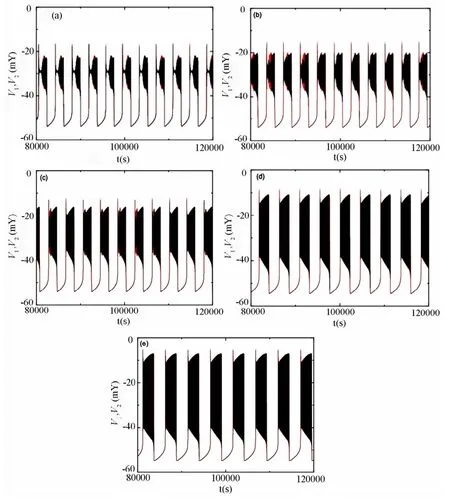
图1 耦合神经元反相簇放电模式(a)gNa=2.2nS;(b)gNa=2.6nS;(c)gNa=3.5nS;(d)gNa=5.5nS;(e)gNa=8.0nSFig.1 Modes of anti-phase bursting for two-coupled neurons(a)gNa=2.2nS;(b)gNa=2.6nS;(c)gNa=3.5nS;(d)gNa=5.5nS;(e)gNa=8.0nS
2.2 耦合神经元的簇放电同步
在pre-Bötzinger复合体中,兴奋性耦合神经元的簇放电同步模式可表现为同相同步和反相同步.鉴于同相簇放电的膜电位完全一致,这里只研究反相簇放电的形式,如图2所示.图2(a)为gNa=5.5nS时的反相簇放电,图2(b)为图2(a)的局部放大,即详细描述反相簇放电中峰与峰的差别,其中v1,v2反相同步.
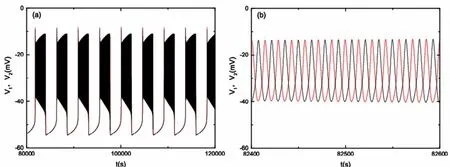
图2 (a)膜电位v1和v2关于时间t的变化;(b)图(a)的局部放大,其中红线代表v1,黑线代表v2Fig.2 (a)Time history of v1(red)and v2(black);(b)Local enlargement of a part in Fig.2(a)
为了研究耦合神经元的反相簇放电中的峰同步,我们引入峰相位的定义[5].神经元在t时刻的峰相位定义如下:

其中,K是总时间内放电产生峰的总量.
根据峰相位定义,我们可以计算两个神经元在时间t的峰相位φ(t)1和φ(t)2,进而得到他们的峰相位差Δφ(t).当同相峰放电同步时,峰相位差总是Δφ(t)=0.这里,我们主要研究反相簇放电同步,本文的数据结果都是在去掉8×106个时间单位的暂态之后得到的.

图3 当gNa=2.2nS时,耦合神经元簇放电的快慢分析及相位差(a)“sup-Hopf/fold cycle”型簇放电的快慢分岔分析;(b)耦合神经元簇放电中峰与峰之间的相位差Fig.3 Fast-slow decomposition of the two-coupled neurons and phase difference when gNa=2.2nS.(a)Fast-slow decomposition of“sup-Hopf/fold cycle”bursting;(b)Phase difference between inter-spiking.
当gNa=2.2nS时,对应的快子系统(1),(3)和(4)关于慢变量h(h1=h2)的分岔如图3(a)所示.在(h,V1)平面上,快子系统的平衡点由三条“S”型曲线组成,其中实线部分代表稳定的平衡点,虚线部分代表不稳定的平衡点.“S”型曲线上的分岔点(F1,F2和F)代表平衡点的鞍结分岔.在“S”型曲线的上部,当慢变量h增加时,不稳定焦点经由超临界Hopf(supH)分岔变为稳定焦点,同时极限环产生.系统轨线的静息态经由平衡点的鞍结分岔(F1)转迁到放电态,放电态又经由极限环的鞍结(LP)分岔转变为静息态,从而形成“sup-Hopf/fold cycle”型反相簇放电.图3(b)为对应的“sup-Hopf/fold cycle”型反相簇放电中峰与峰之间的相位差.图中的红色图框的放大图为其中一个簇放电中的两个神经元的峰相位差,由相位差的变化趋势可知,当gNa=2.2nS时,簇放电未达到反相同步.
当gNa=2.6nS时,超临界Hopf(supH)分岔向右移动,使得稳定焦点的吸引域变小.轨线的振荡在完全被吸引到稳定焦点前便经过了超临界Hopf(supH)分岔点,故而轨线被快速吸引到由超临界Hopf分岔产生的稳定极限环上,如图4(a)所示.因而,“sup-Hopf/fold cycle”型反相簇放电转变为另一类型的放电模式.但这一簇放电还保留了“sup-Hopf/fold cycle”型反相簇放电的某些特征,因此,我们称之为过渡型簇放电.这时两个神经元的簇放电中的峰之间相位差如图4(b)所示.如图中的红色放大框图所示,在一个簇放电开始和结束的时候,峰放电的相位差震荡的幅度比较大,并且在开始时震荡的时间比较长(如图4(b)中红色放大图).处于过渡时期的反相簇放电没有达到完全峰同步.

图4 当gNa=2.6nS时,耦合神经元簇放电的快慢分析及峰相位差(a)过渡时期的簇放电的快慢分岔分析;(b)过渡时期耦合神经元簇放电中峰与峰之间的相位差Fig.4 Fast-slow decomposition of the two-coupled neurons and phase difference when gNa=2.6nS.(a)Fast-slow decomposition of transition bursting;(b)Phase difference between inter-spiking
当gNa逐渐增大时,分别取值为3.5nS,5.5nS,8.0nS,其快慢分岔具有相同的结构,且各分岔点的相对位置相同(如图5(a)、(c)和(e)所示).神经元的静息态经由平衡点的鞍结(F1)分岔转变为放电态,放电态又经由极限环的鞍结(LP)分岔转变为静息态.因此,这几种簇放电具有相同的模式,称为“fold/fold cycle”型反相簇放电.
下面进一步研究这些簇放电同步情况.这里,时间间隔描述的是由8×104到2×105的多个簇放电的相位差.当gNa=3.5nS时,簇放电中峰与峰之间的相位差如图5(b)所示.在一个周期簇放电中,两个神经元的峰放电之间相位差如图5(b)中的红色放大框图所示.在一个簇放电开始和结束的时候,峰放电的相位差震荡的幅度比较大,这表示在这两个阶段簇放电未达到反相同步,而在簇放电中间阶段的峰相位差的震荡幅度非常小,所以簇放电达到了反相同步.其原因是在簇放电开始和结束时,快子系统分别经历了两个分岔,即平衡点的鞍结分岔和极限环的鞍结分岔.
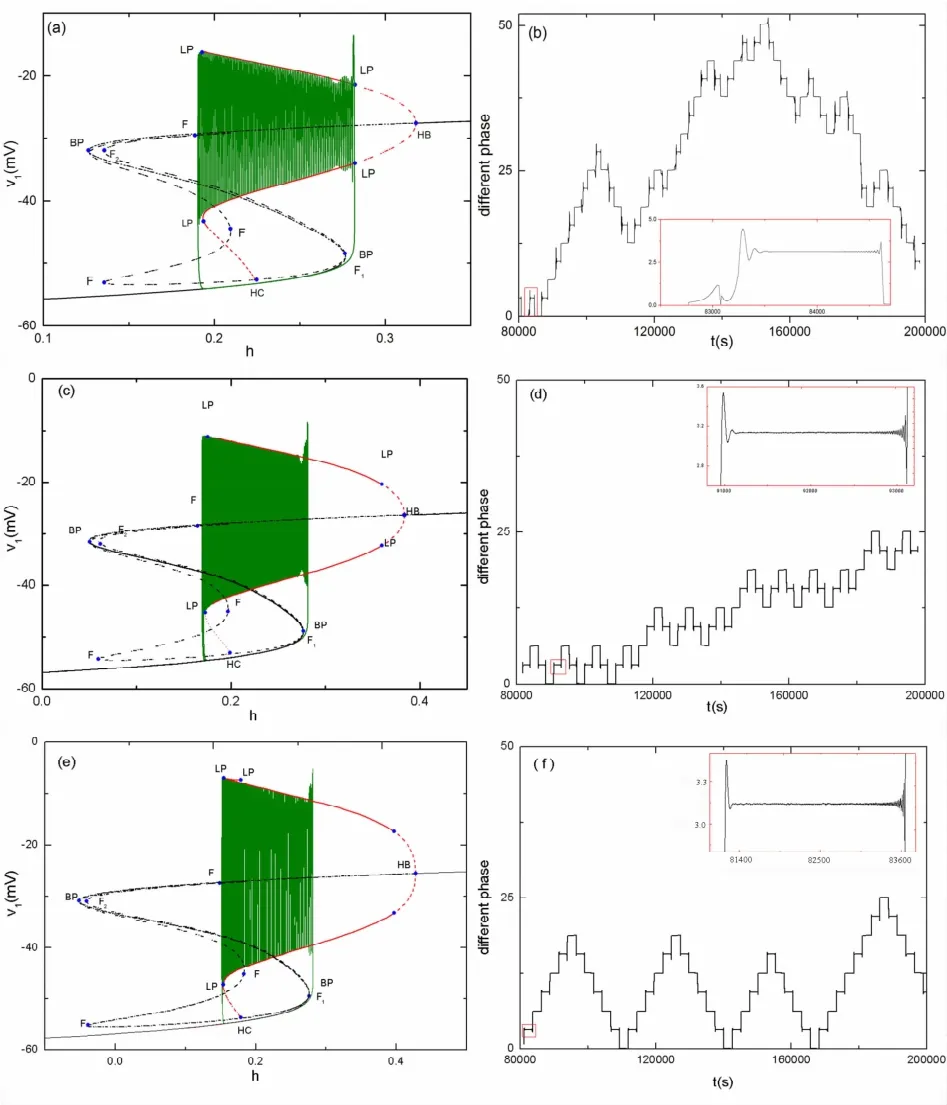
图5 耦合神经元簇放电的快慢分析及峰相位差,“fold/fold cycle”型反相簇放电.(a)当gNa=3.5nS时的快慢分岔分析;(b)gNa=3.5nS时簇放电的相位差;(c)当gNa=5.5nS时的快慢分岔分析;(d)gNa=5.5nS时簇放电的相位差;(e)当gNa=8nS时的快慢分岔分析;(f)gNa=8nS时簇放电的相位差.Fig.5 Fast-slow decomposition of the two-coupled neurons and phase difference for“fold/fold cycle”bursting.(a)Fast-slow decomposition when gNa=3.5nS;(b)Phase difference when gNa=3.5nS;(c)Fast-slow decomposition when gNa=5.5nS;(d)Phase difference when gNa=5.5nS;(e)Fast-slow decomposition when gNa=8nS;(f)Phase difference when gNa=8nS.
当gNa逐渐增大为5.5nS和8.0nS时,电相位差分别如图5(d)、5(f)所示.虽然簇放电类型是相同的,但如图5(d)和图5(f)中的红色放大图的峰放电相位差所示,簇放电中峰与峰之间的同步区域增加,簇放电开始和结束时的峰相位差的震荡幅度减小.
3 结论
本文利用快慢变量分岔分析和相位差等方法,研究了pre-Bötzinger复合体中兴奋性神经元节律性反相簇放电及簇中峰同步的情形.结果表明,当钠电导gNa逐渐增大时,系统的反相簇放电从“sup-Hopf/fold cycle”型反相簇放电逐步过渡到“fold/fold cycle”型反相簇放电,这种转迁会经由一种中间型的过渡型簇放电.在耦合神经元表现为反相“sup-Hopf/fold cycle”型簇放电时,在一个簇放电中的峰与峰之间的相位差的差值相差很大.因此,簇放电未达到反相同步.随着钠电导gNa逐渐增大,耦合神经元表现为反相“fold/fold cycle”型簇放电,此时,簇放电中的峰与峰之间的相位差的震荡幅度逐步减小.因此,簇放电表现为反相同步.因为在哺乳动物的呼吸节律中发现过多种簇放电振荡模式,因而耦合簇放电同步模式的研究对认识pre-Bötzinger复合体中大规模网络的动力学有着重要的意义,为进一步探索呼吸节律的产生机制提供有益的思考.
1 Ivanchenko M V,Osipov G V,Shalfeev V D,et al.Phase synchronization in ensembles of bursting oscillators.Physical Review Letters,2004,93(13):134101
2 Ivanchenko M V,Osipov G V,Shalfeev V D,et al.Network mechanism for burst generation.Physical Review Letters,2007,98(10):108101
3 Rulkov N F.Modeling of spiking-bursting neural behavior using two-dimensional map.Physical Review E,2002,65(4Pt1):041922
4 Cao H,Sanjuán M A F.A mechanism for elliptic-like bursting and synchronization of bursts in a map-based neuron network.Cognitive Processing,2009,10(S1):23~31
5 王青云,石霞,陆启韶.神经元耦合系统的同步动力学.北京:科学出版社,2008(Wang Q Y,Shi X,Lu Q S.Synchronization dynamics for coupled neuron systems.Beijing:China Science Publication,2008(in Chinese))
6 Wang Q Y,Lu Q S.Synchronization transition induced by synaptic delay in coupled fast-spiking neurons.Internation-al Journal of Bifurcation and Chaos,2008,18(4):1189~1198
7 Gu H G,Li Y Y,Jia B,et al.Parameter-dependent synchronization transition of coupled neurons with co-existing spiking and bursting.Physica A:Statistical Mechanics and its Applications,2013,392(15):3281~3292
8 Wang QY,Lu Q S,Wang H X.Transition to complete synchronization via near-synchronization in two coupled chaotic neurons.Chinese Physics,2005,14(11):2189~2195
9 Gaiteri C,Rubin JE.The interaction of intrinsic dynamics and network topology in determining network burst synchrony.Frontiers in Computaional.Neuroscience,2011(5):1~10
10 Best J,Borisyuk A,Rubin J,Terman D,Wechselberger M.The dynamic range of bursting in a model respiratory pacemaker network.SIAM Journal on Applied Dynamical Systems,2005,4(4):1107~1139
11 Rubin J,Terman D.Synchronized bursts and loss of synchrony among heterogeneous conditional oscillators.SIAM Journal on Applied Dynamical Systems,2002,1(1):146~174
12 Dunmyre JR,Rubin JE.Optimal intrinsic dynamics for bursting in a three-cell network.SIAM Journal on Applied Dynamical Systems,2010,9(1):154~187
13 Butera R J,Rinzel J,Smith J C.Models of respiratory rhythm generation in the pre-Bötzinger complex.II.Populations of coupled pacemaker neurons.Journal of Neurophysiology,1999,82:398~415
14 平小方,刘深泉,任会霞,汪雷.呼吸中间神经元及呼吸网络模型的动力学分析.动力学与控制学报,2015,13(3):215~223(Ping X F,Liu SQ,Ren H X,Wang L.Dynamic analysis of interneuron and respiratory network model.Journal of Dynamics and Control,2015,13(3):215~223(in Chinese))
15 袁丹丹,陈汐,段利霞.Pre-Bötzinger复合体中耦合神经元簇同步模式及转迁的分岔分析.动力学与控制学报,2015,13(4):266~271(Yuan D D,Chen X,Duan L X.Synchronous bursting patterns and transitions of coupled neurons in pre-Bötzinger complex.Journal of Dynamics and Control,2015,13(4):266~271(in Chinese))
16 Yuan D D,Chen X,Meng X Y.Transition mechanisms of bursting in a two-cell network model of the pre-Bötzinger complex.International Journal of Bifurcation Chaos,2015,25(5):1550069
17 Izhikevich EM,Neural excitability,spiking and bursting.International Journal of Bifurcation and Chaos,2000,10(6):1171~1266
Received 6 January 2016,revised 11 March 2016.
*The project supported by the National Natural Science Foundation of China(11472009,11502073),Science and Technology Projectof Beijing Municipal Commission of Education(KM201410009012)and Construction Plan for Innovative Research Team of North China University of Technology(XN129)and Doctoral Foundation of Henan Polytechnic University(B2012-107)
†Corresponding author E-mail:duanlx@ncut.edu.cn
STUDY ON ANTI-PHASE BURSTING MODE AND SYNCHRONIZATION OF COUPLED NEURONS IN PRE-BÖTZINGER COMPLEX*
Liu Jing1Cao Qinyu1Wang Zhijian1Zhao Yong2Duan Lixia1†
(1.College of Science,North China University of Technology,Beijing 100144,China)
(2.School of Mathematics and Information Science,Henan Polytechnic University,Jiaozuo 454000,China)
In this paper,anti-phase bursting mode and synchronization transition in the two coupled excitatory neurons in pre-Bötzinger complex are examined.When the parameter of sodium conductance varies within a certain range,the two coupled neurons exhibit anti-phase“sup-Hopf/fold cycle”and“fold/fold cycle”bursting.By calculating the phase difference of spikes in the bursting,we reveal the relationship between different bursting patterns and synchronization transition.
bursting,phase difference,anti-phase synchronization,fast/slow decomposition,pre-Bötzinger complex
10.6052/1672-6553-2016-029
2016-01-06收到第1稿,2016-03-11收到修改稿.
*国家自然科学基金资助项目(11472009,11502073)、北京市教委科技计划项目(KM201410009012)、北方工业大学“科研创新团队建设计划”项目(XN129)和河南理工大学博士基金项目(B2012-107)
†通讯作者E-mail:duanlx@ncut.edu.cn
