山丘区中小河流边滩植被分布驱动因子及响应关系
2017-06-06余根听夏继红毕利东汪颖俊林立怀曹伟杰伊紫函
余根听,夏继红†,毕利东,汪颖俊,林立怀,曹伟杰,伊紫函
(1.河海大学水利水电学院,210098,南京;2.龙游县水利局,324400,浙江龙游)
山丘区中小河流边滩植被分布驱动因子及响应关系
余根听1,夏继红1†,毕利东1,汪颖俊2,林立怀1,曹伟杰1,伊紫函1
(1.河海大学水利水电学院,210098,南京;2.龙游县水利局,324400,浙江龙游)
河岸带滩地植被是维持河流系统健康的重要屏障。以浙江省龙游县灵山港边滩为研究对象,运用聚类分析和RDA约束性冗余分析等方法,分析中小河流边滩植被分布特征及其主要驱动因子,并建立各驱动因子对植被分布的响应关系。结果表明:灵山港滩地植被种类丰富多样,共有62种植被(被子植物61种,蕨类植物1种),其植被群落结构包括9种类型;滩地相对水面高差因子、滩地形态系数和水文特性是植被分布空间差异性的主要影响因子,各因子贡献率分别为37.50%、27.50%和16.82%,各因子对植被分布的驱动响应机制不同。其中:在滩地相对水平面高差因子作用下,植被从滩地外缘到滩地内部,耐水性由强到弱,丰富性由低到高,生物量由小到大;当边缘线发育系数SDI值处于2.0~4.0之间,且滩地狭长指数Pe/Pa值在0.12~0.3之间时,至少有60%的保证率使得滩地植被多样性维持在1.03~1.96的高水平;水文特性能够影响植被的形态特征和滩地的生境条件,水文作用下,滩地外围砂粒质量分数随着离水距离增大而逐渐减小,植被类型也会随着离水距离的增加,从喜湿耐冲型到中生型过渡。掌握滩地植被的分布特征及其影响因子,以期为河道生态修复,提供理论依据。
山丘区; 中小河流; 滩地; 植被分布特征; 驱动因子
滩地植被是发挥滩地功能的重要因素,植被覆盖的变化是生态环境变化的体现[1],一定宽度的滩地植被,可以通过过滤、渗透、吸收、滞留和沉积等机械、化学和生物功能效应,使进入地表和地下水的污染物毒性及污染程度降低,起到控制非点源污染、改善河流水文状况和水质的作用[2-3]。植被还可以通过改变微弯河岸扰动压强大小,影响河岸带滩地侧向潜流交换和物质能量传递,进而影响河流新陈代谢[4-5],并且植被分布下的滩地生态边缘效应、缓冲边缘效应和社会经济边缘效应,对河道的建设管理和开发利用均具有深刻的影响[6]。近年来,由于人为采砂和侵占等干扰,滩地植被资源减少,功能严重退化,这一问题已引起社会各界的认识和重视。例如:高敏等[7]通过二维数学模型,对植被化的漫滩水流进行流速分布研究;王月容等[8]对滩地植被缓冲带进行划分及功能和生态重建;闫静等[9]、陈正兵等[10]分析了不同植物种类和不同排列方式对水流结构的影响效应;Dwire等[11]、Burton等[12]分析不同环境梯度及城市化下的植被种类组成及结构效应;Moffatt等[13]分析植被群落的种类组成、多样性及周围土地利用类型。但是,以往学者的研究多是围绕植被对水流结构的影响方面,或是围绕研究所在地的植被基本情况及功能阐述方面,鲜有对植被的影响因子强度效应进行具体分析。基于此,笔者以龙游县灵山港滩地为研究对象,分析植被分布的特点及其主要驱动因子,深入探讨不同因子对植被分布的响应关系,以期为滩地生态修复中,植被的选择与布置提供理论依据。
1 研究区概况
浙江省龙游县位于E119°01′41″~119°19′52″,N28°44′10″~29°17′15″之间,地处浙江省西部金衢盆地,属浙中丘陵盆地。灵山港源于海拔1 265 m的遂昌县和尚岭,在沐尘乡马戍口流入龙游县境内,由南向北贯穿龙游县中部,于龙游县城汇入衢江,是钱塘江上游衢江右岸的一条重要支流。灵山港流域属亚热带季风气候区,雨量丰沛,四季变化明显。多年平均气温17.3 ℃,年平均日照时间1 966 h,多年平均降水量为1 666.4 mm。河道在龙游县境内流域面积367.6 km2,主流长43.79 km,平均比降2.45‰,是典型的山丘区中小河流。河道沿线滩地资源丰富,据调查统计,2014年河道沿线滩地总面积为1.66 km2。
2 材料与方法
2.1 监测点布设与取样
滩地植被通过布置监测样方进行调查。研究将灵山港分为上、中、下游与河口4个区段,每个区段内选取3个典型滩地(河口段除外),上游区段包括溪口四桥滩地(L1)、江潭滩地(L2)和下徐桥滩地(L3);中游区段包括寺下滩地(L4)、梅村滩地(L5)和周村滩地(L6);下游区段包括姜席堰滩地(L7)、上扬村滩地(L8)和高铁桥滩地(L9);河口段为彩虹桥滩地(L10)。典型滩地分布见图1(a)。每个典型滩地内布设一定数量的乔木样方(10 m×10 m)、灌木样方(5 m×5 m)和草本样方(1 m×1 m)。草本样方布置于滩头、滩中和滩尾断面上,每个断面上按照离水边缘距离大小,每隔7.5 m布置1个草本样方;同时,通过野外流速仪,监测典型滩地边缘水流流速。植被监测样方和流速监测点布置见图1(b)。

L1:溪口四桥滩地,L2:江潭滩地,L3:下徐桥滩地,L4寺下滩地,L5:梅村滩地,L6:周村滩地,L7:姜习堰滩地,L8:上扬村滩地,L9:高铁桥滩地,L10:彩虹桥滩地。以下类同。L1: Xikousiqiao floodplain, L2: Jiangtan floodplain, L3: Xiaxuqiao floodplain, L4: Sixia floodplain, L5: Meicun floodplain, L6: Zhoucun floodplain, L7: Jiangxiyan floodplain, L8: Shangyangcun floodplain, L9: Gaotieqiao floodplain, and L10: Caihongqiao floodplain. The same below.图1 监测点断面及监测点样方布设Fig.1 Map of monitoring sections and plots
对乔木与灌木,主要记录样方内其数量、种类、树高、胸径和冠幅等参数;对草本植被,主要通过摘取、分类和分装,清点株数,并带回实验室,经称重、杀青、烘干和再称重等处理,获得植被鲜质量和干质量。取样调查分别于2015年4月、2015年10月和2016年5月进行。
2.2 数据处理
2.2.1 主要植被参数及处理 根据监测数据,分别计算各滩地监测断面草本植被生物多样性指数[14]和生物量[15],并用均值代表滩地植被指标参数。对于乔木与灌木,主要计算乔灌木的空间测度指数(混交度)[16]、分布均匀度(角尺度)[17]和叶面积指数(LAI)[18]等参数。为了获取植被空间变异规律,采用半变异函数[19]进行分析,描述植被空间连续变异,反映植被不同距离测算值之间的变化。
2.2.2 环境因子数据处理 针对滩地的特点,主要选取滩地相对水面高差(ButtElev)、土壤吸湿系数(HygrCoef)、滩地离水距离(Distance)、滩地形态系数(Shapindx)及水文特性(HydrChar)5个环境因子,通过现场监测及Google earth测量,获得相关因子的数据。其中:ButtElev选取滩地植被样方所在位置与河道水平面之间的垂直距离表示;HygrCoef选取现场样方位置表层20 cm土层测算;Distance选取监测样方与水边的水平距离表示见图1(b);Shapindx选取欧式几何形态指标SDI及滩地狭长性指数Pe/Pa(短轴/长轴)表示;HydrChar选取弗劳德数(Fr)与紊动能(Eε)表示。
3 结果与分析
3.1 植被区系构成及分布特征
3.1.1 总体特征 现场调查发现,灵山港沿线滩地共有61种被子植物,隶属于25科56属。其中,绝大部分为草本植物,主要有菊科、禾本科、蓼科、伞形科、苋科、石竹科、莎草科、荨麻科、豆科、玄参科、唇形科和茜草科等共58种,隶属于23科54属,菊科和禾本科分别占14.52%和17.74%,蓼科和伞形科分别占8.06%和6.45%,苋科和石竹科均占4.84%,莎草科、荨麻科、豆科、玄参科、唇形科和茜草科均占3.23%。仅发现1科1属蕨类植物,为阴石蕨(Humatarepens(L. f.) Diels)。另外,发现少量的核桃楸(JuglansmandshuricaMaxim.)、竹子(Bambusoideae)以及杨树(PopulusL.)等木本植物,共3科3属。
3.1.2 草本群落结构特点 灵山港滩地植被主要为草本植物,植被种类丰富,其群落结构存在较大差异。为准确划分草本植被的群落结构,应用SPSS,对典型滩地内草本植物种类进行聚类分析(表1)。由表1可知,灵山港滩地草本植被可以分为9类群落结构。从分布频率看,沿线滩地草本植物中出现频率较高的为I、II和III类群落;其中,I、II类群落主要为间接分布种,III类群落为广布种。可见,滩地植被中广布种和间接分布种最多,其次是内部分布种;另外,紧邻滩地外缘分布物种种类较少,主要为水蓼(Polygonumhydropiper)、积雪草(Centellaasiatica(L.) Urban)、沿阶草(OphiopogonbodinieriLevl.)等。由此可知,草本植物分布和出现频率存在较大差异,通过现场观测发现,产生这一现象的主要原因是受滩地几何形态、地形、地貌和水流条件等因素的影响。

表1 滩地草本植被空间分布类型
3.1.3 林分空间结构特点 灵山港沿线滩地中,木本植物有其独特的林分结构。现场观测发现,木本植物主要出现于溪口四桥滩地(L1)、寺下滩地(L4)、周村滩地(L6)和彩虹桥滩地(L10)这4个滩地中。各典型滩地中,木本植物的林分指标计算值如表2所示,木本植被个体胸径、株高和LAI指数都不同,其周边的草本植被类型也存在较大差异。滩地的木本LAI指数分别为30.11、28.04、34.74和44.84,基本呈现增长趋势,2项拟合度较高,相关系数达到0.991 4。其中,L4滩地的木本LAI指数略有减小,这是由于滩地破碎化程度较高,使植被分布密度受到极大的限制。从混交度和角尺度上看,混交度均小于0.05,木本植被单一化程度明显,角尺度虽有些许波动,但二者均值均小于0.475,都为均匀分布类型。总体而言,灵山港滩地木本植被产量较为丰富,分布类型单一,有一定的局部小气候效应特征。

表2 林分空间结构特征
3.2 驱动因子对植被分布的响应关系
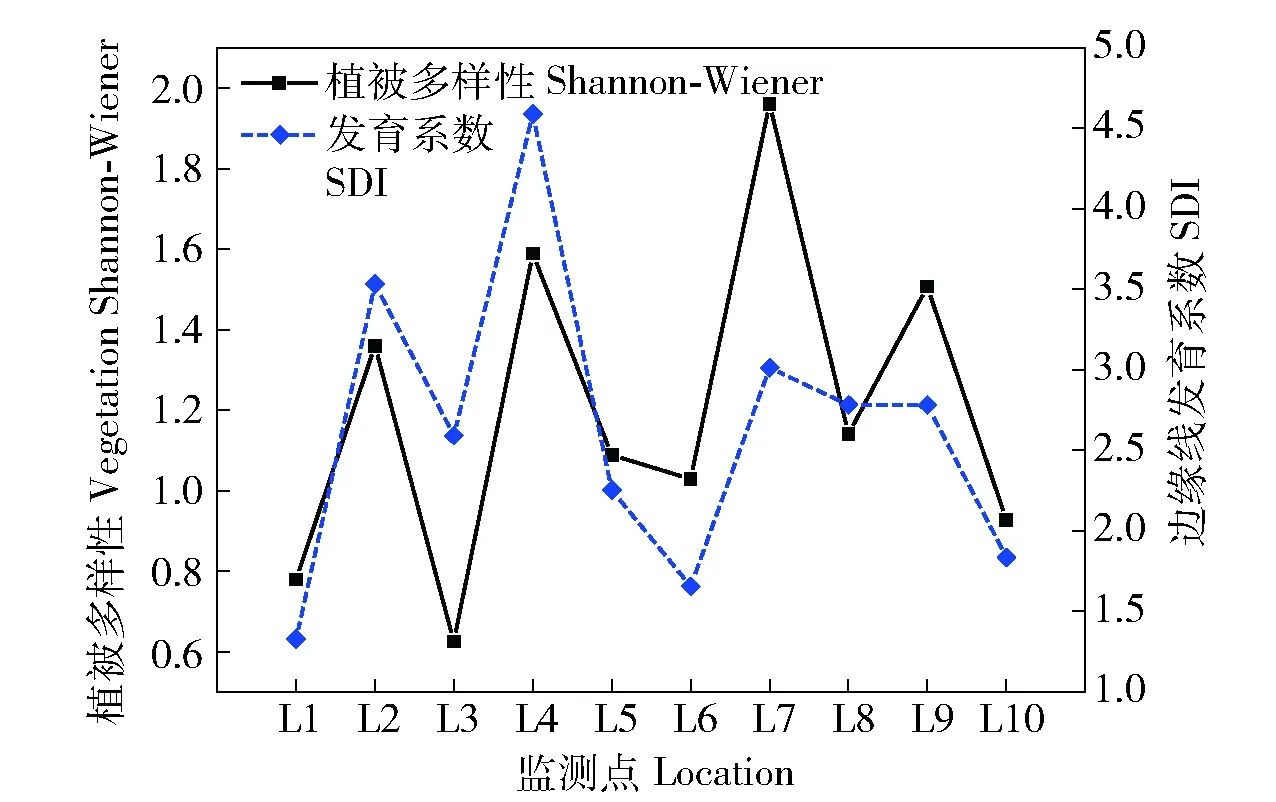
3.2.1 滩地植被空间关联性分析及关键驱动因子的确定 应用半变异函数来研究植被多样性的空间异质性,并确定其影响范围大小,计算结果由表3可知,L1、L5和L10滩地自相关性比较大,分别为0.266、0.222和0.253,表明随机性因子(局地气候、气象条件和局地水文特性等)在空间变异中占据主导地位;其余滩地自相关性较小,表明结构因子(地形地貌变化、地质构造和滩地形态等)在空间变异中起主导作用。基于此,将对灵山港滩地植被空间分布的驱动因子,及其敏感性大小进行深入分析。
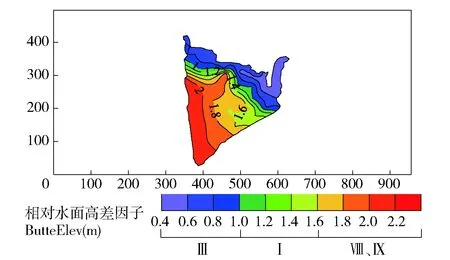
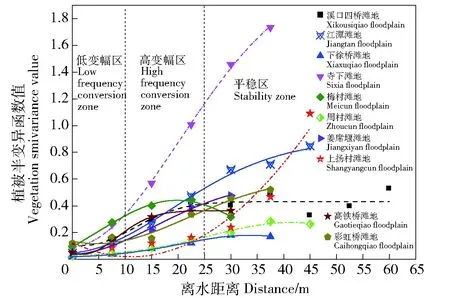
灵山港滩地植被空间分布受多个因子影响。对土壤吸湿系数、滩地离水距离、滩地形态系数、滩地相对水面高差因子和水文特性,进行KMO和Bartlett球形度检验,得KMO>0.7、P<0.05,表明因子选择具有结构效度,可做因子分析。通过Canoco软件RDA约束性冗余分析方法,分别计算各因子对植被分布的贡献率大小,通过贡献率,反映各因子对植被分布的驱动程度,各因子贡献率结果见图2,为了可观性较好,图中只展示了部分表1中的物种。由图2 RDA双序图可知,相对水面高差因子、形态系数和水文特性对植被分布的贡献率分别为37.50%、27.50%和16.82%,三者总体贡献率达到81.82%。土壤吸湿系数和滩地离水距离的贡献率分别为10.56%和7.62%,二者总体贡献率为18.18%。图中蓝色箭头代表草本植被种类,其长度在因子变量上的投影值代表物种与环境因子之间相关性的大小,若投影方向与因子正方向相同,表示物种与环境因子之间是正相关,反之为负相关。从贡献率和投影长度看,主要驱动因子对植被分布的敏感性程度顺序为相对水面高差因子>形态系数>水文特性。
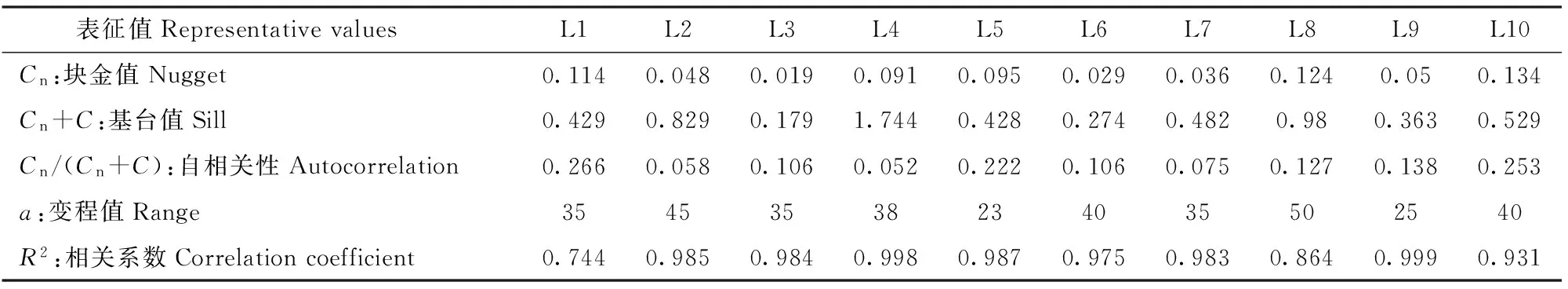
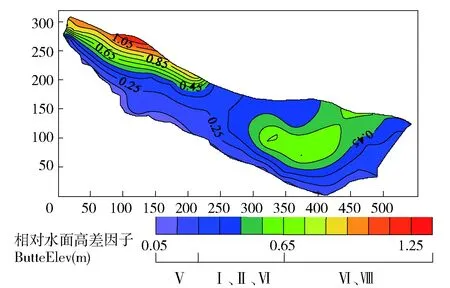
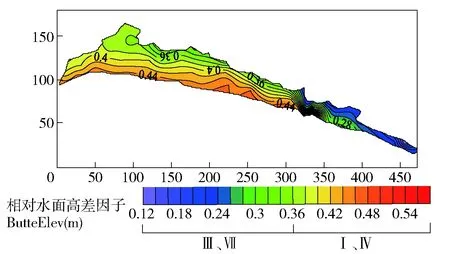
表3 滩地植被分布自相关性表
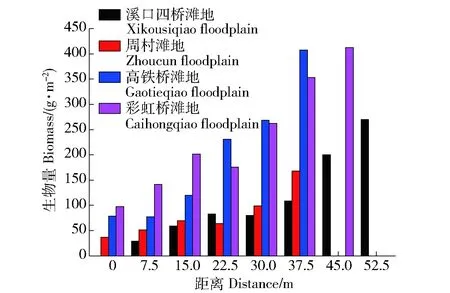
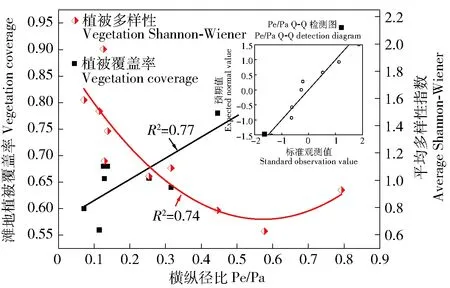
注:Cn:块金方差反映样点间多样性的变化程度;a:变程值用来衡量两样本间的相关性,距离d 1: 狗牙根Cynodon dactylon(Linn.)Pers. 2: 棒头草 Polypogon fugax Nees ex Steud. 3: 大画眉草Eragrostis cilianensis (All.) Link ex Vignolo-Lutati. 5: 看麦娘 Alopecurus aequalis Sobol. 6: 菵草Beckmannia syzigachne(Steud.)Fern. 7: 刺蓼Polygonum senticosum (Meisn.) Franch. et Sav. 10: 小飞蓬 Conyza canadensis (L.) Cronq. 11: 鼠麯 Gnaphalium affine D. Don. 13: 苣荬菜 Sonchus arvensis L. 16: 积雪草Centella asiatica (L.) Urban. 19: 水蓼Polygonum hydropiper. 21: 空心莲子草 Alternanthera Philoxeroides(Mart.)Griseb. 22: 刺苋 Amaranthus spinosus. 24: 毒芹 Cicuta virosa L. 26: 毛豆 Glycine max. 27: 阴石蕨 Humata repens (L. f.) Diels. 28: 毛茛 Ranunculus japonicus Thunb. 29: 通泉草 Mazus japonicus (Thunb.) O. Kuntze. 35: 雪见草 Salvia plebeia R. Br. 36: 旋覆花 Inula japonica Thunb. 37: 假稻 Leersia japonica (Makino) Honda. 38: 鸭跖草 Commelina communis. 40: 野老鹤草 Geranium carolinianum L. 41: 黄鹌菜Youngia japonica. 43: 簇生卷耳Cerastium caespitosum Gilib. The same below. ButtElev:Elevation factor; Shapindx:Shape coefficient; HydrChar:Hydrological characteristics; HygrCoef:Soil moisture absorption coefficient; Distance:Distance away from the water. 图2 物种与环境因子的RDA贡献率双序图Fig.2 RDA double sequence diagram of species and environmental factors 3.2.2 植被分布对滩地相对水面高差因子变化的响应关系 滩地相对水面高差因子是影响植被分布最显著的因子,以灵山港上游溪口四桥滩地(L1)、中游周村滩地(L6)、下游高铁桥滩地(L9)以及衢江口彩虹桥滩地(L10)为例,说明不同区段滩地内,相对水面高差因子对植被分布的影响。4个滩地内,植被分布的相对水平面高差云图见图3~图6。 图3 上游区段植被空间分布Fig.3 Spatial distribution of vegetation in the upper reaches 图4 中游区段植被空间分布Fig.4 Spatial distribution of vegetation in the middle reaches 图5 下游区段植被空间分布Fig.5 Spatial distribution of vegetation in the lower reaches 图6 河口植被空间分布Fig.6 Spatial distribution of vegetation in the estuary 可见,I、II、IV、VI、VIII、IX类植被主要生长在高差较大的滩地内部,满足其排水和向阳条件,III、V、VII类植被主要生长在高差较小的临水边,满足其水分需求较高的特点。各类植被的分布特点也存在一定的差异。I类植被(通泉草、牛毛毡、鼠麯、小飞蓬和狗牙根等)广泛分布于高差0.25~1.2 m内,且易形成大片群落。主要是因为该区间内,土壤受水流作用,土层松动,沙性明显,水分条件及排水较好,适宜I类植被生长。II类植被(小花糖芥、细风轮菜和旋覆花等)分布范围较广,主要分布在高差>1.8 m的位置。IV类植被(艾草、野豌豆和北美车前等)大多分布于高差1.8~2.1 m内,少部分会出现在临水边0.36~0.54 m处。V和VII类植被(沿阶草、积雪草、水蓼等)主要分布于高差<0.9 m的临水边,主要是因为积雪草叶片中的机械组织不发达,抗旱能力极差,是阴生湿生植物,而蓼科和莎草科等阳生湿生植被,根系不发达,没有根毛,但根与茎之间有通气的组织,以保证取得充足的氧气,生活在阳光充足、土壤水分饱和的沼泽地区或湖边。III类植被(棒头草、假稻、鸭跖草和酸模叶蓼等)会与V和VII类植被集群分布。VI类植被(刺蓼、阴石蕨和野菊等)主要分布于高差0.4~1.25 m(刺蓼、山大颜和毒芹)、1.8~2.4 m(阴石蕨和野菊)的位置。VIII和IX类植被主要分布于高差1.8~2.0 m的位置,在滩地内缘有少量出现。 由图7和图8可知,滩地植被从外缘到内缘(高差从小到大),生物量在逐渐增大,且植被多样性随着滩地相对水面高度的增加而增加,对应的植被扩散斑块(圆域面积)大小在逐渐扩增。总体而言,地形高度的差异,可以作为植被种类的分界线,滩地植被从滩地外缘到滩地内部的分布规律为:耐水性由强到弱,丰富性由低到高,生物量由小到大。 图7 滩地外缘至内缘植被生物量变化Fig.7 Biomass variation from outer to inner edge of floodplain 图8 不同相对水面高度下的植被多样性Fig.8 Vegetation Shannon-Wiener under different relative elevation of water 3.2.3 植被分布对形态系数的响应关系 滩地总体形态变迁是影响植被多样性的另一个重要因素,稳定的滩地生境是植被分布特征优化,以及发挥生态效益最大化的重要保障。由图9可知,随着滩地Pe/Pa指数的增大,植被覆盖率也随之增大。从图3~6亦可看出,短宽和窄长滩地对等高线分布及滩地断面植被容纳量有明显的影响。选择滩地边缘线发育系数SDI(即Richardson所提出的紧凑度C0的倒数,SDI=1时,滩地形态近似圆形)和滩地横纵径(短长轴)比值Pe/Pa(避免SDI较大时,滩地出现过分窄长的情况),来反映滩地形态的弯曲和狭长。Pe/Pa和SDI与植被多样性响应关系分别见图9和图10。 图9 Pe/Pa与植被覆盖度和多样性的响应关系Fig.9 Response of coverage and Shannon-Wiener to Pe/Pa 图10 SDI与植被多样性的响应关系Fig.10 Response of Shannon-Wiener to SDI 由图10可知,灵山港滩地SDI系数>2的占滩地总数的70%,说明滩地紧凑度不大,岸线几何形状较为复杂。在自然情况下,滩地植被多样性随着SDI系数的变化而变化,两者呈现同样的趋势效应,节点变化同一性极高。当SDI值处于2.0~4.0之间时,滩地平均植被多样性>1出现的频率占全局60%以上。图9显示,滩地植被多样性随着Pe/Pa的增大而减小;由Q-Q检测可知,各点分布于直线附近,且K-S检验的渐近显著性系数为0.580>0.05。因此,灵山港滩地的形态特征服从正态分布规律,滩地整体形态稳定性较好,而且,当Pe/Pa值在0.12~0.3之间时,植被多样性维持在1.03~1.96之间的次数出现6次占60%。即至少有60%的保证率使得SDI系数处于2.0~4.0之间,且滩地狭长指数Pe/Pa值在0.12~0.3之间时,滩地植被多样性较高。 3.2.4 植被分布对水文特性的响应关系 从植被空间变异特征(图11)可知,距离水边10 m以内,指数的平均差异性Δr(h)约为0.100,空间变异幅度较小,这主要是由于滩地临水边缘小区域受水流影响较大,植被物种差异性不明显,多为喜湿耐冲型植被,将这一区域称为低变幅区。距离水边10~25 m时,指数的平均差异性Δr(h)达0.206,空间变异幅度较大,这主要是由于微地貌的变化,使得滩地淹没所需水流深度增加,这一因素的限制,在平水位情况下,水力冲蚀削弱,主要生长中生型植被,植被种类丰富,将这一区域称为高变幅区。距离水边>25 m时,指数的平均差异性Δr(h)仅为0.052左右,无明显的空间变异性(寺下、上扬村受人为干扰,变异性依旧较大),此时结构因子正在逐渐替代水文特性成为主导因子,且由表3可知,变程值a均>25 m;因此,将这一区域称为平稳区,主要为乔灌草结构。 图11 植被空间相关性曲线Fig.11 Vegetation spatial correlation curve 将低变幅区与高变幅区,即离水边缘25 m以内的植被带,作为植被分布对水文特性响应关系的研究区域。各区段滩地外缘植被带土壤性质、植被多样性和生物量的变化见表4。 由表4可知,同一滩地低变幅区土壤中砂粒质量分数比高变幅区的大,且低变幅区的土壤质地类别中,壤质砂土出现频率极高,达到80%。究其原因,土壤颗粒组成受水文特性的影响极大,水流冲刷和淘蚀滩地岸线,土壤中的粉粒和黏粒等细粒物质大量流失,越靠近河流的淹没带,越容易受到其影响,致使粉黏粒质量分数下降,砂粒质量分数增加。水流作用改变了植物根系与土壤之间物质交换属性,使得植被的空间分布也存在异质性。 表4 植被分布对水文特性的响应 从植被形态可塑性角度看,为了适应高流速、大紊动的水流条件,滩地外缘植被的叶片和杆茎均为轻柔和狭长,能够顺水流方向倾伏,以克服水流的拖拽力及紊动卷携,从而保持在该条件下的生存能力。表现为在滩地外缘25 m带宽范围内,蓼科和棒头草出现的频率均占40%,菵草和狗牙根出现的频率均占30%,沿阶草出现的频率占10%,均为喜湿耐冲型植被,与内缘植被差异性较高。从植被生长属性角度看,灵山港滩地基本呈现上、下游区段植被多样性较高,生物量较低,而中游区段植被多样性较低,生物量较高的现象。这是由于上、下游水流适度的扰动及短期水位变动,为低多样性集群物种的存活率创造了条件,在主要植被类型基础上,新增细风轮菜、通泉草、小飞蓬、鼠麯、小花糖芥、鸭跖草和积雪草等植被物种,使得植被带宽内多样性指数较大;但由于物种茎秆及根系较小,生物量相对较低。而中游在集中的木本植被阻流缓冲和局地小气候效应影响下,植被带宽内水文效应薄弱,滩地生境稳定,种类单一化明显,多为阴石蕨、沿阶草和毒芹等,使得植被多样性较小;但由于庞大的小块根及高含水量,使得生物量较大。 1)由植被空间关联性分析,将灵山港滩地分为低变幅区、高变幅区和平稳区3个区域。其中,植被分布受随机因子与结构因子的共同影响,在滩地外缘至岸边,植被类型表现为喜湿耐冲型植被、中生植被、中生植被+乔木+灌木的过渡。 2)山丘区河道滩地植被分布的关键驱动因子为滩地相对水面高差因子、滩地形态系数和水文特性。从整体范围上看,植被分布对关键驱动因子敏感性大小依次为滩地相对水面高差因子>滩地形态系数>水文特性。 3)地形高度的差异可以作为植被种类的分界线,滩地植被的分布从滩地外缘到滩地内部,耐水性由强到弱,丰富性由低到高,生物量由小到大。 4)滩地不可能无限发育,有其自身的极限展长与极限展宽。当SDI系数处于2.0~4.0之间,且滩地狭长指数Pe/Pa值在0.12~0.3之间时,至少有60%的保证率,使得滩地植被多样性能维持在较高水平。 5)河道水文特性会从不同程度影响植被的空间关联性,如植被形态特征与生境条件。水文特性作用下,常表现为滩地外围植被带砂粒质量分数随离水距离增大而减少。外围植被带中,蓼科、棒头草、菵草、狗牙根和沿阶草等,对水文环境适应能力极佳,可用于生态修复建设过程中优先选择的对象。 [1] 廖建华,李丹勋,王兴奎,等. 长江上游植被覆盖的时空分异季节变化及其驱动因子研究[J].环境科学学报,2009,29(5): 1103. LIAO Jianhua, LI Danxun, WANG Xingkui, et al. The spatial-temporal distribution of seasonal vegetation changes and their driving forces in the upper reaches of the Yangtze River[J]. Acta Scientiae Circumstantiae, 2009, 29(5):1103. [2] 郭二辉,孙然好,陈利顶. 河岸植被缓冲带主要生态服务功能研究的现状与展望[J].生态学杂志,2011,30(8):1830. GUO Erhui, SUN Ranhao, CHEN Liding. Main ecological service functions in riparian vegetation buffer zone: research progress and prospects[J]. Chinese Journal of Ecology, 2011, 30(8):1830. [3] 孙彭成,高建恩,王显文,等. 柳枝稷植被过滤带拦污增效试验初步研究[J].农业环境科学学报,2016,35(2):314. SUN Pengcheng, GAO Jianen, WANG Xianwen, et al. Effectiveness of switchgrass vegetative filter strip in intercepting pollutants and promoting plant biomass[J]. Journal of Agro- Environment Science, 2016, 35(2):314. [4] 林俊强,严忠民,夏继红. 微弯河岸沿线扰动压强分布特性试验[J].水科学进展,2013,24(6):855. LIN Junqiang, YAN Zhongmin, Xia Jihong. Laboratory experiments on pressure distribution along sinuous riverbanks[J]. Advances in Water Science, 2013, 24(6):855. [5] 夏继红,陈永明,王为木,等. 河岸带潜流层动态过程与生态修复[J].水科学进展,2013,24(4):589. XIA Jihong, CHEN Yongmin, WANG Weimu, et al. Dynamic processes and ecological restoration of hyporheic layer in riparian zone[J]. Advances in Water Science, 2013, 24(4):589. [6] 夏继红,林俊强,姚莉,等. 河岸带的边缘结构特征与边缘效应[J].河海大学学报(自然科学版),2010,38(2):215. XIA Jihong, LIN Junqiang, YAO Li, et al. Edge structure and edge effect of riparian zones[J]. Journal of Hohai University(Natural Sciences),2010,38(2):215. [7] 高敏,槐文信,赵明登. 滩地植被化的顺直复式河道漫滩水流计算[J].节水灌溉,2008(6):21. GAO Min, HUAI Wenxin, ZHAO Mingdeng. Overbank flow calculation for straight compound channel with vegetated floodplain[J].Water Saving Irrigation,2008(6):21. [8] 王月容,周志翔,周金星,等. 长江滩地植被缓冲带类型及功能与生态重建[J].湖北林业科技, 2010(4):5. WANG Yuerong, ZHOU Zhixiang, ZHOU Jinxing, et al. Type,function and ecological reconstruction of vegetated buffer strips in Yangtze River Beach[J]. Hubei Forestry Science and Technology, 2010(4):5. [9] 闫静,唐洪武,田志军,等. 植物对明渠流速分布影响的试验研究[J].水利水运工程学报,2011(4):138. YAN Jing, TANG Hongwu, TIAN Zhijun, et al. Experimental study on the influence of vegetation on the velocity distribution of open channel flows[J]. Hydro-Science and Engineering, 2011(4):138. [10] 陈正兵,江春波.滩地植被对河道水流影响[J].清华大学学报(自然科学版), 2012,52(6):804. CHEN Zhengbing, JIANG Chunbo. Effect of floodplain vegetation on river hydrodynamics [J].Journal of Tsinghua University (Science and Technology), 2012,52(6):804. [11] DWIRE K A, KAUFFMAN J B, BROOKSHIRE E N J, et al. Plant biomass and species composition along an environmental gradient in montane riparian meadows[J]. Oecologia, 2004, 139(2):309. [12] BURTON M L, SAMUELSON L J. Influence of urbanization on riparian forest diversity and structure in the Georgia Piedmont, US[J]. Plant Ecology, 2008, 195(1):99. [13] MOFFATT S F, MCLACHLAN S M, KENKEL N C. Impacts of land use on riparian forest along an urban-rural gradient in southern Manitoba[J]. Plant Ecology, 2004, 174(174):119. [14] 李悦,马溪平,李法云,等.细河河岸带植物多样性研究[J].广东农业科学,2011(19):131. LI Yue, MA Xiping, LI Fayun, et al. Study on plant species diversity of riparian zone along Xi river[J].Guangdong Agricultural Sciences,2011(19):131. [15] 孟伟,张远,渠晓东等.河流生态调查技术方法[M].北京:科学出版社,2011:65. MENG Wei, ZHANG Yuan, QU Xiaodong, et al.Technical methods of river ecological investigation[M]. Beijing: Science Press, 2011:65. [16] 惠刚盈,胡艳波,赵中华. 基于相邻木关系的树种分隔程度空间测度方法[J].北京林业大学学报,2008,30(4):131. HUI Gangying, HU Yanbo, ZHAO Zhonghua. Evaluating tree species segregation based on neighborhood spatial relationships[J]. Journal of Beijing Forestry University, 2008, 30(4):131. [17] 惠刚盈,Gadow K V,胡艳波.林分空间结构参数角尺度的标准角选择[J].林业科学研究,2004,17(6):687. HUI Gangying, Gadow K V, HU Yanbo. Theoptimum standard angle of the uniform angle index[J]. Forest Research, 2004, 17(6):687. [18] 麻雪艳,周广胜. 春玉米最大叶面积指数的确定方法及其应用[J].生态学报,2013,33(8):2596. MA Xueyan, ZHOU Guangsheng. Method of determining the maximum leaf area index of spring maize and its application[J]. Acta Ecologica Sinica, 2013, 33(8):2596. [19] 李斌.青藏高原植被时空分布规律及其影响因素研究[D]北京:中国地质大学出版社,2016:62. LI Bin. Study of vegetation’s spatial and temporal distribution and influencing factors on the Tibetan Plateau[D]. Beijing: China University of Geosciences,2016:62. Vegetation distribution and its driven-forces on the floodplains of small and medium rivers in hilly area YU Genting1, XIA Jihong1, BI Lidong1, WANG Yingjun2, LIN Lihuai1, CAO Weijie1, YI Zihan1 (1.College of Water Conservancy and Hydropower, Hohai University, 210098, Nanjing, China; 2.Water Conservancy Bureau, 324400, Longyou, Zhejiang, China) [Background] Vegetation in riparian zone is an important buffer to sustain the health of a river system. It is a typical ecotone characterized as edge effect, and plays a significant role in the conservation of river system health. Therefore, it is vital to know how the vegetation in floodplain distributes and what the driven-forces of the distribution are. [Methods] Taking the Lingshan River, in Longyou County, Zhejiang Province, as a typical example of small and medium rivers in hilly area, the characteristics of the vegetation distribution and its driven-forces were studied. After investigating the vegetation species in the quadrat sampling, the data was calculated and analyzed using the Clustering Analysis (CA) in SPSS software, and the Redundancy Analysis (RDA) in Canoco5 software and through which the composition and cluster numbers of vegetation species, the cosine of the angle at the origin of the two vectors, the contribution rate of driven-forces were calculated. On the basis of GPS site survey and the maps downloaded from Google Earth, the elevation and shape coefficient were analyzed by Digital Elevation Model (DEM) in Surfer software. Then combined with the regression analysis, the relationship between the vegetation and driven-forces were obtained. [Results] In the floodplain of Lingshan River, there were 62 vegetation species (61 kinds ofAngiospermaesand one kind ofPteridophyta) and the vegetation community structure was mainly divided into 9 classes. The major three driven-forces of the vegetation distribution were elevation factor (ButtElev), shape coefficient (Shapindx) and hydrological characteristics (HydrChar), and their contribution rates were 37.50%, 27.50%, 16.82%, respectively. Particularly, under the influence of ButtElev, from outer to inner edge of floodplain, the moisture capacity varied from strong to weak, the diversity varied from low to high, and the biomass varied from small to large. There was at least 60 percent of the guarantee rate making the vegetation diversity between 1.03 and 1.96 when the SDI coefficient was between 2.0 and 4.0, and the Pe/Pa was between 0.12 and 0.3. The hydrological characteristics affected the morphological plasticity of vegetation and the habitat conditions of the floodplain. The sand content in the outer zone of the floodplain decreased with the increase of the distance from the water, and the anti-scour ability and moisture capacity of the vegetation decreased from strong to weak. [Conclusions] The elevation factor, shape coefficient and hydrological characteristics are the key driven-forces of vegetation distribution in small and medium rivers in hilly area. When we design ecological restoration projects, the three factors should be focused on, and the suitable measures should be employed in order to control and optimize the three factors. hilly area; small and medium rivers; floodplain; vegetation distribution; driven-forces 2016-09-19 2017-03-23 项目名称: 国家自然科学基金“蜿蜒型河岸带潜流层水动力学机制及溶质运移规律研究”(41471069);浙江省水利科技项目“龙游县中小河流滩地时空演化机理及生态修复技术研究”(RC1527) 余根听(1992—),男,硕士研究生。主要研究方向:河岸带生态机理。E-mail: yugenting@hhu.edu.cn †通信作者简介: 夏继红(1970—),男,教授,博士生导师。主要研究方向:河岸带生态机理。E-mail: syjhxia@hhu.edu.cn X171 A 2096-2673(2017)02-0051-11 10.16843/j.sswc.2017.02.007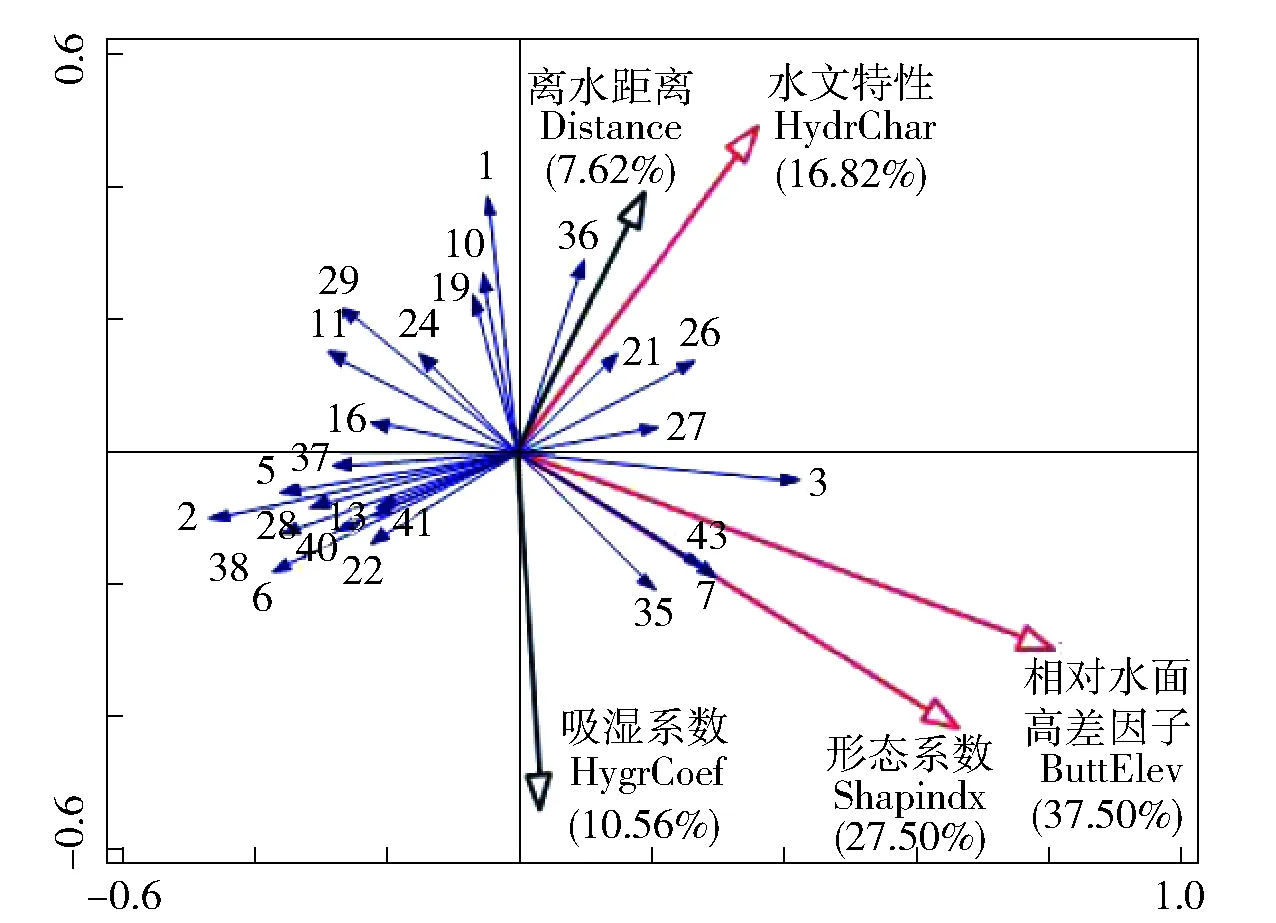



4 结论
