3种龙胆科植物的资源分配对策
2017-06-05侯勤正文静李昊聪任昱兰沙建华赵东光
侯勤正,文静,李昊聪,任昱兰,沙建华,赵东光
(西北师范大学生命科学学院,甘肃 兰州 730070)
3种龙胆科植物的资源分配对策
侯勤正,文静,李昊聪,任昱兰,沙建华,赵东光
(西北师范大学生命科学学院,甘肃 兰州 730070)
选择青藏高原东缘同一生境类型下具有不同交配系统特点和不同传粉者丰度的3种龙胆科植物湿生扁蕾(Gentianopsispaludosa)(自花授粉)、祁连獐牙菜(Swertiaprzewalskii)(高昆虫访问异花授粉)和线叶龙胆(Gentianafarreri)(低昆虫访问异花授粉)为对象,研究其资源分配特征。结果表明,1)3种植物的花粉胚珠比例(P/O)存在显著差异(P<0.05),湿生扁蕾的P/O显著低于另外两种异花授粉植物的(P<0.05),祁连獐牙菜的P/O显著低于线叶龙胆的(P<0.05);2)3种植物的繁殖分配比例存在差异,湿生扁蕾的繁殖分配比例显著高于另外两种异花授粉植物的(P<0.05),线叶龙胆的繁殖分配比例显著高于祁连獐牙菜的(P<0.05);3)在繁殖分配比例与个体大小关系上,湿生扁蕾与其个体大小不存在显著相关关系(P>0.05),而另外两种异花授粉植物则与其各自个体大小之间存在显著负相关关系(P<0.05)。研究表明,交配系统类型和传粉者丰度与植物的资源分配模式有关,尽管3种植物的繁殖绝对投入量都与其个体大小显著正相关(P<0.05),但自花授粉植物相对于异花授粉植物具有较高的繁殖分配比例,并且这种较高的繁殖分配比例不受个体大小影响,这种高繁殖投入的资源分配模式可能与自花授粉植物具有繁殖保障,从而降低其繁殖代价有关。
资源分配;交配系统;繁殖分配;传粉者丰度;自花授粉;异花授粉
在植物的整个生长发育过程中,可以被用于分配到生长、贮藏、繁殖和防御等功能的资源是先从环境中进行获取,而后实现配置的。然而,环境中的资源往往是一定的或有限的,因此,各种功能对于有限的可用于分配的资源始终存在着竞争,而植物必须权衡这些功能间的资源分配[1-2]。当前的研究焦点集中在异质环境下植物的资源分配策略方面,这是由于环境的异质性会造成同一基因型植物资源分配的巨大差异,亦即异质环境植物的表型可塑性(phenotypic plasticity),例如:海拔的差异[3]、生境的差异[4-6]等,因而也为研究植物的资源分配和进化稳定策略提供了理想条件[7]。然而,资源分配的差异还体现在交配系统上,交配系统的差异不但会影响植物个体的适合度和遗传结构,同时也会对花构成、性比分配和资源投资模式造成影响[8-10]。
植物的交配系统特点与其表型之间存在功能联系,同时这种联系会影响物种的生态行为[11-12]。根据植物对资源利用的利益最大化原则,具有不同交配系统的植物在雌雄功能的投资上表现出典型的差异性,例如Cruden[13]早在1977年就发现异交植物的花粉胚珠比例远大于自交植物,此结果后来得到了很多数据的支持,并且发现性分配比例随着自交率的增加而降低[14-16]。之后,人们的目光又扩大到花吸引结构(如花被、花蜜量)上,众多研究表明随着自交率的增加,植物在花吸引结构上的投资逐渐减小[10,14,17-18]。为最大化利用有限的可获得资源,植物在繁殖和营养器官的资源投资上显然存在着权衡[19],而这种权衡关系在不同交配系统类型的植物之间也存在差异,例如随着自交率增加,8种毛茛科植物花期繁殖分配降低[10]。然而,关于此类问题的研究较少,要想得出普遍规律则需要更多的试验来证实。自交率的高低与传粉者的丰度有着密不可分的关系,虽然不同交配系统类型植物的资源分配存在差异,但不同的交配系统特点并不一定意味着自交率的差异,因而考虑传粉者的数量将更有助于理解,采取不同交配系统特点的植物其资源分配策略,并为研究植物生活史策略提供理论依据。另外,研究表明,植物的繁殖分配与植株个体大小存在依赖效应(size-dependent effect),即随着植株个体增大,繁殖绝对投入会相应增加,但繁殖分配比例(相对投入)会减少[4,20-22],但是也有学者认为,繁殖分配与个体大小并不存在依赖效应(size-independent effect)[23]。正如前文所述,植物交配系统特点与其表型之间存在功能联系,那么,传粉者的多寡是否会对植物的繁殖分配造成影响?而这种繁殖分配比例是否与各自的植株大小之间存在依赖关系?显然,对于此问题的回答有助于扩大人们对植物资源分配策略的理解。
青藏高原是陆地上最为极端环境之一,其恶劣的生态环境,如低温、多雨、强辐射和短生活周期等,影响着高山植物的生长发育和后代延续,但是该地区却拥有许多形态特化的植物并具有较高的物种多样性,这一现象引起了诸多进化学者的高度关注[24]。另外,青藏高原多样化的植被群落会造成植物生境的巨大差异[25],因此,本研究在青藏高原同一地区选择在交配系统特征和传粉者丰度上具有差别的3种龙胆科植物,湿生扁蕾(Gentianopsispaludosa)、线叶龙胆(Gentianafarreri)和祁连獐牙菜(Swertiaprzewalskii)作为研究对象,探讨这3种植物的繁殖分配特征。3种植物表现出明显差异的交配模式及传粉频率(表1):湿生扁蕾的繁殖成功不需要传粉者,表现为延迟自交的繁育系统特征[26],而祁连獐牙菜和线叶龙胆的繁殖成功需要传粉媒介[27-28];在传粉者数量上,湿生扁蕾在青藏高原地区没有传粉者,祁连獐牙菜的传粉者为克什米尔熊蜂(Bumbuskeshimirensis)和蜜蜂(Apismellifera),两种昆虫的访问频率约为1.302次·(花·小时)-1,线叶龙胆的主要传粉者为克什米尔熊蜂和苏氏熊蜂(B.sushikini),两种昆虫的访问频率约为0.005 5次·(花·小时)-1[26-28]。这3种植物亲缘关系相近但在交配系统特征和传粉频率上差异显著,这为研究不同交配系统特点植物的繁殖分配提供了理想材料,因而本研究通过对这3种植物繁育系统的研究,旨在解决以下问题:1)这3种植物的繁殖分配特点各是什么?其繁殖分配是否与个体大小之间存在依赖效应?2)不同的繁殖分配特性与其交配系统特点和传粉者的多寡之间存在什么关系?

表1 3种植物的交配模式、传粉者种类和访花频率
1 材料和方法
1.1 研究区概况
本研究野外观察和试验地点均在青海省门源县中国科学院海北高寒草甸生态系统定位研究站(以下简称海北站)开展。海北站地处青藏高原东北部,祁连山东段北支冷龙岭南麓,大通河河谷的西北部,37°29′-37°45′ N,101°12′-101°23′ E,海拔约3 200 m,年平均气温为-1.7 ℃(最高气温达27.6 ℃,最低气温为-37.1 ℃),平均降水量为426~860 mm,降水多集中在5月-9月。
1.2 研究材料
试验所用材料为青藏高原东缘分布的龙胆科的3种常见植物:湿生扁蕾、祁连獐牙菜和线叶龙胆。湿生扁蕾为两年生草本,盛花期在7月底8月初;茎单生,直立或斜升,近圆形,在基部分枝或不分枝;花单生茎或分枝顶端;花梗直立,花冠筒状漏斗形,具4个花冠檐。祁连獐牙菜为多年生草本,盛花期在7月底8月初;茎直立,不分枝;简单或复聚伞花序狭窄,花冠黄绿色,裂片披针形。线叶龙胆为多年生草本,盛花期在9月中旬;花枝多数,丛生,铺散;花单生枝顶,倒锥状桶形。
1.3 研究方法
1.3.1 样品采集 由于3种植物盛花期不同,故2013年8月1日-3日采集湿生扁蕾和祁连獐牙菜样品,2013年9月15日采集线叶龙胆样品。在生境相同区域随机选取25~30个1 m×1 m样方,每一样方各随机选取3~5株完整个体,挖取整株植物,根部用清水洗净后晾干,分株装于信封袋后带回实验室。
1.3.2 样品生物量的测定 在样品生物量测定之前,对各植株进行清理,首先小心去除可能混夹于茎叶中的杂草及其它杂质,再用清水冲去根部所粘连的土壤。之后将清理完毕的样本放置于烘箱中,在80 ℃下烘干24 h,然后用1/10 000电子天平分别对各样品的整株、地下部分、茎叶和繁殖器官进行称重测量,统计并计算各指标。其中个体大小为植株整体干重,繁殖投入为繁殖器官干重,繁殖分配比例为繁殖器官干重占植株整体干重百分比。
1.3.3 数据分析 根据个体大小(X)和繁殖组分(Y)的异速关系,采用异速模型Y=aXb来分析个体大小与繁殖投入及繁殖分配的相关性。即,将个体大小、繁殖投入、繁殖分配的所有数据在Excel软件中进行自然对数转换后再用SPSS 13.0 for Windows统计软件包的Linear regression进行线性回归分析。用非参数检验(Nonparametric test)中的单样本K-S检验(1-Sample K-S test)检验数据是否服从正态分布。对于服从正态分布的数据,用独立样本T检验(Independent-samples T test)比较两组数据的差异;用单因素方差分析(One-way ANOVA)和Posthoc-LSD比较3组或者3组以上数据的差异;对于不服从正态分布的数据,用非参数检验中2 Inde-pendent Samples比较两组数据的差异,用K Inde-pendent Samples 比较3组或者3组以上数据的差异。
2 结果与分析
2.1 不同植物的花粉胚珠比例
3种植物的花粉胚珠比存在显著差异(P<0.05),其中自花授粉植物湿生扁蕾的P/O为(46±11),高昆虫访问频率异花授粉植物祁连獐牙菜的P/O为(266±36),低昆虫访问频率异花授粉植物线叶龙胆的P/O为(694±245)(图1)。
2.2 不同植物的繁殖投入
由于繁殖器官生物量(R)与个体大小(V)存在异速增长效应[21],因此对R-V关系分析均采用对数(lg)转换后的线性关系分析。结果表明,3种不同交配系统类型植物的繁殖器官绝对投入量(lgR)均随着个体大小(lgV)的增大而增大,且呈现出显著正相关关系(P<0.05)(图2)。

图1 3种植物的花粉胚珠比例及繁殖分配比例
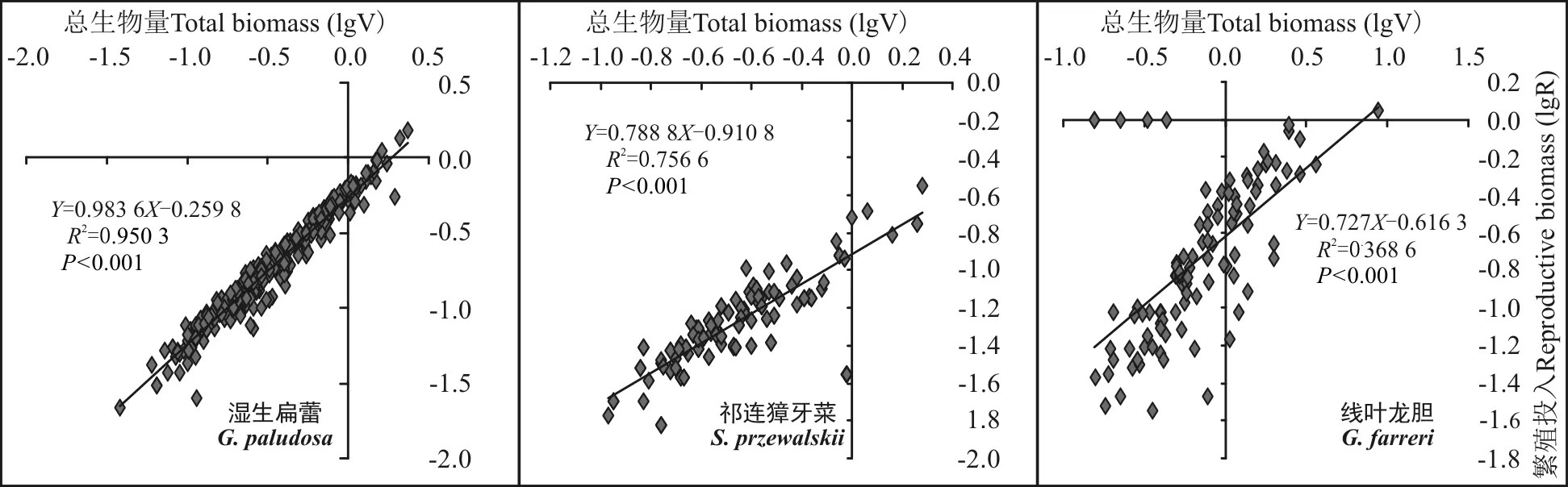
图2 3种植物的繁殖器官生物量(lgR)与个体大小(lgV)的相关性分析
2.3 不同植物的繁殖分配比例
在种群水平上,3种交配系统类型植物的繁殖分配比例具有显著差异(P<0.05),其中湿生扁蕾的繁殖分配比例最高(0.56±0.01),祁连獐牙菜的繁殖分配比例最低(0.16±0.01),而线叶龙胆的繁殖分配比例介于二者之间(0.22±0.03)(图1)。
从个体水平看,湿生扁蕾繁殖分配比例与个体大小之间不存在显著相关关系(P>0.05),表明其繁殖投入的比例并不会随个体变化而变化;祁连獐牙菜和线叶龙胆的繁殖分配比例与个体大小显著负相关(P<0.05),表明随着植株个体增大,其繁殖分配比例下降(图3)。

图3 3种植物的繁殖分配比例与个体大小的相关性分析
3 讨论
植物一般通过遗传分化和异质环境的表型可塑性两种对策来适应环境[29-30]。本研究结果显示,不同植物的P/O之间存在显著差异,自花授粉植物湿生扁蕾的P/O显著低于另外两种异花授粉植物,这与前人所研究的结果相同,如萝藦科、含羞草科[13]以及杠柳亚科[31]等。较低的花粉投入降低了自花授粉植物的资源消耗,从而使植物可以将更多资源投资到胚珠数目和种子发育,以利于子代繁育。在具有不同昆虫访问频率的异花授粉植物之间,低昆虫访问频率的线叶龙胆的P/O显著高于高昆虫访问频率的祁连獐牙菜,这可能是因为对于高度依赖传粉者的虫媒植物而言,低传粉者丰度为植物提供了进化压力,而增加雄性功能投资显然是对这种压力的适应。虽然本研究没有对线叶龙胆花粉的“质”(即花粉活力)进行检测,但其“量”的增加无疑可以扩大胚珠受精几率,提高传粉者频率低的植物的适合度。
根据权衡理论,植物需要将有限的资源在各功能之间做到合理的分配以达到其最优适合度,因而植物的资源分配模式可以很好反映植物的生活史策略,反过来说,植物的生活史策略可以通过植物的资源分配模式来反映[4,20]。前人研究结果显示,随着自交率的增加,植物在花吸引结构的投资上逐渐减小[10,18],相应的花期繁殖分配也降低[4,21-22],这是因为自交所带来的繁殖保障可以降低交配代价,但也有结果显示专性自交植物的繁殖分配比例相对异交植物反而更大[32]。本研究结果显示,对于具有不同交配系统特性和传粉者丰度的3种植物而言,其资源分配模式表现出差异性。从繁殖分配比例来看,自花授粉植物湿生扁蕾的繁殖分配比例显著高于另外两种异花授粉植物,表明更多的资源投资到了繁殖器官。对于自花授粉植物而言,虽然相对较低的交配代价可以降低花部吸引结构的投资,但自交的“受精保障”需要更多的资源投入以保障胚珠发育和种子生成,这也有可能是因为植物的生长中心向繁殖器官转移的结果。对于两种异花授粉植物而言,低昆虫访问频率的线叶龙胆相对高昆虫访问频率的祁连獐牙菜具有更高的繁殖分配比例,这可能是由于植物对传粉者的依赖导致低昆虫访问植物需要更多的花部器官投资来吸引昆虫,从而增大了其繁殖器官的投资比例,线叶龙胆相对祁连獐牙菜具有更大的花结构也是对该理论的支持[33]。
分析个体大小依赖的繁殖策略是理解植物繁殖资源分配的第一步[34]。众多研究表明,植物繁殖器官的绝对投入量与植株个体大小正相关[3,22],即随着植株个体增大,其繁殖分配比例也会增多。本研究中,3种植物虽然在交配系统特征上存在差异,但是在繁殖器官和个体大小之间都存在显著正相关关系,表明交配系统类型并不会影响这种相关关系,而这种同时增加对营养器官和繁殖器官资源投入的策略有利于适应变化的环境,并很有可能作为一种进化稳定策略在植物界普遍存在。从繁殖分配比例角度分析,繁殖分配和个体大小之间可能存在正相关、负相关和不相关3种相关关系[22,35],但在青藏高原地区,已有的研究证实多数植物的繁殖分配与植株个体之间存在负相关关系[3-4,22,36],这是因为青藏高原地区恶劣的自然环境会增加较大个体植株的繁殖代价,进而降低繁殖分配以提高其生存适合度[37]。但是,也有数据说明青藏高原地区植物的繁殖分配与植物个体大小之间不存在显著相关关系[38],对于这种现象的解释科学界倾向于认为青藏高原地区环境复杂,而这种多变复杂的环境造成繁殖分配和个体大小之间关系的不一致,进而总体上表现为二者无显著相关关系[39]。本研究结果显示,3种不同交配系统类型的植物的繁殖分配具有差异,但繁殖分配与个体大小之间关系却不相同。自花授粉植物湿生扁蕾的繁殖分配比例与其个体大小之间不存在显著的相关关系,这可能与湿生扁蕾处于异质化的生境有关[4],然而考虑到湿生扁蕾为自花授粉植物,这种自交的生活史特点降低了其繁殖代价,因而植物对繁殖器官的资源投资比例对个体大小的依赖性减小,亦即植物个体大小不再是限制植物繁殖的决定因素,所以表现为湿生扁蕾的繁殖分配与其个体大小之间无显著的相关关系。对于异花授粉植物而言,由于其繁殖成功需要传粉媒介参与,较大个体的植株固然可以通过增加较大的繁殖投入来增加其对传粉昆虫的吸引,但由此不可避免的增大了其为应对多变恶劣环境而增加的繁殖代价,相应地增加了繁殖支持结构的资源投资,从而降低了繁殖分配比例[22,39]。
4 结论
3种植物的P/O存在显著差异,而这种差异与不同物种的交配系统特点有关。3个物种的繁殖分配比例不同,自交物种的繁殖分配比例显著高于异交物种。在个体大小依赖效应上,3个物种的繁殖投资与植株个体大小存在显著正相关关系,但是繁殖分配比例却存在差异,表现在自交物种的繁殖分配比例与植株个体大小不存在显著相关关系,而异交物种的繁殖分配比例与植株个体大小存在显著负相关关系。本研究结果证实,植物交配系统特点会影响到繁殖分配特性,而植物的交配系统特点所带来的不同的繁殖代价可能是造成上述结果的主要原因。
References:
[1] Cheplick G P.Life history trade-offs inAmphibromusscabrivalvis(Poaceae):Allocation to clonal growth,storage,and cleistogamous reproduction.American Journal of Botany,1995,82(5):621-629.
[2] 张大勇.理论生态学.北京:高等教育出版社,2000:332. Zhang D Y.Theoretical Study of Ecology.Beijing:Higher Education Press,2000:332.(in Chinese)
[3] 王一峰,靳洁,侯宏红,赵博,曹家豪,李筱姣.川西风毛菊花期资源分配随海拔的变化.植物生态学报,2015(9):901-908. Wang Y F,Jin J,Hou H H,Zhao B,Cao J H,Li X J.Changes in flowering resource allocation ofSaussureadzeurensiswithelevations.Chinese Journal of Plant Ecology,2015(9):901-908.(in Chinese)
[4] 侯勤正,叶广继,马小兵,苏雪,张世虎,孙坤.青藏高原不同生境下湿生扁蕾(Gentianopsispaludosa)个体大小依赖的繁殖分配.生态学报,2016,36(9):2686-2694. Hou Q Z,Ye G J,Ma X B,Su X,Zhang S H,Sun K.Size-dependent reproductive allocation ofGentianopsispaludosain different habitats of the Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2016,36(9):2686-2694.(in Chinese)
[5] 梁坤伦,张洪荣,张丽静,代万安,李晓忠,周志宇,刘雪云,王瑞.紫穗槐表型可塑性及植株资源分配对高寒生境的响应.草业科学,2012,29(3):440-446. Liang K L,Zhang H R,Zhang L J,Dai W A,Li X Z,Zhou Z Y,Liu X Y,Wang R.Response of phenotypic plasticity and plant resource allocation ofAmorphafruiticosato alpine habitat.Pratacultural Science,2012,29(3):440-446.(in Chinese)
[6] 赵宏魁,马真,张春辉,雷占兰,姚步青,周华坤.种植密度和施氮水平对燕麦生物量分配的影响.草业科学,2016,33(2):249-258. Zhao H K,Ma Z,Zhang C H,Lei Z L,Yao B Q,Zhou H K.The reproductive allocation ofAvenasativaunder different planting densities and nitrogen addition treatments.Pratacultural Science,2016,33(2):249-258.(in Chinese)
[7] 张大勇.植物生活史进化繁殖生态学.北京:科学出版社,2004:421. Zhang D Y.Plant Reproductive Ecology.Beijing:Science Press,2004:421.(in Chinese)
[8] Zhang D Y.Resource allocation and the evolution of self-fertilization in plants.American Naturalist,2000,155(2):187-199.
[9] 张大勇,姜新华.植物交配系统的进化、资源分配对策与遗传多样性.生态学报,2001,25(2):130-143. Zhang D Y,Jiang X H.Mating system evolution,resource allocation,and genetic diversity in plants.Acta Ecologica Sinica,2001,25(2):130-143.(in Chinese)
[10] 赵志刚,杜国祯.毛茛科植物交配系统的特征与花期资源分配对策.兰州大学学报:自然科学版,2003,39(5):70-74. Zhao Z G,Du G Z.Characteristics of the mating system and strategies for resource allocation in Ranunculaceae.Journal of Lanzhou University:Natural Sciences,2003,39(5):70-74.(in Chinese)
[11] Mepeek M A,Miller T E.Evolution biology and community ecology.Ecology,1996,77:1319-1320.
[12] Silvertown J W,Franco M,Harper J L.Plant Life Histories:Ecology,Phylogeny and Evolution.Cambridge:Cambridge University Press,1997:331.
[13] Cruden R W.Pollen-Ovule ratios:A conservative indicator of breeding systems in flowering plants.Evolution,1977,31(1):32-46.
[14] Charlesworth D,Charlesworth B.The effect of investment in attractive structures on allocation to male and female functions in plants.Evolution,1987,41(5):948-968.
[15] Brunet J.Sex allocation in hermaphroditic plants.Trends in Ecology & Evolution,1992,7(3):79-84.
[16] 赵志刚,杜国祯,刘左军.雌雄同花植物的性分配.生态学报,2005,25(10):2725-2733. Zhao Z G,Du G Z,Liu Z J.Sex allocation in hermaphroditic plants.Acta Ecologica Sinica,2005,25(10):2725-2733.(in Chinese)
[17] Cruden R W,Lyon D L.Patterns of biomass allocation to male and female functions in plants with different mating systems.Oecologia,1985,66(2):299-306.
[18] Lloyd D G.Allocations to pollen,seeds and pollination mechanisms in self-fertilizing plants.Functional Ecology,1987,1(2):83-89.
[19] 李莹,曾晓琳,游明鸿,刘金平,蔡捡.5种川西北沙化地草本植物生态适应策略的差异性.草业科学,2016,33(5):843-850. Li Y,Zeng X L,You M H,Liu J P,Cai J.Differences of ecological adaptation strategies of 5 herbs from the desertified grassland in the northwest Sichuan.Pratacultural Science,2016,33(5):843-850.(in Chinese)
[20] Weiner J.Allocation,plasticity and allometry in plants.Perspectives in Plant Ecology,2004,6(4):207-215.
[21] Weiner J,Campbell L G,Pino J,Echarte L.The allometry of reproduction within plant populations.Journal of Ecology,2009,97(6):1220-1233.
[22] 王赟,胡莉娟,段元文,杨永平.岩白菜(虎耳草科)不同海拔居群的繁殖分配.云南植物研究,2010,32(3):270-280. Wang Y,Hu L J,Duan Y W,Yang Y P.Altitudinal variations in reproductive allocation ofBergeniapurpurascens(Saxifragaceae).Acta Phytotaxonomica Sinica,2010,32(3):270-280.(in Chinese)
[23] Jongejans E,Kroon H D,Berendse F.The interplay between shifts in biomass allocation and costs of reproduction in four grassland perennials under simulated successional change.Oecologia,2006,147(2):369-78.
[24] 彭德力,张志强,牛洋,杨扬,宋波,孙航,李志敏.高山植物繁殖策略的研究进展.生物多样性,2012,20(3):286-299. Peng D L,Zhang Z Q,Niu Y,Yang Y,Song B,Sun H,Li Z M.Advances in the studies of reproductive strategies of alpine plants.Biodiversity Science,2012,20(3):286-299.(in Chinese)
[25] 郭小伟,杜岩功,林丽,李以康,张法伟,李茜,刘淑丽,欧阳经政,曹广民.青藏高原北缘3种高寒草地的CH4、CO2和N2O通量特征的初步研究.草业科学,2016,33(1):27-37. Guo X W,Du Y G,Lin L,Li Y K,Zhang F W,Li Q,Liu S L,Ouyang J Z,Cao G M.CH4,CO2and N2O flux among three types of alpine meadow in the north regions of Qinghai-Tibetan Plateau.Pratacultural Science,2016,33(1):27-37.(in Chinese)
[26] Duan Y W,Dafni A,Hou Q Z,He Y P,Liu J Q.Delayed selfing in an Alpine BiennialGentianopsispaludosa(Gentianaceae) in the Qinghai-Tibetan Plateau.Journal of Integrative Plant Biology,2010,52(6):593-599.
[27] 段元文,刘建全.青藏高原特有植物祁连獐牙菜(龙胆科)的花综合征与虫媒传粉.植物分类学报,2003,41(5):465-474. Duan Y W,Liu J Q.Floral syndrome and insect pollination of the Qinghai-Tibet Plateau endemicSwertiaprzewalskii(Gentianaceae).Acta Phytotaxonomica Sinica,2003,41(5):465-474.(in Chinese)
[28] 侯勤正,段元文,司庆文,杨慧玲.青藏高原晚期开花植物线叶龙胆的传粉生态学.植物生态学报,2009,33(6):1156-1164. Hou Q Z,Duan Y W,Si Q W,Yang H L.Pollination ecology ofGentianalawrenceiVAR.Farreri,a Late-flowering Qinghai-Tibet plateau species.Chinese Journal of Plant Ecology,2009,33(6):1156-1164.(in Chinese)
[29] Sultan S E.Phenotypic plasticity for plant development,function and life history.Trends in Plant Science,2000,5(12):537-542.
[30] Sultan S E.Promising directions in plant phenotypic plasticity.Perspectives in Plant Ecology Evolution & Systematics,2004,6(4):227-233.
[31] De Jager J C L,Joubert L,Venter H J T,Venter A M.Variation in pollen-ovule ratio of southern African Periplocoideae (Apocynaceae):Evolutionary implications.South African Journal of Botany,2015,98:176.
[32] Sharma N,Kaul Vkoul A K.Pattern of resource allocation of six plantago species with different breeding systems.Journal of Plant Research,1999,112(1):1-5.
[33] Ho T N,Liu S W.A Worldwide Monograph ofGentiana.Beijing:Science Press,2001.
[34] Weiner J,Rosenmeier L,Massoni E S,Vera J N,Plaza E H,Sebastià M T.Is reproductive allocation inSeneciovulgarisplastic.Botany-botanique,2009,87(5):475-481.
[35] Hautier Y,Randin C F,Stöcklin J,Guisan A.Changes in reproductive investment with altitude in an alpine plant.Journal of Plant Ecology,2009,2(3):125-134.
[36] 何亚平,段元文,费世民,刘建全,杨慧玲.青藏高原天山报春高寒湿地种群的花期资源分配.应用与环境生物学报,2008,14(2):180-186. He Y P,Duan Y W,Fei S M,Liu J Q,Yang H L.Resource allocation ofPrimulanutanspopulation in the alpine wet land of the east Qinghai-Tibetan Plateau,China.Chinese Journal of Applied and Environmental Biology,2008,14(2):180-186.(in Chinese)
[37] Reekie E G.An explanation for size-dependent reproductive allocation inPlantagomajor.Canadian Journal of Botany,2011,76(1):43-50.
[38] 陈学林,梁艳,齐威,苏梅,杜国祯.一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究.草业学报,2009,18(5):58-66. Chen X L,Liang Y,Qi W,Su M,Du G Z.Studies on reproductive allocation,floral size and its trade-off with floral number of annualGentiana.Acta Prataculturae Sinica,2009,18(5):58-66.(in Chinese)
[39] 赵志刚,杜国祯,任青吉.5种毛茛科植物个体大小依赖的繁殖分配和性分配.植物生态学报,2004,28(1):9-16. Zhao Z G,Du G Z,Ren Q J.Size-dependent reproduction and sex allocation in five species of Ranunculaceae.Acta Ecologica Sinica,2004,28(1):9-16.(in Chinese)
(责任编辑 武艳培)
Reproductive allocation of three Gentianaceae species
Hou Qin-zheng, Wen Jing, Li Hao-cong, Ren Yu-lan, Sha Jian-hua, Zhao Dong-guang
(Northwest Normal University, Lanzhou 730070, China)
To test the tactics of resource allocation, pollinator abundance, and mating system, the reproductive allocation characters of three species of the family Gentianaceae [Gentianopsispaludosa(selfing),Gentianafarreri(outcrossing with high insect visit frequencies), andSwertiaprzewalskii(outcrossing with low insect visit frequencies)], were tested at the same habitat in the eastern Qinghai-Tibetan Plateau. The results showed that the P/O ratios differ significantly among these three species. The P/O ratio ofG.paludosawas significantly lower than that of the other two cross-fertilized species, and the P/O ratio ofS.przewalskiiwas significantly lower than that ofG.farreri. Reproductive allocation differed significantly among the three species.G.paludosahad a higher reproductive allocation than that in the two cross-fertilized species, and the reproductive allocation ofG.farreriwas higher than that inS.przewalskii. The reproductive allocations of the two cross-fertilized speciesG.farreriandS.przewalskiiwere negatively correlated with their respective individual sizes, whereasG.paludosadid not reveal any relationship between these two parameters. The results indicated that mating system and pollinator abundance might influence resource allocation characteristics. Although the reproductive biomass of the three species was positively correlated with their individual size, the reproductive allocation of the self-fertilized species was significantly higher than that in the two cross-fertilized species. In addition, its high reproductive allocation was not affected by individual size. It appeared that the high reproductive allocation might be caused by reproductive assurance, which in turn might reduce the reproductive cost in the self-fertilized species.
resource allocation; mating system; reproductive allocation; pollinator abundance; self-pollination; cross-pallination
Hou Qin-zheng E-mail:hou_qzh@163.com
10.11829/j.issn.1001-0629.2016-0343
侯勤正,文静,李昊聪,任昱兰,沙建华,赵东光.3种龙胆科植物的资源分配对策.草业科学,2017,34(5):1017-1023.
Hou Q Z,Wen J,Li H C,Ren Y L,Sha J H,Zhao D G.Reproductive allocation of three Gentianaceae species.Pratacultural Science,2017,34(5):1017-1023.
2016-06-24 接受日期:2016-11-03
国家自然科学基金(31360044)
侯勤正(1984-),男,山东菏泽人,副教授,博士,主要从事植物生态学研究。E-mail:hou_qzh@163.com
Q945.3
A
1001-0629(2017)05-1017-07
