磷素供给对米槁幼苗光合作用—CO2响应的影响
2017-05-30熊雪刘济明王军才柳嘉佳李佳邓明明
熊雪 刘济明 王军才 柳嘉佳 李佳 邓明明


摘要:[目的]探索不同磷素水平下米槁(Cinnamomum migao H.W. Li)幼苗光合作用对CO2的响应机制,为西南地区推广种植米槁及其科学施用磷肥提供参考依据。[方法]以0.5年生米槁幼苗为对象,设4个磷素(KH2P04)供给浓度(cK为1.000mmol/L,P1、P2和P3处理分别为0.125、0.500和2.000mmol/L)处理,测定各处理米槁幼苗的光合特性,利用直角双曲线模型、非直角双曲线模型及直角双曲线修正(PJG)模型拟合米槁幼苗的光响应曲线和参数。[结果]3种米槁幼苗光响应曲线拟合模型的拟合决定系数R2均在0.9900以上,表达的效力趋势排序为Pn/Ci模型>非直角双曲线模型>直角双曲线模型,以Pn/Ci模型对米槁的光合作用-CO2响应最实用,且不同磷素水平下模型拟合效力均符合此效力趋势。各磷素供给处理中,除胞间CO2浓度(Ci)外,净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tt)的排序均为CK>P3>P2>P1,且P1处理的Pn、Gs和Tr均与其他处理差异显著(P<0.05),表明1.000mmol/L KH2P04处理下米槁幼苗的光合作用效率最高。[结论]Pn/Ci模型最适宜进行米槁幼苗光合作用-CO2响应拟合;施用适量磷素能促进米槁幼苗的光合作用,最适宜米槁幼苗生长的土壤磷含量为1.000mmol/L。
关键词:米槁幼苗;光合作用;C02n向应;双曲线模型;土壤磷含量
中图分类号:S723.7 文献标志码:A 文章编号:2095-1191(2017)11-1983-06
0引言
[研究意义]米槁(Cinnamomum migao H.W.Li)是樟科樟属常绿乔木,主要分布于我国西南地区,其成熟干燥果实大果木姜子是贵州省重点发展的民族特色药材(邱德文等,1993),但目前对米槁的研究主要集中在栽培與病虫害防治(江兴龙等,2005)、果实挥发油和药用成分的生物活性与药理作用(周涛等,2010;张小波等,2011)等方面,对其光合作用方面的研究仅见骆畅等(2016)、王军才等(2016)分别对不同氮素、磷素水平下米槁光响应模型的研究报道。因此,探索不同磷素供给水平下米槁幼苗光合作用对C02的响应机制,对西南地区推广种植米槁及其科学施用磷肥具有重要意义。[前人研究进展]植物的光合作用除受光和土壤水分等影响外,还受CO2浓度和土壤肥力等因素影响。于海秋等(2005)研究指出,磷素是影响植物生长的第二大元素,在光合作用中以二磷酸腺苷(ADP)和三磷酸腺苷(ATP)的形式储存和转运能量。叶子飘和于强(2009)研究认为,植物CO2响应曲线主要用以研究植物净光合速率与CO2间的关系,并根据曲线估算植物C02补偿点(CCP)、CO2饱和点(CSP)、表观羧化效率(ACE)和光呼吸速率(Rp)等光合参数。Han等(2012)研究发现,CO2是植物光合作用的基本原料,也是1,5-二磷酸核酮糖(RuBP)羧化酶的主要底物,直接参与光合作用过程。常用的植物光合作用对CO2响应曲线生化拟合模型有直角双曲线模型(Baly,1935)、Farquhar模型及其修正模型(Voncaemmerer and Farquhar,1981)、Michaelis-Menten模型(Harley et a1.,1992)、指数方程模型和非直角双曲线模型(Yu et a1.,2004)等。但叶子飘(2010)研究指出,双曲线模型和Michaelis-Menten模型本质上类同,实际运用中直角双曲线模型和指数方程模型较常见。王建林等(2005)、叶子飘和高峻(2008)的研究结果均认为,直角双曲线模型和指数方程模型在实际运用中不能准确模拟出植物光合作用C02饱和点的缺陷。叶子飘和于强(2009)、李丽霞等(2016)研究提出直角双曲线修正模型(简称Pn/Ci模型)在实际运用中能有效弥补其他模型的缺陷,可直接得出CO2饱和点,较直观地反应正常培养条件下植物C02的响应情况,但该模型在不同磷素条件影响下是否能保持同样的优势尚不清楚。[本研究切入点]米槁结果大小年现象明显,急需通过对米槁幼苗科学施用磷素促进其光合作用,加快育苗进程进而促进米槁人工林发展,以提高果实产量来缓解市场资源匮乏状况,但目前关于米槁幼苗不同磷素水平下光合作用对CO2响应机制的研究鲜见报道。[拟解决的关键问题]采用直角双曲线模型、非直角双曲线模型及POn/Ci模型对不同梯度磷素浓度处理下米槁光合作用对CO2响应情况进行拟合,探讨不同磷营养条件下米槁幼苗光合作用对CO2的响应机制,明确CO2响应过程与磷素含量的关系,为西南地区推广种植米槁及其科学施用磷肥提供参考依据。
1材料与方法
1.1研究地概况
试验于2015年在贵州大学林学院(东经106°40′10″、北纬26°25′26″)苗圃进行。试验地平均海拔1020m,年均气温14.9℃。一年中气候温和,属中亚热带温润季风气候。
1.2试验材料
供试材料为贵州省罗甸县米槁育苗基地培育的120d龄健康、无病害、长势一致的米槁幼苗,于2015年3月底分别移栽至贵州大学林学院苗圃装有石英砂的花盆中,每盆种植3株。
1.3试验方法
1.3.1试验设计 移栽苗采用全营养液(CK)浇灌,石英砂基质培养,缓苗处理1个月后,选取长势一致、健康无病的幼苗进行不同浓度磷素处理,每处理3个重复。供试苗木每3d(上午8:00-9:00)浇1次营养液,每盆每次浇200mL,试验期间适量补水,直至当年10月5日试验结束。
供试营养液参照Jarkko和Toini(2001)的霍格兰营养液及王军才等(2016)的营养液配方进行配制。其中,通过调节KH2P04浓度形成4个磷素养分梯度,即0.125mmol/L(P1处理)、0.500mmol/L(P2处理)、1.000mmol/L(CK)和2.000mmol/L(P3处理),用KC1调节钾素浓度,再用CaCl2调节离子平衡,其他成分含量相同。
1.3.2指标测定 于2015年9月米槁生长最活跃时段,选择晴朗天气,于上午9:00~11:30,采用LI-6400XT便携式光合作用测定系统(美国LICOR公司)测定各处理米槁叶片的CO2响应曲线,每处理3个重复,每重复测定3片叶片。
叶片活化条件(活化20min):C02浓度(400.00±5.00)pmol/mol,叶片温度(25.0±0.5)℃,气体流速500gmol/s,光强1400gmol/m2·s。在上述条件下,用LI-6400XT的外源CO2注入系统设定叶室C02浓度梯度为1800.00、1500.00、1200.00、1000.00、800.00、600.00、400.00、300.00、200.00、100.00、80.00、50.00和20.00gmol/mol,采用自动程序测量,每个设置值停留4min,自动记录净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)等参数,3次重复,取平均值。
1.4统计分析
试验数据采用Excel 2013及SPSS 22.0进行整理,用非线性回归功能对光合-CO2响应曲线分别进行直角双曲线模型、非直角双曲線模型和Pn/Co模型拟合比较,以非线性最小二乘法的Marquardt迭代原理求解模型参数,并采用Dancans多重比较法进行差异显著性检验。
2结果与分析
2.13种双曲线模型下米槁幼苗的光合-CO2响应参数拟合比较分析
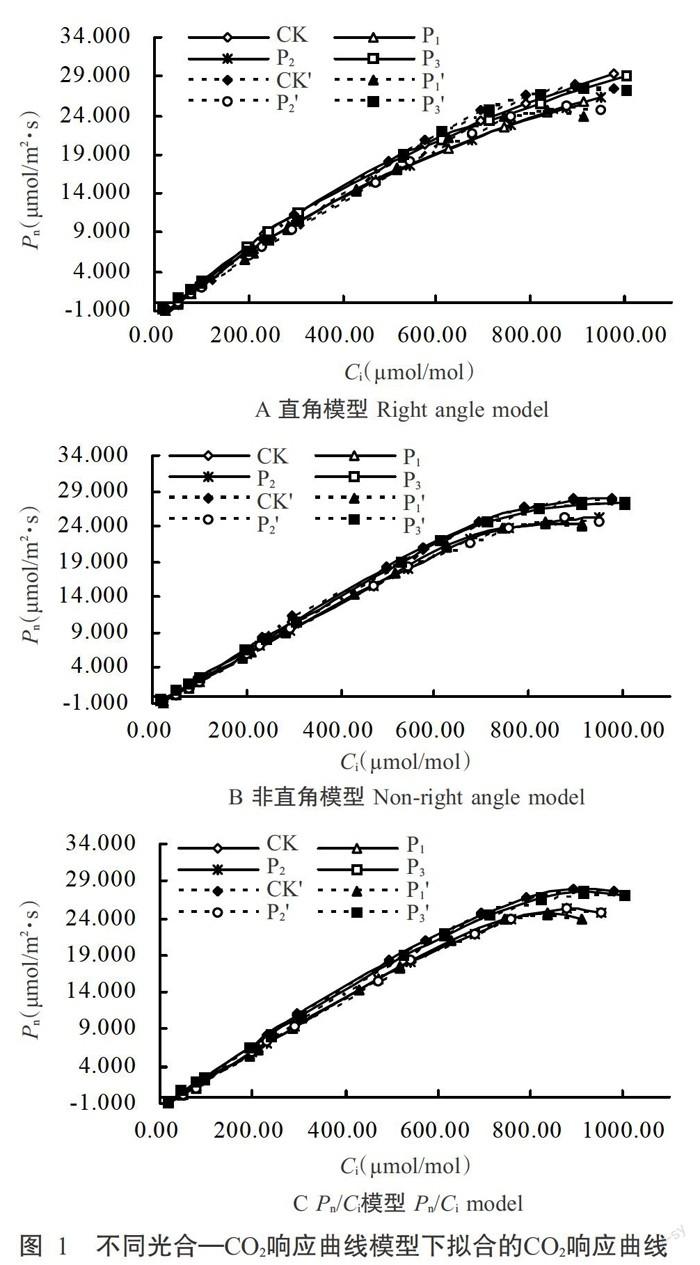
从图1可看出,3种光合C02响应曲线模型的米槁幼苗叶片Pn实测值在低Ci阶段(Ci< 200.00 gmol/m01)呈近似直线状快速上升,之后随着G的升高上升速度减缓;Ci达饱和值(P1处理为836.33μmol/mol,P2处理为878.20μmol/mol,CK为893.65μmol/mol,P3处理为914.23μmol/mol)时,Pn出现最大值(P1处理为24.599μmol/m2·s,P2处理为25.357μmol/m2·s,CK为27.980μmol/m2·s,P3处理为27.383μmol/m2·s);P3处理下Cli饱和值最大(914.23μmol/mol),但Pn仅27.383μmol/m2·s,而CK的G在饱和状态下(893.65μmol/mol)Pn最高(27.980μmol/m2·s)。说明施用磷素能有效提高米槁幼苗叶片CO2利用率,从而提高光合作用能力,但施用磷量过大,Ci饱和点也增大,光合作用能力有所降低,即低磷和高磷条件对米槁幼苗叶片光合作用均有不同程度的抑制性。
比较图1和表1结果可知,3种光合-CO2响应曲线模型均能模拟出米槁幼苗叶片Pn对Ci的大致变化趋势,即Pn皆随Ci的增大而上升,当Ci达到一定值后,Pn也随磷素处理浓度的增加而升高,其中CK的pn最高。3种拟合模型的拟合决定系数(R2)均在0.9900以上,但通过直角双曲线模型拟合,各磷素处理下CSP、Pnmax和CCP均大于实测值(表1),且Pn不断上升(图1-A),与Pn实测值差距较大;非直角双曲线模型中Pn随着Ci的增加出现不明显的近似饱和现象(图1-B),也无明显下降趋势,且各磷素处理下该模型拟合参数CSP和CCP均小于实测值,Pnmax略大于实测值(表1);Pn/Ci摸型拟合参数与实测值最接近(表1),且出现Pn随Ci增大到一定值后略有下降的现象(图1-C)。从拟合准确性看,3种模型表达的效力趋势排序为Pn/Ci摸型>非直角双曲线模型>直角双曲线模型,且各磷素处理下模型拟合效力均符合此趋势,说明在不同磷素浓度处理下Pn/Ci模型比其他模型更具优势。
通过3种模型拟合参数与实测值的对比,确定以Pn/Ci模型光合参数拟合值探讨磷素处理下各光合参数的变化情况。由表1可知,拟合值中磷素浓度的增加能提高米槁ACE和Pnmax,但呈波动变化趋势;而Rp值恰好相反,呈波浪状降低。随着磷素浓度的增加,CSP含量增加,P3处理下CSP含量最高(932.037pmol/mol2);而CCP则相反,随着磷素浓度的增加而减小,P3处理下CCP含量最低(36.683 gmol/mol2)。说明其他条件相同情况下,施加磷素能提高米槁幼苗对胞间CO2的利用率。
2.2不同磷素浓度水平下米槁幼苗光合作用对CO2的响应
2.2.1Ci对CO2浓度变化的响应如图2所示,在低CO2浓度阶段(CO2浓度<200.00μnol/mol),米槁幼苗叶片G随着CO2浓度的增加呈直线上升趋势,其后Ci增加速度有所减缓;在CO2浓度/>200.00μmol/mol后,随施磷浓度增大,同-CO2浓度下各处理Ci存在差异,其排序为P3>CK>P2>P1,即增施磷量可增强米槁幼苗叶片对CO2的利用能力。随着施磷浓度和CO2浓度的增大,米槁幼苗叶片利用胞问CO2能力也逐渐增强,其中P3处理下的米槁对CO2利用能力增幅最大,其次为CK,但处理问差异不明显,且P3浓度下的Ci与其他浓度下Ci的差距随着CO2浓度的增大呈不断缩小趋势,说明增施磷量能提高米槁对CO2的利用率,但提高效果不明显。
2.2.2Gs对CO2浓度变化的响应 从图3可看出,在PAR=1400.00μmol/m2·mol时,Gs随着CO2浓度的升高先快速下降而后缓慢下降,说明低浓度CO2促进气孔张开,高浓度CO2使气孔关闭;适当增加磷量能提升Gs,P3处理下Gs最大,说明增施磷量能减缓CO2浓度上升对Gs的负面影响,但未改变Gs的整体下降趋势。
2.2.3Tr对CO2浓度变化的响应 从图4可看出,Z的变化趋势与Gs相似,皆随着CO2浓度的升高急速下降,下降速率先快后缓;当CO2浓度不断增大时,米槁叶片气孔收缩,明显抑制Tr;在CO2浓度达到一定值后,增加磷量可减缓Tr降幅,但未改变Tr的总体下降趋势。
2.3不同磷素水平对米槁幼苗气体交换性指标的影响
由表2可知,各磷素浓度处理中,除Ci外,Pn、Gs及Tr的排序均为CK>P3>P2>P1,即CK(1.000 mmol/LKH~P04)處理下的米槁幼苗光合作用效率最高,P1处理下的米槁幼苗光合作用效率最低,且P1处理的Pn、Gs和Tr均极显著(P<0.01)低于其他处理,说明低磷条件下米槁幼苗的光合作用明显受到抑制,气体交换较慢,适当增加磷量能促进米槁幼苗叶片的光合作用,但促进作用不显著,而磷量过度会抑制米槁幼苗叶片的光合作用。
3讨论
植物光合作用对CO2的响应曲线通常在该植物的饱和光强下进行测定(Yu et a1.,2004),用以研究植物Pn与C2间的关系,并以此估算植物相关光合参数。叶子飘(2010)的研究曾提出直角双曲线模型与非直角双曲线模型能拟合米槁Pn-CO2的大致变化规律,但直角双曲线模型是无极点的渐近线,无法模拟出CO2饱和现象,且其模拟值远大于实测值,严重影响其适用性及准确度,本研究也获得了相似的研究结果。此外,非直角双曲线模型曾被用于冬小麦(Yu et a1.,2004)和小蓬竹(李丽霞等,2016)等物种Pn-CO2的响应研究,但无法求取CSP的解析解。本研究中,非直角双曲线模型中Pn随着Ci的增加出现不明显的近似饱和现象,但无明显下降趋势,不能准确表达米槁Pn-CO2的响应情况,且拟合值中CSP和CCP均小于实测值,而Pnmax却大于实测值,无法表达植物在胁迫条件下光合作用对CO2的响应情况,与Yu等(2004)、李丽霞等(2016)的研究结果相似。叶子飘和高峻(2008)、吴芹等(2013)研究认为,目前应用最广的Pn/Ci模型可弥补以上各研究存在的缺陷,能较准确地表达Pn-Ci间的关系。本研究获得同样的结果,所用3种拟合模型中以Pn/Ci模型最实用。
结合拟合参数及实测值,米槁幼苗叶片光合作用的Pn对CO2响应有3个明显反应阶段,即Pn随Ci增大(Ci<200.00μmol/m01)呈近似直线上升阶段、Pn随Ci增大呈缓慢上升阶段(200.00μmol/mol≤Ci
