温度胁迫对三七光合特性及生理指标的影响
2017-05-30左应梅张金渝杨天梅杨绍兵杨维泽李纪潮杨美权许宗亮简邦丽
左应梅 张金渝 杨天梅 杨绍兵 杨维泽 李纪潮 杨美权 许宗亮 简邦丽
摘要:【目的】研究三七对温度胁迫的生理响应,揭示三七的耐寒和耐热机制,为三七抗逆品种的选育、区域化种植和栽培管理提供理论依据。【方法】以盆栽生长第2年的三七植株为试验材料,利用人工气候培养箱进行5、10、25(对照)和35℃温度处理,持续处理48 h后测定三七叶片的光合特性及叶绿素含量、保护酶活性等生理指标,并分析其变化规律。【结果】在5~25℃的温度条件下,三七叶片的叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素[Chl(a+b)]含量呈先升后降的变化趋势,与25℃处理相比,5℃处理分别降低21.91%、23.30%和22.78%,10℃处理分别提高51.69%、52.43%和51.96%,差异均达显著水平(P<0.05,下同);与25℃处理相比,35℃高温条件下,Chla、Chlb和Chl(a+b)含量及Chl(a/b)分别降低14.61%、6.80%、11.74%和8.10%,差异均达显著水平。低温胁迫下,三七叶片的最大净光合速率(Pmax)、表观量子效率(a)、光饱和点(LSP)、净光合速率(Pn)、气孔导度(G)和蒸腾速率(Tr)均显著下降,胞间二氧化碳浓度(Ci)显著升高,光补偿点(LCP)呈先升后降的变化趋势;高温胁迫下,三七叶片的Pmax、LSP、Pn、Gs和Tr均显著下降,Ci、a和LCP则呈上升趋势。在低、高温胁迫条件下,三七叶片的过氧化氢酶(CAT)活性显著降低,过氧化物酶(POD)和超氧化物歧化酶(SOD)活性总体上显著升高;丙二醛(MDA)含量变化不明显。低温胁迫下三七叶片的可溶性蛋白含量显著升高。【结论】低、高温胁迫均显著抑制三七的光合能力。低温胁迫下,三七主要通过提高POD和SOD活性,增加可溶性蛋白和叶绿素含量来增强其抗冷性;高温胁迫下,三七主要通过提高POD和SOD活性来增强其抗热性。
关键词:三七;温度胁迫;光合特性;生理特性
中图分类号:S567.211 文献标志码:A 文章编号:2095-1191(2017)12-2145-07
0引言
【研究意义】三七[Panax notoginseng(Burk.)F.H.Chen]为五加科人参属多年生药用植物,是我国传统常用名贵药材,也是云南省最具特色的药用生物资源。三七以其干燥后的根及茎叶入药,具有抗炎、止血、抗衰老等药理活性(王莹等,2015)。三七对其生长的环境温度要求严格,适宜种植在夏凉冬暖,热量充沛,年温差变化较小,年均温15.8~19.3℃的低纬度区域。云南省地处低纬度高原山地,地形条件复杂多变,受季风气候影响明显,极端或异常天气频发,对当地三七的生长造成较大影响。此外,三七连作障碍严重,轮作需间隔10~20年,导致主产区的种植场地越来越少,三七种植带逐渐北移,但北移后三七的生长易受冬春季极端低温和持续低温天气的影响。同时,由于三七种植过程中病害发生较严重,常采用避雨栽培以减少病虫害发生,但该种植模式易造成棚内温度过高,对三七的生长发育、产量及品质产生不良影响。因此,深入研究温度对三七光合特性、生理指标及抗逆性的影响,可为三七区域化种植、栽培管理和抗逆性品种的选育提供科学参考依据。【前人研究进展】低温或高温逆境将导致植物体内的活性氧代谢失调,发生氧化胁迫,引起细胞膜脂过氧化加剧、蛋白质变性及核苷酸损伤等,胁迫危害加剧可致使细胞死亡(Kuk et al.,2003),而植物可通过调节自身抗氧化系统对温度胁迫作出适应性响应。同时,低温或高温胁迫可破坏叶片光合机构,使叶绿素合成受阻,光合电子传递链受限,影响碳同化所需能量,导致光合效率下降(武辉等,2014;李孟洋等,2015;许娟等,2016)。高温胁迫方面,许桂芳和张朝阳(2009)研究表明,随着高温胁迫的加剧,4种珍珠菜属植物叶片的质膜透性上升,超氧化物歧化酶(SOD)和过氧化物酶(POD)活性先升后降,叶绿素、可溶性蛋白和脯氨酸含量增加;任飞等(2012)研究表明,高温胁迫处理24 h,日本无刺野蔷薇和七姊妹叶片中的SOD活性比中国无刺野蔷薇和普通月季显著提高,4种供试蔷薇植物叶片的POD活性均比对照(常温)低;李孟洋等(2015)研究表明,随着高温胁迫时间的延长,不同产地茅苍术叶片的净光合速率(Pn)、氣孔导度(Gs)、胞间CO2浓度(G)和蒸腾速率(Tr)均呈下降趋势,且叶片SOD、POD和过氧化氢酶(CAT)活性及丙二醛(MDA)含量均随胁迫时间的延长呈上升趋势。低温胁迫方面,武辉等(2014)研究表明,低温胁迫下棉花幼苗叶片的Pn、Gs和Ci等指标均小幅度降低,胁迫解除后恢复正常,但随着胁迫强度增加,各指标变化幅度显著,胁迫后恢复能力降低,同时,低温胁迫下叶片对光辐射的利用能力降低,且胁迫加强,叶片最大净光合速率(Pmax)、表观量子率(a)及光饱和点(LSP)急剧下降;许娟等(2016)研究表明,低温胁迫后马铃薯叶片的CAT、POD和SOD活性及MDA含量上升,且可溶性糖、可溶性蛋白和脯氨酸含量整体均呈上升趋势,但随着胁迫时间的延长,各生理指标变化表现出不同趋势;陈奇等(2017)研究白菜型油菜在低温胁迫下叶片光合特性与内源激素的响应对其抗寒性的影响,结果表明,低温胁迫均降低了Pn和Gs,且抗寒性弱的品种降幅较大。【本研究切入点】目前,关于温度胁迫对三七光合特性及生理指标影响的研究报道极少。【拟解决的关键问题】以生长第2年的三七植株为试验材料,研究持续低、高温胁迫对三七叶片光合特性及生理指标的影响,揭示三七的耐寒和耐热机制,为三七抗逆品种的选育及其区域化种植和栽培管理提供理论依据。
1材料与方法
1.1试验材料
试验用三七为1年生三七苗,由昆明市禄劝县翠华镇种植农户提供,以生长第2年的三七植株为试验材料。
试验用土壤为石灰岩母质发育的山地红壤,其基础养分:有机质36.57 g/kg,全氮2.38 g/kg、全磷0.75 g/kg,全钾14.15 g/kg,速效氮195.30 mg/kg、速效磷20.25 mg/kg、速效钾272.80 mg/kg,pH 5.90~6.27。
1.2试验方法
2014年1月15日~5月30日在云南省农业科学院药用植物研究所小哨试验基地进行试验。1月15日,将大小一致、来源相同、无破损及病害的1年生三七苗,栽人到上口径21 cm,下口径14 cm,高19 cm的花盆中,基质为壤土:农家肥=2:1,每盆3.5kg,每盆种4株,农艺管理措施一致。
5月22日采用智能人工气候箱(宁波江南仪器厂制造)进行试验。设25(对照)、5、10和35℃共4个温度处理,光照强度3500 lX,相对湿度60%,每处理3次重复;各温度梯度持续处理48 h后,测定三七叶片的光合特性,并将测过光合特性的叶片立刻用蒸馏水冲洗干净后于-80℃保存,进行各项生理指标测定。
1.3测定项目及方法
1.3.1光合特性指标测定 采用LI-6400便携式光合测定系统(LI-COR,USA)测定光合一光强曲线(Pn-PAR)。在上午9:00~11:30进行测定,连续测定3 d,叶室温度设为25℃,CO2浓度设为400 μmol/mol,光强(PAR)设为0、20、50、80、100、150、200、400、600、800、1200和1600μmol/m2·s,共12个梯度,由仪器配置的红蓝光源产生。依据Pn-PAR曲线的初始斜率(PAR<250μmol/m2·s)计算出a。参考Bassman和Zwier(1991)的方法计算Pmax、LSP及光补偿点(LCP)等。此外测定各处理同一光强(PAR=200μmol/m2·s)下的Pn、Tr、Gs和Ci等参数。
1.3.2生理特性指标测定 SOD活性采用NBT(淡蓝四唑)光还原法测定,以抑制NBT光还原反应50%所需酶量为一个酶活性单位(U),活性以u/gFW·min表示,SOD试剂盒购于南京生物工程研究所;POD活性采用愈刨木酚显色法测定,以每分钟OD470变化0.01为一个酶活性单位(U),活性以U/gFW·min表示;CAT活性采用紫外吸收法测定,以1 min内OD240降低0.01为一个酶活性单位(U),活性以U/gFW·min表示。MDA含量采用硫代巴比妥酸(TBA)比色法测定;可溶性蛋白含量采用考马斯亮蓝G-250染色法测定;叶绿素含量采用一定体积的95%酒精提取,用分光光度计测665、649和470 nm 3个波长的吸光度,根据高俊凤(2006)的方法计算叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素[Chl(a+b)]含量及叶绿素a和叶绿素b的比值[Chl(a/b)]。
1.4统计分析
采用Excel 2007进行数据整理和制图,利用JMP7进行差异显著性分析。
2结果与分析
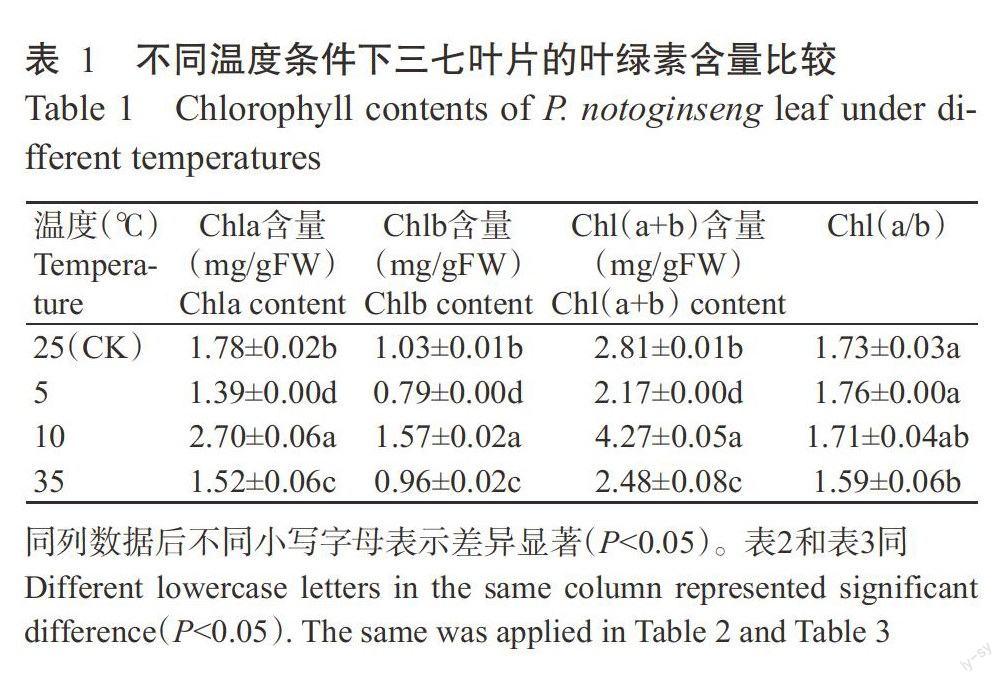
2.1不同温度条件对三七叶片叶绿素含量的影响
由表1可知,在5~25℃的温度条件下,三七叶片的Chla、Chlb和Chl(a+b)含量呈先升后降的变化趋势。与25℃处理的Chla、Chlb和Chl(a+b)含量相比,5℃处理分别降低21.91%、23.30%和22.78%,10℃处理分别提高51.69%、52.43%和51.96%,差异均达显著水平(P<0.05,下同),而Chl(a/b)变化不显著(P>0.05,下同)。在35℃高温条件下,Chla、Chlb、Chl(a+b)含量和Chl(a/b)分别比25℃处理降低14.61%、6.80%、11.74%和8.10%,差异显著。表明三七叶片的叶绿素含量与其耐寒性有关,在一定的低温胁迫下三七可通过增加其叶片叶绿素含量以提高光合作用,超过一定低温胁迫后,叶片叶绿素合成受阻,而高温胁迫下其叶绿素合成同样受阻。
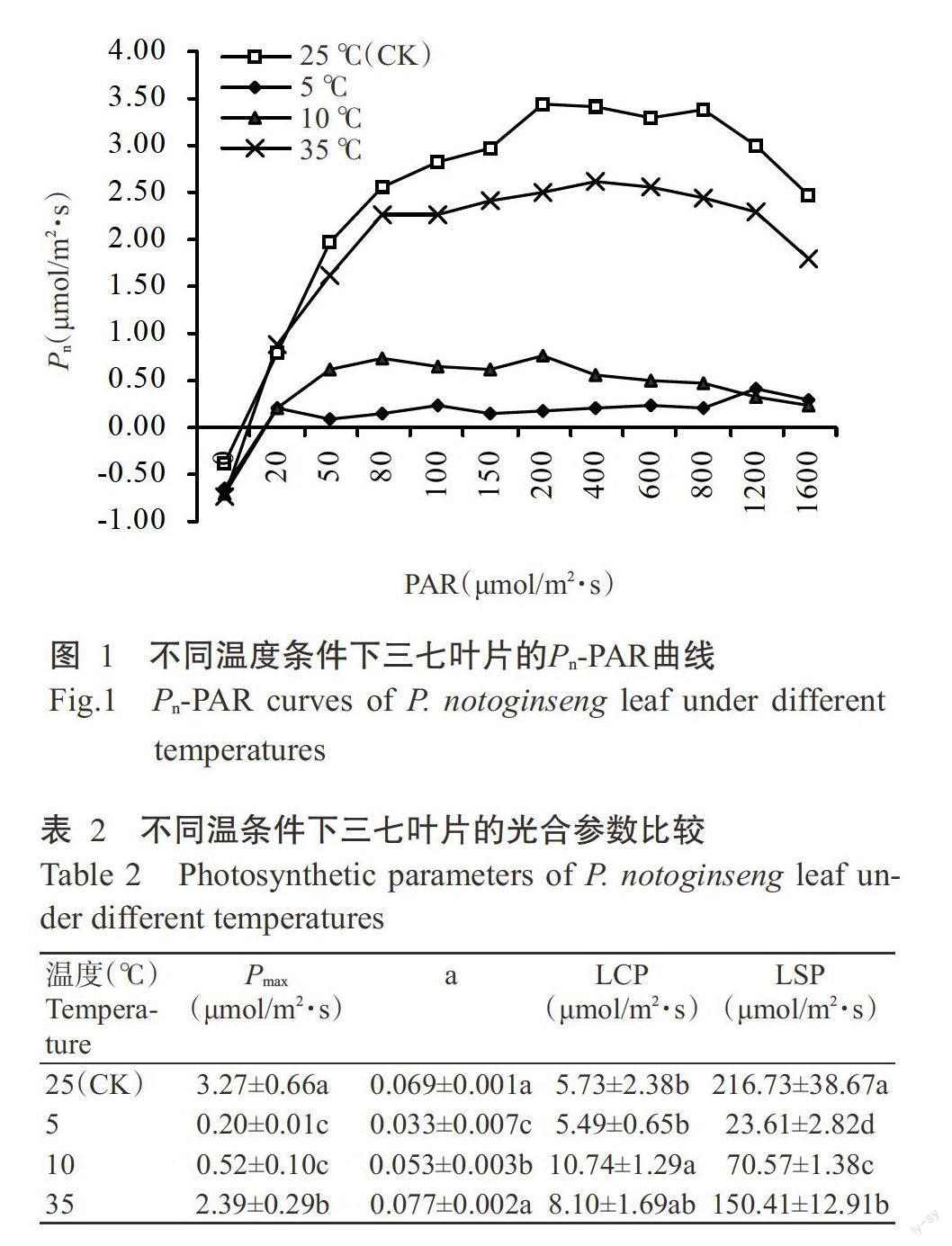
2.2不同温度条件对三七叶片光合特性的影响
2.2.1光响应曲线和光合参数比较 由图1可看出,不同温度条件下,三七叶片的Pn随PAR增强其变化趋势并不一致。10和35℃处理下的Pn变化趋势与25℃基本一致,均随PAR的增强先增加后下降,且PAR相同时,10和35℃处理下的Pn均低于25℃对应值。5℃处理下的Pn随PAR增强变化不明显,表明在该温度条件下三七的光合作用严重受阻。Pn-PAR曲线方程及相应方程的计算结果(表2)表明,温度胁迫导致三七叶片的Pmax、a、LCP和LSP呈不同程度的变化。低、高温胁迫均使三七叶片的Pmax和LSP显著下降,与25℃处理相比,5、10和35℃处理的Pmax分别下降93.88%、84.10%和26.91%,LSP分别下降89.11%、67.44%和30.60%。在低温胁迫下,a呈显著下降趋势,即与25℃处理相比,5和10℃处理的a分别下降52.17%和23.19%。5℃處理的LCP下降4.19%,与25℃处理差异不显著,10℃处理的LCP则显著升高87.43%。在35℃高温条件下,a和LCP的变化均不显著。说明高、低温胁迫均抑制了三七的光合作用,在一定条件下低温胁迫的伤害程度较高温严重;三七在强光下均出现不同程度的光抑制,低、高温胁迫越严重,越容易出现光抑制现象。
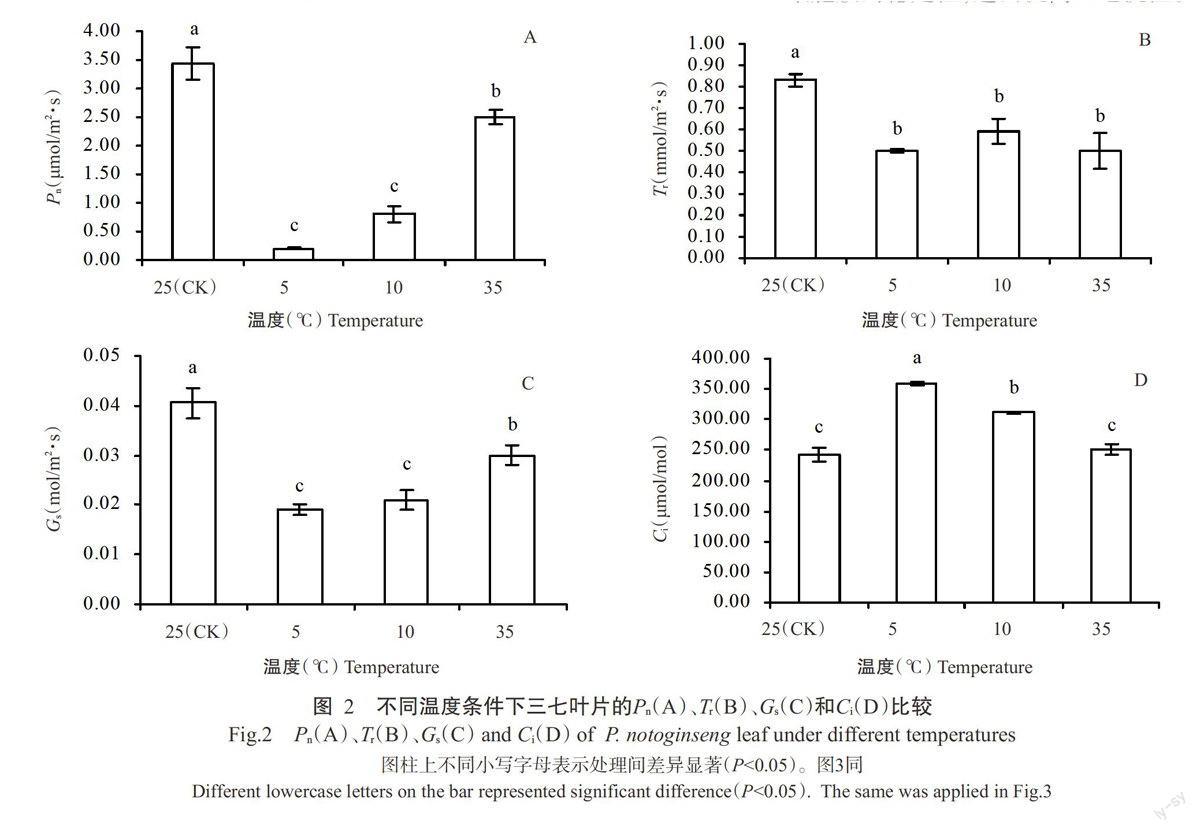
2.2.2 Pn、Tr、Gs和Ci的比较在200μmol/m2·s光照强度下,低、高温胁迫使三七叶片的Pn(图2-A)、Tr(图2-B)和Gs(图2-C)均显著下降,与25℃处理相比,5、10和35℃处理的Pn分别下降94.19%、76.74%和27.33%,Tr分别下降39.76%、28.92%和39.76%,Gs分别下降53.09%、48.15%和25.93%。G(图2-D)则呈相反的变化趋势,与25℃处理相比,5和10℃处理的Ci分别显著升高47.74%和27.98%,35℃处理的G升高3.29%,变化不显著。
2.3不同温度条件对三七叶片保护酶活性和MDA含量的影响
由表3可知,低温和高温胁迫下,三七叶片的CAT活性降低,SOD活性升高。与25℃处理相比,5、10和35℃处理的CAT活性分别显著降低60.71%、46.83%和68.15%,SOD活性分别显著升高18.49%、107.54%和22.36%。5和35℃处理的POD活性较25℃处理显著升高,10℃处理的POD活性较25℃处理略有降低,但差异不显著。低温或高温胁迫下,三七叶片的MDA含量均较25℃处理有所增加,但差异不显著。说明三七的保护酶系统对温度胁迫较敏感,可通过增强其叶片的POD和SOD活性,及时清除活性氧,保护细胞膜的稳定性,进而提高三七抗性。
2.4不同溫度条件对三七叶片可溶性蛋白含量的影响
由图3可看出,不同温度条件下,三七叶片的可溶性蛋白含量均呈不同程度的升高趋势。与25℃处理相比,5和10℃处理的可溶性蛋白含量显著升高22.72%和32.77%;35℃处理的可溶性蛋白含量也有所升高,但与25℃处理差异不显著。说明三七叶片的可溶性蛋白含量与三七的耐低温能力密切相关。
3讨论
3.1不同温度条件对三七叶片叶绿素含量的影响
叶绿素是植物细胞内参与光合作用的重要色素,起着接收光能和转换能量的作用,其含量与光合能力密切相关。目前,关于逆境胁迫下叶绿素含量变化的研究结果尚存在一定差异。任彩虹等(2007)研究指出,高温胁迫下,马铃薯幼苗叶片的叶绿素含量明显下降。金花等(2011)分析比较了7种常绿阔叶植物越冬期间叶绿素含量的变化,结果显示,叶绿素含量随低温胁迫的加剧先降后升。邵怡若等(2013)研究表明,随低温胁迫时间的延长,假连翘幼苗的叶绿素含量先升后降,盐肤木、老鸭嘴和葛藤幼苗的叶绿素含量则表现为持续下降,叶绿素含量的变化趋势与植物类型和胁迫时间密切相关。本研究中,在5-25℃的温度条件下,三七叶片的Chla、Chlb和Chl(a+b)含量呈先升后降的变化趋势;在35℃高温条件下,Chla、Chlb、Chl(a+b)含量和Chl(a/b)较25℃处理显著下降。表明在一定的低温范围内,三七可通过增加其叶片叶绿素含量以维持或提高其光合作用,随着低温胁迫的加剧,三七叶片的叶绿素合成受阻;高温胁迫也可导致三七叶片叶绿素合成受阻。低、高温胁迫使得叶片叶绿素含量明显下降。这是由于叶绿素合成是在叶绿体或前质体中一系列酶的作用下所形成,低、高温胁迫可能通过影响叶绿素合成相关的酶类,如叶绿素合成酶等,进而抑制叶片的叶绿素合成(Tewan and Tripathy,1998);低、高温胁迫引起活性氧积累,加速叶绿素降解,造成叶片叶绿素含量降低(Willekens et al.,1994)。
3.2不同温度条件对三七叶片光合特性的影响
光合作用是植物体的重要代谢之一,可作为判断植物生长状况和抗逆性强弱的重要指标,光补偿点和光饱和点的高低直接反映植物对光环境的适应能力(刘悦秋等,2007)。低温胁迫导致三七叶片对光辐射的利用能力下降,随着胁迫温度降低,叶片的Pmax、a和LSP快速下降,LCP则呈先升后降的变化趋势,与武辉等(2014)在棉花幼苗上的研究结果相似。此外,本研究结果显示,低温胁迫下Pn、Gs和Tr呈显著下降趋势,而Cli呈显著上升趋势,与邵怡若等(2013)在假连翘幼苗上的研究结果相似。据Farqu-har和Sharkey(1982)报道,当Pn和Gi变化方向一致,而Gs下降时,认为Pa的下降主要是由Gs引起。本研究中,随着三七叶片Gs显著下降,Ci则显著上升,说明Pn下降是由非气孔限制占主导因素。由此可知,低温胁迫使三七叶片组织受损,导致光合机构的稳定性能降低、暗反应过程所需的ATP和NADPH供应不足、关键酶活性下降、反应底物的再生速率减慢,对CO2的固定造成阻碍,引起细胞间隙CO2积累,从而导致叶肉细胞光合性能降低(Yu et al.,2002)。在35℃高温胁迫下,三七叶片的Rmax和LSP显著下降,a和LCP则呈上升趋势。说明高温胁迫同样可导致三七叶片对强光环境的适应能力降低。同时,高温条件下,三七叶片的Pn、Gs和Tr均呈显著下降趋势,G呈上升趋势。与杜尧东等(2012)研究得出的高温处理后水稻剑叶的Pn、Gs下降,Ci上升的结果相似。
3.3不同温度条件对三七叶片保护酶活性和膜质过氧化水平的影响
保护酶是植物适应逆境的重要生理基础(马成仓等,2004),其活性高低可反映植物对环境条件适应能力的强弱。保护酶系统主要包括CAT、POD和SOD,其协同作用可降低逆境不良信号对细胞膜脂的攻击(Jebara et al.,2005)。本研究中,低温和高温胁迫下,CAT活性显著降低,POD和SOD活性总体上显著升高。表明三七的保护酶系统对温度胁迫的反应十分敏感,一定程度的低温或高温胁迫下,三七可通过增强其叶片POD和SOD活性,及时清除因胁迫而积累的H2O2及活性氧,以保持细胞内活性氧产生和清除的动态平衡,保护细胞膜稳定性,进而提高其对低、高温逆境的抗性。这可能是三七叶片对低、高温胁迫的一种应激性保护方式,类似结果也表现在其他植物对溫度胁迫后的应激反应中(陈颖等,2012)。POD和SOD可能是三七主要的耐冷、耐热保护酶系统。逆境胁迫下产生的活性氧若不能及时清除,可造成自由基积累,损害细胞膜系统,引起或加剧膜质过氧化作用,产生MDA破坏细胞内活性氧的代谢平衡(Chen,1991)。本研究结果表明,一定程度的低温或高温胁迫下,三七叶片的MDA含量变化不显著,说明三七具有一定的耐冷和耐热性能,可能与其具有的较高POD和SOD活性有关。
3.4不同温度条件对三七叶片可溶性蛋白含量的影响
低温引起植物细胞中蛋白质发生变化,主要表现为可溶性蛋白和酶类的变化及激发信号产生抗寒性蛋白(王毅等,1994)。可溶性蛋白能增加细胞的持水力,并具有较强的亲水性,可通过束缚细胞周围的水分缓解低温条件下原生质因结冰而受到伤害,提高植物体抗寒性(沙伟等,2008)。本研究结果表明,低温胁迫下三七叶片内可溶性蛋白含量显著升高。表明低温可诱导产生新的蛋白,增加可溶性蛋白含量,有利于提高三七的抗寒性,与朱政等(2011)在茶树叶片上的研究结果相似。高温条件会造成正常蛋白合成受限、蛋白分解加剧、细胞内可溶性蛋白含量下降,因此,高温胁迫会引起植物体内可溶性蛋白含量下降(Chaitanya et al.,2001)。但本研究中高温胁迫下三七叶片的可溶性蛋白含量有小幅度上升。说明三七具有一定的耐热性,与鲁歧等(1992)研究的三七起源于第三纪,属古热带残遗植物的结论吻合。
4结论
低温或高温胁迫对三七叶片光合特性及生理特性的影响明显。低、高温胁迫均显著抑制三七的光合能力,与三七“喜冬暖夏凉,畏严寒酷热”的生长习性相符。低温胁迫下,三七主要通过提高其叶片POD和SOD活性,增加可溶性蛋白和叶绿素含量来增强其抗冷性,从而缓解低温对其造成的伤害;高温胁迫下,三七主要通过提高POD和SOD活性来增强其抗热性,从而缓解高温对其所造成的伤害。
