芸薹属作物花粉发育相关基因MSI的生物信息学分析
2017-05-30唐文武吴秀兰李桂花
唐文武 吴秀兰 李桂花

摘要:【目的】利用生物信息学分析芸薹属作物MS1基因的结构功能,为作物杂种优势利用提供理论参考。【方法】以拟南芥花粉发育关键基因AtMS1为参考序列,通过BLAST比对获得同源基因序列,运用生物信息学方法对其编码氨基酸序列进行预测分析。【结果】从甘蓝型油菜、白菜、甘蓝等芸薹属作物基因组中获得4条同源序列,与AtMS1基因的相似性在88.0%以上,均含有3个外显子,其CDS序列长度均为2004 bp,编码667个氨基酸。4个芸薹属作物MS1蛋白均含有1个植物同源结构域(Plant homeodomain,PHD),属于亲水性不稳定蛋白,定位于细胞核,磷酸化以丝氨酸(Ser)为主,以苏氨酸(Thr)和酪氨酸(Tyr)为辅;二级结构均由α-螺旋、β-转角、延伸链和无规则卷曲组成,其中α-螺旋所占比例最高,在40.00%以上,β-转角所占比例最低,仅为10.00%左右;其三级结构大致相同,均为球状的功能结构域。4个芸薹属作物MS1蛋白和AtMS1蛋白序列的相似性为95.69%。4个芸薹属作物MS1蛋白的PHD结构域序列高度保守,仅有3个位点氨基酸残基存在差异。19个不同植物的MS1同源蛋白聚为两大类,其中琴叶拟南芥、亚麻荠、萝卜的MS1蛋白与4个芸薹属作物MS1蛋白及拟南芥AtMS1蛋白聚为一类,均属于十字花科植物,即MS1蛋白的聚类结果与植物系统分类结果相吻合。【结论】芸薹属作物MS1基因属于PHD-finger基因家族,其序列高度保守,参与调控花粉发育成熟过程。
关键词:芸薹属;MS1基因;生物信息学;花粉发育;同源序列
中图分类号:S634.03 文献标志码:A 文章编号:2095-1191(2017)12-2122-07
0引言
【研究意义】雄性不育是植物有性繁殖过程中雄蕊发育异常引起无法授粉结实的现象,而花粉败育是雄性不育的表型特征,因此揭示花粉发育分子调控机理是了解雄性不育的关键。十字花科芸薹属作物存在明显的杂种优势,通过雄性不育系生产F1代种子是利用杂种优势的主要手段(唐文武等,2005)。目前,芸薹属的甘蓝(Brassica oleracea L.)、白菜(B.rapa L.)和甘蓝型油菜(B.napus L.)存在丰富的雄性不育变异材料,主要为核不育、细胞质不育及核质互作不育类型,在生产上已得到较好地利用,但其雄性不育的分子调控机理尚不明确。十字花科鼠耳芥属(拟南芥属)的拟南芥(Arabidopsis thaliana L.)作为模式植物,其花粉发育分子调控机理与雄性不育研究较深入,研究发现大多数从拟南芥克隆获得的育性调控基因在其他植物中均有同源基因,且表现出高度的进化保守性和相似的结构功能(Ma et al.,2015)。因此,以拟南芥相关研究为基础,预测分析芸薹属植物同源基因的结构功能,对了解油菜、甘蓝、白菜等作物的雄性不育分子机理及其杂种优势利用具有重要意义。【前人研究进展】花粉发育是高度复杂的生物学过程,由造孢细胞分化成花粉母细胞,经减数分裂形成四分体,然后在绒毡层酶的作用下形成单核的自由小孢子,再经过两次有丝分裂,最终发育成成熟的花粉粒,任何环节发生异常均可能导致雄性不育(McCormick,1993;Hong et al.,2012;Zhang et al.,2017a)。此外,花粉壁构成异常(Shi et al.,2011;Aloisi et al.,2016)及其最内层的绒毡层细胞发育不完全(Li et al.,2011;Ma et al.,2015)也是花粉败育的主要原因。至今,已从拟南芥克隆获得13个雄性不育相关基因,其中拟南芥MS1基因(AtMS1)是调控花粉形成的关键基因(Re-imegfird et al.,2017),主要在四分体至小孢子释放期表达,其编码含植物同源结构域(Plant homeodo-main,PHD)的核蛋白,与花粉壁构成及其绒毡层发育密切相关(no et al.,2002;Yang et al.,2007)。水稻PTCl基因为AtMS1同源基因,同样编码含PHD结构域的核蛋白,表现出相似的调控绒毡层细胞发育和花粉壁构成的功能,且PTC1基因能与拟南芥突变体atms1实现部分遗传互补,说明在单子叶植物和双子叶植物中均存在一个控制雄蕊程序化发育的PTC1/MS1开关(Li et al.,2011;Zhang et al.,2017b)。由于擬南芥突变体armsl在表型方面仅表现花粉败育,植株形态与花器官生长发育均正常(Wilson et al.,2001),因此AtMSl基因在生产上具有较高应用价值。【本研究切入点】芸薹属植物作为重要的油料与蔬菜作物种质来源,尤其随着白菜(Wang et al.,2011)、油菜(Chalhoub et al.,2014)和甘蓝(Liu et al.,2014)全基因组序列的公布,其雄性不育的分子生物学研究已成为热点,但鲜见芸薹属作物雄性不育相关基因的研究报道。【拟解决的关键问题】以AtMSl基因序列为参考,搜索并下载其同源序列,利用生物信息学方法对其编码蛋白质的理化性质、二级结构、三级结构、功能结构域及进化关系进行预测分析,为芸薹属作物雄性不育的分子机制和核不育系创制工作提供参考。
1材料与方法
1.1数据来源
从拟南芥TAIR网站(http:∥www.arabidopsis.org/)下载其花粉发育关键基因AtMS1的基因序列,其序列号为AT5G22260。以AtMS1基因序列为参考,在NCBI数据库(https:∥www.ncbi.nlm.nih.gov/)和芸薹属BRAD基因组数据库(http:∥brassicadb.org/brad/)中进行BLAST比对,设筛选标准为序列相似性≥88.0%。
1.2生物信息学分析
采用ProtParam进行蛋白质理化性质预测,利用SignalP 4.1进行蛋白质信号肽预测,以ProtScale进行蛋白亲/疏水性分析,利用WoLF PSORT进行亚细胞定位预测,通过SOPMA在线工具进行蛋白二级结构预测,以SWISS-MODEL进行三级结构预测,利用NetPhos 3.1进行蛋白质的磷酸化位点预测,采用SMART进行蛋白质保守结构域预测,并以DNAman和MEGA 5.2进行不同物种的同源蛋白多重比较和构建系统发育进化树。
2结果与分析
2.1 AtMS1基因同源序列获取
从芸薹属作物基因组中获得4个与AtMSl基因序列相似性大于88.0%的同源序列,其中2条序列来自甘蓝型油菜(B.napus),分别位于A基因组的3号和10号染色体,将其命名为BnMS1A03(序列号106443696)和BnMSlAl0(序列号106370785);1条序列来自白菜(B.rapa),位于A基因组的10号染色体,命名为BrMS1A10(序列号103845560);1条序列来自甘蓝(B.oleracea),位于C基因组的9号染色体,将其命名为BoMS1C09(序列号106316680)。这4个基因均含有3个外显子,其CDS序列长度均为2004 bp,编码667个氨基酸。
2.2 MS1蛋白的理化性质及亚细胞定位预测结果
蛋白质理化性质预测结果如表1所示,4个MS1基因所编码蛋白的分子量约76.5 kD,不稳定指数均超过41.00,属于不稳定蛋白;理论等电点(pI)以BoMS1C09最低(8.17),以BrMs1A10最高(8.47);脂肪氨基酸系数以BoMS1C09和BnMslA03最高(89.81),以BnMS1A10最低(86.01);疏水性系数均为负值(-0.319~-0.356),属于亲水性蛋白;均定位于细胞核。
2.3 MS 1蛋白的保守结构域及磷酸化位点分析结果
由图1可知,4个MSl蛋白均含1个PHD结构域,且PHD结构域均位于661-657 aa,其前端有一个低复杂区。同样,AtMSl蛋白仅含1个PHD结构域,位于616~662 aa。由此推测,MSl蛋白属于PHD-fmger蛋白家族成员,N_PHD结构域在芸薹属进化中高度保守。PHD结构域含Cys4-His-Cys3(C4-HC3)锌结合基序,其形态一般为球状,能结合2个锌离子而保持稳定的空间结构,在基因转录和染色质状态调控方面发挥重要作用(张帆等,2009)。
4个MS1蛋白磷酸化位点预测结果显示,其均含有较多的磷酸化位点,以丝氨酸(Ser)磷酸化位点最多,其中又以BrMS1A10最多(30个位点),BoMS1C09和BriMS1A03最少(26个位点);其次是苏氨酸(Thr)磷酸化位点,以BnMS1A10最多(17个位点),BoMS1C09最少(12个位点);以酪氨酸(Tyr)磷酸化位点最少,其中BoMS1C09最多(8个位点),而BrMS1A10和BnMS1A10最少(6个位点)(图2)。可见,MS1蛋白的磷酸化以Ser为主,以Thr和Tyr磷酸化为辅。
2.4 MS1蛋白的二、三级结构预测结果
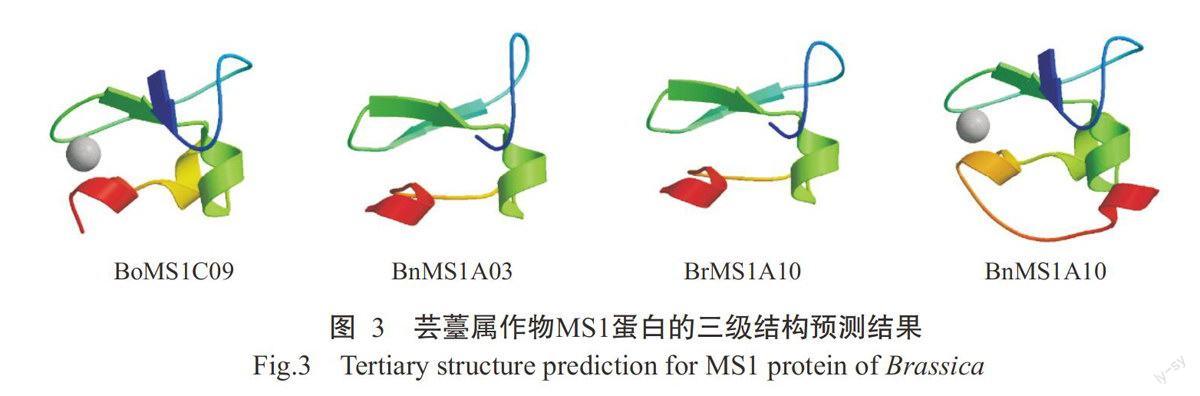
二级结构预测结果如表2所示,4个MS1蛋白的二级结构均由α-螺旋、β-转角、延伸链和无规则卷曲组成,其中α-螺旋为主要结构元件,所占比例均在40.00%以上,可能与此类蛋白含有PHD结构域和亮氨酸(Leu)拉链的结构有关,而β-转角所占比例最低,仅为10.00%左右。对4个MS1蛋白进行同源建模,预测其三级结构。结果显示,4个MS1蛋白均含有一个PHD结构域,位于616~652 aa,具有典型的C4HC3锌结合基序;三级结构大致相同,均为球状的功能结构域(图3),有利于特异性识别组蛋白H3K4的甲基化密码,从而发挥基因转录调控功能。
2.5 MS1蛋白的同源性分析结果
利用DNAMAN对拟南芥、白菜、甘蓝和油菜中的MS1蛋白进行多序列比对分析,结果如图4所示,5个MS1蛋白的氨基酸序列相似性为95.69%,表明其同源性极高,其中4个芸薹属作物MS1蛋白的氨基酸序列相似性更高,达98.39%。4个芸薹属作物MS1蛋白双序列比对结果显示,BoMS1C09与BnMS1A03的氨基酸序列相似性最高,达99.55%,BrMs1A10与BnMS1A10的氨基酸序列相似性也高达98.35%。拟南芥AtMS1与4个芸薹属作物MS1蛋白的双序列比对结果显示,AtMS1与BrMs1A10、BoMS1C09、BnMS1A03、BnMS1A10的氨基酸序列相似性分别为88.15%、88.44%、88.59%和88.59%,表明十字花科植物MS1基因进化中,拟南芥与芸薹属作物的亲缘关系较远,而3个芸薹属作物(白菜、甘蓝和甘蓝型油菜)的亲缘关系较近。
由图4还可知,5个MS1蛋白PHD结构域的序列高度保守,仅有3个位点氨基酸残基存在差异。方框内的7个半胱氨酸(Cys)和1个组氨酸(His)构成了C4HC3锌结合基序,PHD结构域的共有序列为C-X1-C-X12-C-X2-C-X4-H-X2-C-X14-C-X2-C,与张帆等(2009)的研究结果一致。PHD结构域Cys C2与C3之间的12个氨基酸残基形成Loopl,而C5与C6之间的14个氨基酸残基形成Loop2,每环各与1个锌离子结合形成稳定的Cross-brace拓扑结构,此类PHD结构域能与特异的核蛋白相互作用,进而调节基因转录表达。同时Cys之间及Cys与His之间的氨基酸较保守,与Bienz(2006)的研究结果一致。表明MS1蛋白在进化过程中其序列高度保守性,不易发生变异,以确保其花粉育性调控功能的稳定性。
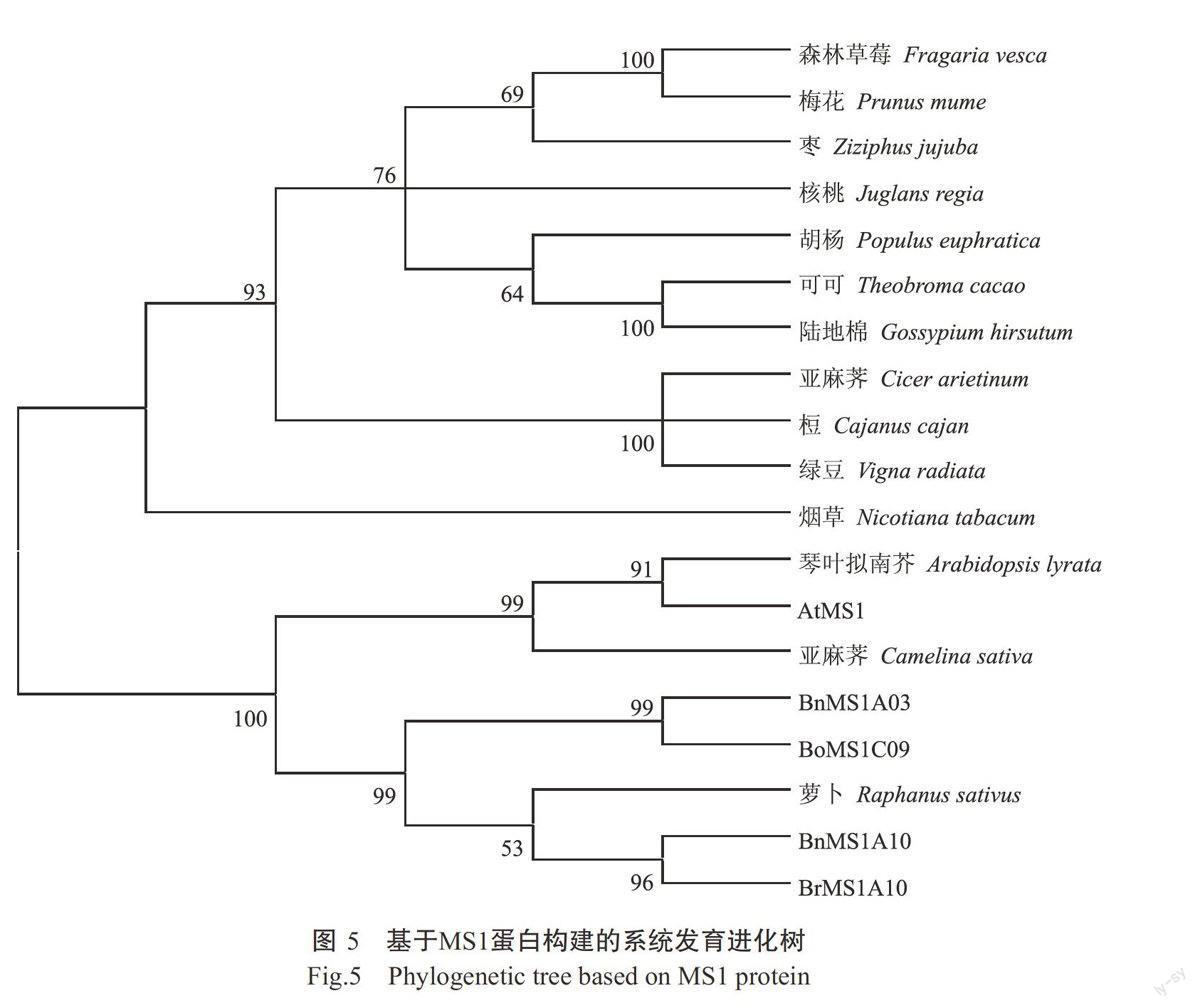
2.6 MS1同源基因的系统进化分析结果
通过NCBI等数据库检索,从豆科的木豆(Caja-nus cajan)、鹰嘴豆(Cicer arietinum)、绿豆(Hgna radiata)、蔷薇科的森林草莓(Fragaria vesca)、梅花(Prunus mume)、茄科的烟草(Nicotiana tabacum)、杨柳科的胡杨(Populus euphratica)、梧桐科的可可(Theobroma cacao)、鼠李科的枣(Ziziphus Jujuba)、锦葵科的陆地棉(Gos~pium hirsutum)、胡桃科的核桃(Juglans regia)及十字花科琴叶拟南芥(Arabidop-sis lyrata)、亚麻荠(Camelina sativa)、萝卜(Rapha-nus sativus)等植物中獲得MS1同源蛋白氨基酸序列,均含PHD结构域,被预测为雄性不育的同源蛋白序列。将其与AtMS1和4个芸薹属MS1蛋白(BnMS1A03、BnMSlA10、BrMS1A10和BoMS1C09)进行聚类分析并构建系统发育进化树,设自举值(Bootstrap)为1000。结果如图5所示,19个MS1同源蛋白聚为两大类,其中琴叶拟南芥、亚麻荠和萝卜的MS1蛋白与4个芸薹属MS1蛋白及AtMS1蛋白聚为一类,均属于十字花科植物;其他科的植物MS1蛋白聚为另一类,其中豆科的木豆、鹰嘴豆和绿豆的MS1蛋白聚为一小簇,蔷薇科森林草莓、梅花与鼠李科枣的MS1蛋白聚为一小簇,而锦葵科的陆地棉、梧桐科的可可与杨柳科的胡杨MS1蛋白聚为一小簇。该聚类结果与植物系统分类关系、进化历程基本吻合,因此该类蛋白可用于植物亲缘关系鉴定及遗传进化分析。
3讨论
PHD是真核生物中广泛存在的一种进化保守的锌指结构域,目前在400多种真核生物蛋白质中均有发现(Kaadige and Ayer,2006),由50~80个氨基酸残基组成,具有典型的C4HC3锌结合基序,其7个半胱氨酸残基和1个组氨酸残基结合2个锌离子而形成2个环,促使整个结构域保持稳定并发挥调节功能(Townsley et al.,2004)。PHD结构域是14种已知的锌指结构域中的一种,作为组蛋白密码的解读器之一,在基因转录调控、细胞周期调控等过程发挥重要作用。但这些研究主要集中在人类、小鼠等动物上,在植物研究较少,包括PHD-finger基因家族蛋白在植物开花调控、逆境胁迫应答等方面的研究。PHD-finger基因家族在植物开花调控方面发挥重要作用,如在拟南芥春化途径中,含PHD结构域的春化不敏感蛋白(VIN3)能通过对开花抑制基因FLC的染色质组蛋白进行甲基化、乙酰化等共价修饰,使得FLC染色质从松弛状态转为凝缩状态,通过关闭FLC转录活性而促进开花(Kim and Sung,2017);拟南芥At-MS1基因编码含PHD结构域的核蛋白与花粉壁构成及其绒毡层发育有关(Ito et al.,2007;Yang et al.,2007);水稻PTC1基因编码含有PHD结构域的核蛋白调控花粉壁构成及绒毡层细胞发育,形成控制雄蕊程序化发育的PTC1/MS1开关(Li et al.,2011;Zhang et al.,2017b)。PHD-finger基因家族蛋白在生物逆境应答方面也发挥重要作用,如水稻OsPHD1在逆境环境下表达量明显升高,且转基因水稻通过调节OsPHD1基因表达量来调控其抗逆性(刘雨等,2011);玉米PHD-finger基因家族在玉米发育过程中广泛表达,其中有15个基因可能与逆境胁迫应答密切相关(Wang et al.,2015)。也有研究表明,在拟南芥雄蕊减数分裂前期Ⅰ至中期Ⅰ中,染色质蛋白MMD1/DUET中的MMD1蛋白PHD结构域在染色体压缩及与甲基化的组蛋白尾结合中发挥重要功能作用(Wang et al.,2016)。本研究以AtMS1基因为参考,从NCBI数据库中筛选获得芸薹属植物(甘蓝型油菜、白菜和甘蓝)的MS1同源基因,其编码蛋白均含有PHD结构域,属于PHD-fmger基因家族成员(Wilson et al.,2001),定位于细胞核内,与AtMS1蛋白(Ye et al.,2010)和水稻PTC1蛋白(Li et al.,2011)的亚细胞定位结果一致。
PHD-finger基因家族蛋白主要通过甲基化、磷酸化、乙酰化、泛素化等方式修饰组蛋白,从而改变染色质的状态而参与基因的转录调控,如動物ME-KK1蛋白的PHD结构域具有E3泛素连接酶活性,能介导ERK1的泛素化和降解(马红辉等,2008);PHD结构域能特异识别并结合组蛋白H3K4进行甲基化修饰,从而发挥基因转录调控功能,如肿瘤抑制蛋白ING家族的PHD结构域能识别H3K4me2/3,并将复合物结合在D1启动子上,从而抑制基因转录(Shi et al.,2006)。磷酸化作为重要的蛋白修饰方式,由激酶和磷酸酶分别催化磷酸化和去磷酸化反应,在花药发育过程中参与不同信号转导(Yang et al,2015),但鲜见蛋白磷酸化与花粉发育相关性的研究报道。本研究中4个芸薹属作物MS1蛋白含有较多的磷酸化位点,且各蛋白的磷酸化以Ser为主,以Thr和Tyr磷酸化为辅,但由于蛋白磷酸化具有短暂性、高动态性及低丰度等特点,因此MS1蛋白的生物学功能与各位点的磷酸化修饰相关性尚需进一步研究。
4结论
芸薹属作物MSl基因属于PHD-finger基因家族,其序列高度保守,参与调控花粉发育成熟过程。
