珍稀植物华木莲的现代濒危机制:探析与展望
2017-05-30杨清培肖智勇胡晓东欧阳明陈昕林国江徐佳文杨光耀
杨清培 肖智勇 胡晓东 欧阳明 陈昕 林国江 徐佳文 杨光耀
摘要: 华木莲(Sinomanglietia glauca)在探讨木兰科系统演化与被子植物起源等方面具有重要科学价值,但其分布区域狭窄、种群规模较小,被列为我国濒危植物和极小种群植物。该文在系统综述华木莲生物生态学特性和更新维持机制的基础上,对华木莲的现代濒危原因进行了总结:(1)华木莲属古老残遗濒危植物,环境变迁、瓶颈效应、高频自交和遗传漂变,导致其遗传多样性下降、适应性差;(2)喜光喜肥的生物生态学特性和群落种间竞争驱赶作用,“林—窗”游击式更新是华木莲种群延续的重要方式,然而当前人类干扰导致有效林窗供给不足、种子传播效率下降、“游击”机会减少,更新难以完成;(3)毛竹扩张与林下箬竹过度增长严重影响华木莲开花结实,并妨碍其幼苗更新。今后应加强华木莲谱系地理与群落系统发育、群落环境对华木莲生長发育的影响及其应答、林窗更新机制与种子长距传播、群落结构调整与保护技术、遗传复壮与迁地保护等方面研究,为华木莲以及相似生物生态学特征的濒危植物保护提供理论指导。
关键词: 华木莲, 濒危机制, 竞争驱赶, “林—窗”游击更新, 有效林窗, 保护生态学
中图分类号: Q948
文献标识码: A
文章编号: 10003142(2017)05065308
Abstract: Sinomanglietia glauca, a species with scientific value to reveal magnoliaceae evolution and the origin of angiosperms, distributing discretely in Jiangxi and Hunan provinces, is catalogued as a rare and endangered plant and wild plant species with extremely small populations. The current endangered causes of S. glauca were systematically reviewed in light of its bioecological characteristics and regeneration mechanisms. The results were as follows: (1) As an ancient relic, history environmental change, the bottleneck effect, high frequency selfing and genetic drift, all could result in a decline in genetic diversity and poor adaptability for S. glauca. (2) “ForestGap” guerrilla regeneration model was an important way for this species surviving, due to its light and nutrientdemanding characteristics, and exclusion stress from interspecific competition in community. However, this model was interrupted by insufficient forest gaps and lower seed dispersal efficiency, less chance to guerrilla. (3) Bamboo expansion and overgrowth seriously reduced S. glauca flowering and fruiting, and interfered seedlings recruitment. Anyhow, the main causes of threats to this resource are selfing genetic recession, interspecific competition exclusion, insufficient effective forest gaps. The information of endangered mechanism and protection ecology is still very weak, the following are hoped to be strengthened in the future: the phylogeography and community system development of S. glauca at the molecularlevel, how environment factors affect the growth and development of S. glauca and its responses, “ForestGap” regeneration mechanism and seed longdistance dispersal, conservation technology by community adjustment, genetic rejuvenation and reintroduction. The review can provide theoretical guidance for S. glauca and other rare and endangered plants with similar bioecological characteristics.
Key words: Sinomanglietia glauca, endangered mechanism, competitive exclusion, “ForestGap” guerilla regeneration model, effective gap, conservation ecology
生物多样性是人类赖以生存的物质基础,一个基因决定一个国家的命运早已得到印证,然而人类活动导致的生物灭绝速率远超过地质历史的任何时期,生物多样性丧失已成为当前全球最严重的环境问题之一(钱迎倩等,1994; Cardinale et al, 2012; Pimm et al, 2014)。在我国近30 000种高等植物中,至少有3 000多种处于受威胁或濒临灭绝的境地,远高于世界平均水平(LópezPujol et al, 2006; Ren et al, 2014)。一个物种消失常联级性地导致其他10~30种生物的生存危机(黎磊等, 2014)。因此,揭示珍稀植物濒危机制和探索有效保护途径已成为当前保护生物学急需解决的重大课题。
华木莲(Sinomanglietia glauca)又名落叶木莲(Manglietia decidua),是20世纪90年代发现的一种木兰科植物。它在探讨木兰科系统演化与被子植物起源等方面具有极其重要的科学价值(俞志雄等, 1999; Li & Conran, 2003; 廖文芳等, 2004),但其分布区狭窄,仅孤岛状分布于江西省宜春市明月山和湖南省永顺县朗溪和回龙乡,现分布面积总计不足36 km2(廖文芳等, 2004; 侯伯鑫等, 2006),且母树稀少,结实率低,天然更新困难,处于珍稀濒危的境地(俞志雄和李志强, 1999; 侯伯鑫等, 2006)。1999年华木莲被列为我国一级保护植物,2001年列入IUCN濒危植物红皮书(廖文芳等, 2004),现又被列入全国极小种群野生植物拯救保护工程规划(2011-2015年)(http://rep.iplant.cn/protlist/6)。近年来对华木莲系统发育(俞志雄和李志强, 1999)、遗传变异(林新春等, 2003;廖文芳等, 2004;熊敏等, 2014)、物候(余格非等, 2007; 詹鹏等, 2013)、木材解剖(俞志雄和李志强, 1999)、引种繁殖(龚德海等, 2011)、园林应用(施建敏等, 2013)、化工利用(施建敏等, 2006;庞会忠等, 2007)等进行了研究,但对华木莲的濒危机制研究缺乏系统分析。
本文总结了近年来华木莲生物学、生态学、遗传学等方面的研究成果,在分析其生物生态学特性及更新机制的基础上,重点探索华木莲濒危的现代原因,并指出今后亟待解决的相关科学问题和研究方向,以期为充分认识、合理利用与有效保护华木莲这一珍稀濒危种质资源提供参考。
1喜光喜肥的生物生态学特征
华木莲是一种阳性喜光植物,除幼苗外,中年、成年大树均不耐荫蔽,喜酸性肥沃而不耐瘠薄,喜凉爽湿润气候而不耐干燥(俞志雄等, 1999; 施建敏等, 2013)。另外,华木莲是一种速生树种,成熟时间早,生理年龄相对较短。如15年生的华木莲树高、胸径的平均生长量分别为0.82 m、1.27 cm,树高、胸径生长成熟年龄分别为20~40、40~60 a(俞志雄等, 1999)。与近缘种乳源木莲(Manglietia yuyuanensis)、巴东木莲(M. patungensis)相比,华木莲净光合速率、蒸腾速率、表观量子产额、羧化效率、光补偿点较高(郭起荣等, 2004),这从生理学上反映其阳性喜光特征。从群落层次看,华木莲群落次生性明显,其种群主要分布在次生落叶阔叶林、落叶—常绿阔叶混交林、毛竹林,偶残存于天然常绿阔叶林片段(俞志雄和李志强, 1999; 侯伯鑫等, 2006)。
2“林—窗”游击式更新方式
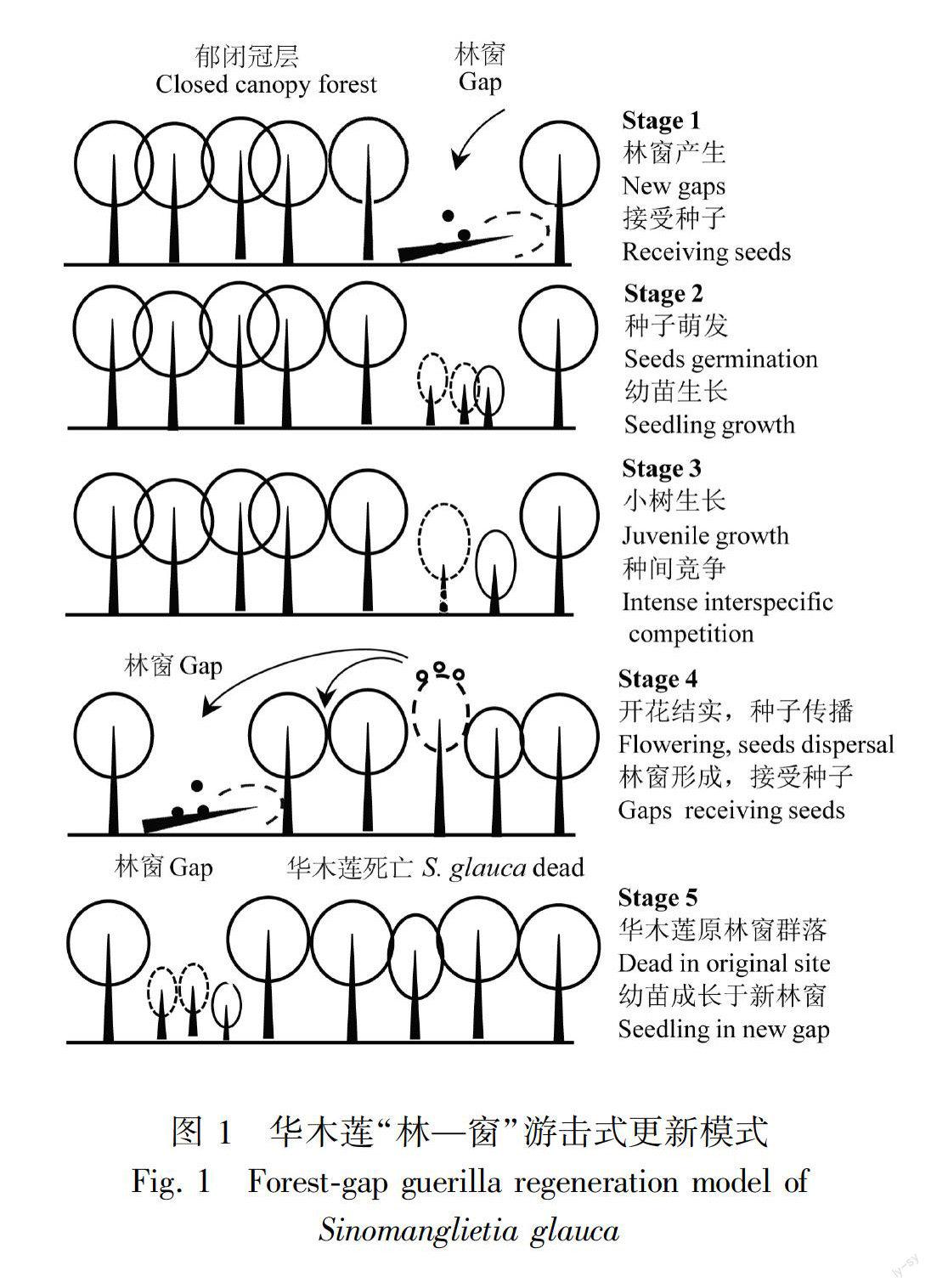
任何种群的延续离不开一代代的自我更新。第四纪冰期我国东部出现了干冷与湿暖气候的交替变化,极不利于植物的生长。华木莲在历史气候变迁中经历了严重的瓶颈效应后(廖文芳等, 2004),部分居群残存了下来成为孑遗植物。从区域景观视角看,这些残遗居群主要依靠林窗或疏林(源于自然干扰或大树自然死亡)进行游击式的自然更新,才一直延续至今(俞志雄和李志强, 1999)。这种更新方式可划分为(1)林窗形成;(2)种子萌发;(3)幼苗生长;(4)开花结实;(5)竞争退出等5个阶段,本文称之为“林—窗”游击更新模式(ForestGap guerilla regeneration model),也因此可知华木莲是一种林窗依赖型植物(图1)。
(1)林窗形成:由于受大风、冰雪、滑坡等外界因子的干扰,或大树的生理衰老、病虫害等的原因,华木莲分布区域内成熟森林中有大树倒伏或树冠破损,郁闭林冠开放,形成林窗,并接受周边种子的迁入,其中有些林窗接受到华木莲种子。
(2)种子萌发:林窗形成后,光照或温湿度条件得到大大改善,有利于原土壤里的种子或新接受的种子萌发,产生幼苗。开始主要是华木莲、华桑(Morus cathayana)、山樱花(Cerasus serrulata)、青榨槭(Acer davidii)等阳性树种,而耐荫种的幼苗生长受到抑制,成活率不高,其组成比例相对较小。林窗面积越大,阳性树种比例就越高,华木莲出现的机会也就越大(调查资料)。
(3)小树成长:经过种内(间)竞争,部分胜出的华木莲个体逐渐由小树成长大树。此时耐荫树种也逐渐萌发、幼苗生长,耐荫树种的组成比例逐渐增加,林窗群落组成与结构变得复杂(俞志雄和李志强, 1999; 侯伯鑫等, 2006)。
(4)开花结实:少数华木莲长成大树,并进入生殖阶段,那些远高出常绿阔叶树的个体开花结实(俞志雄和李志强, 1999; 侯伯鑫等, 2006)。有些果实借助自身重力滚动传至下坡,有些种子借动物传播远方,其中有些种子被传到新形成的林窗,进入新一轮的生活史。
(5)退出群落:随着林窗群落的发育,耐荫常绿树种增多,资源竞争加剧,华木莲等阳性树种生长减缓、衰老加速,最后被细叶青冈(Cyclobalanopsis myrsinaefolia)、少花桂(Cinnamomum pauciflorum)等耐蔭种替代而退出林窗群落(侯伯鑫等, 2006)。至此,在林窗处华木莲完成从种子、幼苗、成株到死亡的生活史。
可见,林窗形成是华木莲更新的必要条件,这与许多其他林窗依赖型孑遗植物的更新维持机制非常相似(Tang et al, 2002, 2012)。这种更新方式具有明显的随机性和不稳定性,受到的威胁较大。
3现代濒危的原因
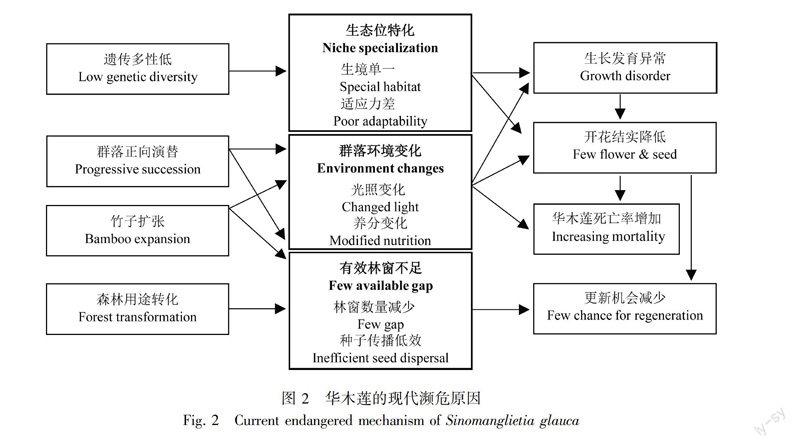
当前日益频繁的人类活动,使华木莲“林—窗”游击式更新模式受到极大挑战,其种群维持受到极大威胁,其濒危程度日益加剧(图2)。具体原因与过程表现:(1)遗传多样性下降;(2)群落正向演替;(3)有效林窗不足;(4)竹子扩张;(5)不合理利用与破坏。
3.1 遗传多样性低,适应能力差
不论是江西明月山居群,还是湖南永顺朗溪居群和云盘居群,华木莲遗传多样性都较低(林新春等, 2003; 熊敏等, 2014)。廖文芳等(2004)采用ISSR技术,分析江西明月山华木莲居群的遗传多样性,发现其物种水平的遗传多样性HT为0.065,居群水平的遗传多样性HS为0.052,多态位点百分率P为17.28 %,远低于木兰科的其它植物(廖文芳等, 2004)。林新春(2003)利用RAPD分子标记,发现江西明月山华木莲居群遗传多样性水平低于观光木(Michelia odora)、北美鹅掌楸(Liriodendron tulipifera)
及鹅掌楸(Liriodendron chinense),而与水杉(Metasequoia glyptostroboides)相当(林新春等, 2003)。
较低的遗传多样性,导致较低的竞争力与适应性,造成生境特化,环境敏感。经过瓶颈效应幸存下来的华木莲种群偏居一隅,分布区范围没有得到有效扩张,且只能生活在光照充足、土壤肥沃的优越生境。华木莲所在群落正向发展,环境条件不断改变、种间资源竞争加剧时,华木莲生长发育受到严重影响,成活率下降、死亡率上升、开花结实率低(俞志雄和李志强, 1999; 裘利洪等, 2004)。在良好环境没有竞争力,在胁迫环境又没有繁殖力,说明该物种适应力差。同时种群隔离、种群规模小,加之较强的自然或人为破坏, 导致华木莲近期居群衰退、自交频率上升、遗传漂变加速,遗传多样性进一步丧失(熊敏等, 2014)。遗传多样性低严重制约着物种适应性进化的水平,降低了华木莲的环境适应能力(廖文芳等, 2004),形成恶性循环,成为华木莲现代濒危的内在原因。
3.2 群落正向演替,种间竞争驱赶
尽管在自由木或纯林中,华木莲不但长势良好,而且开花结实率高(龚德海等, 2011),但在野外华木莲很少呈单株孤立或同种纯林,而总是以群落形式存在。随着群落发育与演替,细叶青冈、少花桂、细叶香桂(Cinnamomum pauciflorum)等常绿耐荫树种增多,群落郁闭度逐渐增加,华木莲幼树与小树受到大树遮荫,光照降低,光资源不足成为华木莲种群更新的主要限制因子(郭起荣等, 2004),当林分郁闭度达到0.8时,华木莲完全不能自我更新,导致壮树个体急剧减少,种群呈衰退趋势(俞志雄和李志强, 1999; 侯伯鑫等, 2006)。华木莲受竞争驱赶被迫退出群落,这是华木莲濒危的生物生态学原因。
3.3 有效林窗不足,更新难以完成
一定数量的林窗产生与种子传播是林窗更新的必要因素。华木莲受竞争驱赶被迫退出原群落后,其种子必须得到林窗接受,才能延续其更新,然而其分布区内许多天然林被改造成杉木林、毛竹林或其他经济林,即使形成部分林窗,也因经济利益的原因,华木莲幼苗、幼树被人为去除。或者在某些地段因反复干扰,土壤肥力下降,即使产生了林窗,华木莲也会因其喜湿喜肥的特性而竞争不过马尾松(Pinus massoniana)、枫香(Liquidambar formosana)、化香(Platycarya strobilacea)等耐贫瘠树种(裘利洪等, 2004),因此这些林窗成为无效林窗。另外,华木莲为聚合蓇葖果,种子圆形至椭圆形,种子长0.43~0.71 cm,宽0.51~0.84 cm(夏国华等, 2006),千粒重达83 g,种子擴散主要靠动物传播。因此,即使有林窗出现,也因传播种子的动物减少,扩散效率降低,而成为另一种无效林窗。可见,有效林窗不足、更新机会减少,成为华木莲现代濒危的重要原因。
3.4 竹类植物扩张,资源竞争加剧
竹子扩张加剧了华木莲濒危。华木莲适生于落
叶阔叶林、常绿—落叶混交林或常绿阔叶林受干扰而形成的林窗,然而这些地方也是毛竹(Phyllostachys pubescens)、箬竹(Indocalamus sp.)、方竹(Chimonobambusa quadrangularis)的适生生境。当这些群落受到干扰时,邻近竹类植物就会向这些地方扩张并大量繁殖,增加对华木莲的竞争压力。毛竹向华木莲群落扩张后,一方面改变了华木莲的受光状况,可能也会改变土壤养分化学计量学特征(宋庆妮等, 2013),从而影响华木莲的开花结实(俞志雄等, 1999)。箬竹、方竹等中小竹子在林下大量繁殖,不但改变了地表的光环境,而且其盘根错节的地下鞭根,加剧土壤养分竞争,也不利于华木莲种子发芽定居。因此,控制竹子扩张,有利于华木莲的就地保护(裘利洪等, 2004)。
3.5 不合理利用与破坏,种群数量减少
华木莲除了具有重要科学研究价值外,其树形端庄、树姿清新飘逸,花香沁人心脾,花色淡黄、典雅宛若睡莲,也具有极高的园林观赏价值(施建敏等, 2013),曾被定为江西省第十届运动会的吉祥物和宜春市的市花。曾有许多群众慕名而上山盗挖华木莲小苗、采摘果实,妨碍华木莲正常更新。另外,一方面主管部门对华木莲只有大树记录而幼苗无记录,当地对毛竹林习惯于劈山抚育、纯林经营,因此竹林经营时常将华木莲幼苗砍除,直接导致了华木莲种群数量减少(廖文芳等, 2004)。因此,不合理利用和破坏是华木莲濒危的又一直接原因。
4研究展望
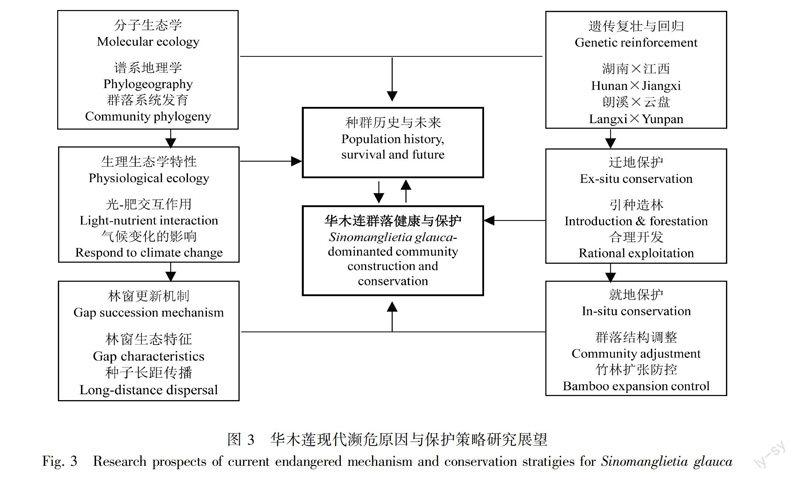
华木莲是一个刚被发现20 a的物种,人类对其认识还有限,虽然从生物生态习性、遗传多样性、种间竞争驱赶、有效林窗不足、不合理利用与破坏等方面,初步分析华木莲濒危的原因,但这些研究还很不完善,仍停留在定性描述与表观层面。要深入了解华木莲的过去、现在和未来,并采取积极有效的保护措施,无疑还有很长的路要走。我们认为今后应重点加强以下研究:(1)华木莲谱系地理学与群落系统发育学;(2)多因子(尤其是光照-养分)交互作用对华木莲生长生殖策略的影响;(3)种子长距传播与林窗更新格局;(4)群落调整的就地保护技术;(5)居群间杂交复壮与回归技术(图3)。
4.1 华木莲谱系地理与群落系统发育研究
了解历史、查清资源是认识和保护濒危植物的基础。从20世纪90年代华木莲首次发现于明月山后,又在永顺发现(侯伯金等, 2006),后来又在江西分宜和安福发现有少量个体(未发表),说明华木莲资源数量和分布区范围还有待进一步查实。近年分子谱系地理学为濒危、珍贵植物的研究提供了全新的理论指导(Ribeiro et al, 2011; Kutnjak et al, 2014),促进了对多个濒危物种的有效保护(Chen et al, 2015)。在化石资料缺乏的情况下,可充分利用现代分子技术,积极开展华木莲谱系地理学研究,阐明华木莲系统发育与分类地位,揭示种群起源、演化、分布、迁移的时空规律与濒危历史原因。同时,构建群落系统发育的分子技术日趋成熟(CavenderBares et al, 2009; Kress et al, 2009)。因此,利用分子技术构建华木莲群落的系统发育关系,从历史的角度分析群落组成、种间关系状况,种间关系如何影响华木莲的遗传进化与生态适应性,为指导现代华木莲群落构建提供理论。今后重点研究:①华木莲资源与分布区现状调查; ②种群系统发育与分类地位确定; ③华木莲发生、迁移与分布格局的形成;④群落系统发育与种间关系演变。
4.2 群落环境对华木莲生长发育的影响及其应答
环境变化是影响华木莲生长竞争与开花结果的主要因素。群落发育过程中环境因子(尤其是光照、养分)的改变,导致华木莲生长发育异常、开花结果较少,而引种栽培的华木莲生长发育正常。因此,深入了解华木莲的环境敏感性及其有限可塑性,是华木莲保护的重要生理基础,但至今群落环境对华木莲的光合作用、养分吸收、能量分配、生长发育、开花结果等方面机制研究尚少,未能有效解释华木莲荫蔽胁迫与竞争驱赶效应。
同时,全球变化已成为了当前人类面临一个不争事实,而气候变化会给全球生物多样性带来难以预料的影响(Bell & Gonzalez, 2011; Bellard et al, 2012)。模拟气候变化对濒危生物生态学特性的影响,及其对全球变化、气候变暖、CO2浓度增加、降水格局变化的响应,预测濒危植物对未来环境条件变化的适应,是今后的重要研究内容(Brooker, 2006; 黎磊和陈家宽, 2014)。华木莲是历史气候变化的产物,加强气候变化对华木莲资源与分布影响的认识, 评价华木莲适应气候变化的能力, 是华木莲保护机制研究的需要,今后重点研究方向:①华木莲的环境敏感性生理机制及有限可塑性;②光照、养分对华木莲生长发育、开花结实的影响与应答;③华木莲与微生物间相互作用过程;④全球气候变化对华木莲迁移与分布的模拟与预测。
4.3 林窗更新机制与种子长距扩散过程
林窗更新是许多珍稀植物种群延续的重要方式,种子扩散是种群扩散的重要方式(谢宗强, 1999; 张志祥等, 2008)。然而林窗的起源、分布格局、形状大小、地被物、土壤理化性质等对华木莲更新的影响研究还知之甚少,加强华木莲本身与其他植物的生物生态学特性及林窗的环境条件,包括光合生理,蒸腾作用,物候规律,C、N、P等养分分配与利用规律等因素。开展华木莲林窗更新过程的长期监测,包括从种子萌发、幼苗生长、大树开花结实。同时,充分利用红外相机技术,深入开展华木莲种子传播动物及活动规律,分析种子散布、种子雨、种子库及幼苗更新格局。试在母树周边开启一些小面积林窗,營造对华木莲幼苗发育有利的生境,促进华木莲林窗天然更新和空间扩展,扩大现有居群。重点研究方向:①群落演替与竞争驱赶作用的长期定位研究;②林窗类型、大小和空间格局对华木莲更新的影响;③华木莲种子长距离传播监测与分析;④林窗更新过程长期定位研究。
4.4 群落结构调整与就地保护技术
竞争驱赶是华木莲濒危的一个主要原因。因此,在有效林窗形成不足的情况,避免竞争、增强互利是就地保护华木莲的具体措施。近年在江西和湖南分别建立了江西玉京山落叶华木莲自然保护区和湖南省小溪国家级自然保护区,这是保护华木莲的重大进步,但建立保护区,不是简单地封山育林。群落的物种组成、种群的空间结构都会直接影响到种间关系和群落发展,有人建议将群落内常绿阔叶树砍掉、降低林分郁闭度、营造疏林环境、蓇葖果成熟脱落后,清理林下灌木,尤其是竹类植物。那么,什么样的物种组成与群落结构有利于华木莲长期保护,同时又保证群落生态功能最大?在林分尺度上,调整物种组成、竞争关系和种群空间分布格局,构建华木莲群落的最优结构,以提高种群保护效果。今后重点研究:①华木莲群落动植物多样性研究;②群落种群空间配置与结构优化技术;③群落动植物种间关系;④生态系统整体功能评价与保护。
4.5 遗传复壮与迁地保护技术
一般认为,遗传多样性低是植物濒危的内在根本原因,遗传复壮是珍稀濒危植物保护生物学的重要策略。有人发现通过不同居群交互引种,可提高遗传变异水平、促进基因流(杨琴军等, 2006),但有人发现居群间杂交可能会带来杂交没化、远交衰退的风险,而建议采用加大居群内杂交(杨慧等, 2011)。华木莲孑遗居群间存在一定的地方差异性与遗传分化(Zhang et al, 2009; 熊敏等, 2014)。因此,居群间杂交是否可增强华木莲遗传适应性和生态适应性,还值得进一步研究。
迁地保护与回归重建是扩大种群的必要措施(林新春等, 2003; Ren et al, 2014),但种源选择、生境条件、种群规模与遗传管理对濒危植物保护的质量影响较大(Mace & Purvis, 2008; Bischoff et al, 2010)。因此在华木莲迁地保护或回归自然重建中,要尽可能地避免近交或远交衰退,保持遗传纯正性和足够的遗传基础,最大限度地保持华木莲环境适应能力、提高保护质量或效率,成功实现物种的复壮和种群自然回归。建议开展以下研究:①优良种源收集与遗传多样性分析;②生境选择与遗传变异性的关系;③居群间杂交与遗传适应性评价;④遗传多样性动态检测与有效群体构建。
5结论
华木莲为第四纪残遗濒危植物。环境变迁、瓶颈效应,导致其遗传多样性水平低、环境适应性差、竞争力弱,这是华木莲濒危的根本原因。同时,喜光喜肥的生物生态学特点和群落种间竞争的驱赶作用,决定了“林—窗”游击式更新模式是华木莲维持种群延续的重要方式,然而在当前人类干扰、森林转化、竹子入侵情况下,有效林窗形成不足、种子传播低效、更新机会减少,是华木莲濒危的主要原因。然而,华木莲濒危机制及保护生态学研究非常薄弱,建议今后加强该物种的起源、扩散与分布、多种环境因子对该物种生长生殖的影响、种子扩散与林窗更新机制、群落调整保护技术、遗传复壮与自然回归等方面研究,以期为华木莲以及类似生物生态学特征的濒危植物保护提供理论指导。
参考文献:
BELL G, GONZALEZ A, 2011. Adaptation and evolutionary rescue in metapopulations experiencing environmental deterioration [J]. Science, 332(6035): 1327-1330.
BELLARD C, BERTELSMEIER C, LEADLEY P, et al, 2012. Impacts of climate change on the future of biodiversity [J]. Ecol lett, 15(4): 365-377.
BISCHOFF A, STEINGER T, MLLERSCHRER H, 2010. The importance of plant provenance and genotypic diversity of seed material used for ecological restoration [J]. Restor Ecol, 18(3): 338-348.
BROOKER RW, 2006. Plantplant interactions and environmental change [J]. New Phytol, 171(2): 271-284.
CARDINALE BJ, DUFFY JE, GONZALEZ A, et al, 2012. Biodiversity loss and its impact on humanity [J]. Nature, 486(7401): 59-67.
CAVENDERBARES J, KOZAK KH, FINE PVA, et al, 2009. The merging of community ecology and phylogenetic biology [J]. Ecol Lett , 12(7): 693-715.
CHEN JM, ZHAO SY, LIAO YY, et al, 2015. Chloroplast DNA phylogeographic analysis reveals significant spatial genetic structure of the relictual tree Davidia involucrata (Davidiaceae) [J]. Conserv Genet, 16(3): 583-593.
GONG DH, CHEN Y, XIA MJ, et al, 2011. Preliminary report on adaptability Sinomanglietia glauca of northern Jiangxi [J]. Jiangxi For Sci & Technol, (6):24-25. [龔德海, 陈英, 夏敏娟, 等, 2011. 华木莲在赣北的适应性初报 [J]. 江西林业科技, (6):24-25.]
GUO QR, YU ZX, SHI JM, 2003. A physioecological study on photosynthesis of Sinomanglietiaca glauca and two species in Manglietia Bl [J]. Acta Agric Univ Jiangxi, 25(5):645-651. [郭起荣, 俞志雄, 施建敏, 2003. 华木莲与木莲属两树种光合生理生态研究 [J]. 江西农业大学学报, 25(5): 645-651.]
HONNAY O, JACQUEMYN H, 2007. Susceptibility of common and rare plant species to the genetic consequences of habitat fragmentation [J]. Conserv Biol, 21(3): 823-831.
HOU BX, YI H, LIN F, et al, 2006. Investigation on resource of Manglietia decidua of Yongshun County in Hunan [J]. Hunan For Sci & Technol, 33(1):7-10. [侯伯鑫,易宏,林峰, 等, 2006. 湖南永顺县落叶木莲资源考察初报 [J]. 湖南林业科技, 33(1):7-10.]
KRESS WJ, ERICKSON DL, JONES FA, et al, 2009. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama [J]. P Natl Acad Sci USA, 106(44): 18621-18626.
KUTNJAK D, KUTTNER M, NIKETIC' M, et al, 2014. Escaping to the summits: Phylogeography and predicted range dynamics of Cerastium dinaricum, an endangered high mountain plant endemic to the western Balkan Peninsula [J]. Mol Phylogenet Evol, 78: 365-374.
LI J, CONRAN JG, 2003. Phylogenetic relationships in Magnoliaceae subfam. Magnolioideae: a morphological cladistic analysis [J]. Plant Syst Evol, 242(1-4): 33-47.
LI L, CHEN JK, 2014. Influence of climate change on wild plants and the conservation strategies [J]. Biodivers Sci, 22(5):549-563. [黎磊, 陈家宽, 2014. 气候变化对野生植物的影响及保护对策 [J]. 生物多样性, 22(5):549-563.]
LIAO WF, XIA NH, DENG YF, et al, 2004. Study on genetic diversity of Manglietia decidua (Magnoliaceae) [J]. Acta Bot Yunnan, 26(1):58-64. [廖文芳,夏念和,邓云飞, 等, 2004. 华木莲的遗传多样性研究 [J]. 云南植物研究, 26(1):58-64.]
LIN XC, 2004. Pollen viability of four species of Magnoliaceae [J]. Jiangxi For Sci & Technol, (4): 4-5. [林新春, 2004. 4种木兰科植物花粉生活力的测定 [J]. 江西林业科技, (4):4-5.]
LIN XC, YU ZX, QIU LH, et al, 2003. Studies on genetic diversity of endangered Sinomanglietia glauca (Magnoliaceae) [J]. Acta Agric Univ Jiangxi, 25(6): 805-810. [林新春, 俞志雄, 裘利洪, 等. 2003. 瀕危植物华木莲的遗传多样性研究 [J]. 江西农业大学学报, 25(6): 805-810.]
LPEZPUJOL J, ZHANG FM, GE S, 2006. Plant biodiversity in China: richly varied, endangered, and in need of conservation [J]. Biodivers Conserv, 15(12): 3983-4026.
MACE GM, PURVIS A, 2008. Evolutionary biology and practical conservation: bridging a widening gap [J]. Mol Ecol, 17(1): 9-19.
PANG HZ, YU ZX, SHI JM, et al, 2007. The antioxillation effect of polysaccharide extracted from Sinomanglietia glaula leaf [J]. Acta Agric Univ Jiangxi, 28(6): 813-818. [庞会忠, 俞志雄, 施建敏, 等, 2007. 华木莲叶黄酮类化合物抗氧化作用 [J]. 江西农业大学学报, 28(6): 813-818.]
PIMM SL, JENKINS CN, ABELL R, et al, 2014. The biodiversity of species and their rates of extinction, distribution, and protection [J]. Science, 344(6187):1246752.
QIAN YQ, MA KP, 1994. The principle and methods research on biodiversity [J]. Beijing: China Science and Technology Press. [钱迎倩, 马克平, 1994. 生物多样性研究的原理与方法 [J]. 北京: 中国科学技术出版社.]
QIU LH, YU ZX, SHI JM, et al, 2004. Studies on interspecific association of communities with Sinomanglietia glauca [J]. Acta Agric Univ Jiangxi, 26(1): 25-30. [裘利洪, 俞志雄, 施建敏, 等, 2004. 华木莲群落种间联结性研究 [J]. 江西农业大学学报, 26(1): 25-30.]
REN H, JIAN SG, LIU HX, et al, 2014. Advances in the reintroduction of rare and endangered wild plant species [J]. Sci China Life Sci , 57(6): 603-609.
RIBEIRO RA, LEMOSFILHO JP, RAMOS ACS, et al, 2011. Phylogeography of the endangered rosewood Dalbergia nigra (Fabaceae): insights into the evolutionary history and conservation of the Brazilian Atlantic Forest [J]. Heredity, 106(1): 46-57.
SHI JM, QIU LH, HUNAG ZY, et al, 2013. The selection of landscape greening sites for sinomanglietia glauca: a study based on light intensity [J]. Guangdong Landsc Archi, 2013, (2): 62-64. [施建敏, 裘利洪, 黄芝云, 等, 2013. 基于光照强度探讨珍稀树种华木莲绿化生境的选择 [J]. 广东园林, (2):62-64.]
SHI JM, YU ZX, PANG HZ, et al, 2006. The antioxidation effect of sinomanglietia glaula leaf flavonoids [J]. Acta Agric Univ Jiangxi, 28(5): 693-697. [施建敏, 俞志雄, 庞会忠, 等, 2006. 华木莲叶黄酮类化合物抗氧化作用 [J]. 江西农业大学学报, 28(5):693-697.]
SONG QN, YANG QP, LIU J, et al, 2013. Effects of Phyllostachys edulis expansion on soil nitrogen mineralization and its availability in evergreen broadleaf forest [J]. Chin J Appl Ecol, 24(2): 338-344. [宋庆妮, 杨清培, 刘骏, 等, 2013. 毛竹扩张对常绿阔叶林土壤氮素矿化及有效性的影响 [J]. 应用生态学报, 24(2):338-344] .
TANG CQ, OHSAWA M, 2002. Tertiary relic deciduous forests on a humid subtropical mountain, Mt. Emei, Sichuan, China [J]. Folia Geobot, 37(1): 93-106.
TANG CQ, YANG YC, OHSAWA M, et al, 2012. Evidence for the persistence of wild Ginkgo biloba (Ginkgoaceae) populations in the Dalou Mountains, southwestern China [J]. Am J Bot, 99(8): 1408-1414.
XIA GH, HUANG JQ, TANG GG, et al, 2006. Identification of species from Magnoliaceae via seeds morphology [J]. J Nanjing For Univ, 29(5):97-100. [夏国华, 黄坚钦, 汤庚国, 等, 2006. 木兰科常用绿化树种种子形态鉴别 [J]. 南京林业大学学报:自然科学版, 29(5):97-100.]
XIAO DX, ZHANG JP, 1998. Karyotype analysis of Sinomanglietia glauca Z. X. Yu et Q. Y. Zheng [J]. Acta Agric Univ Jiangxi, 20(1):56-59. [肖德兴, 张津平, 1998. 华木莲染色体核型的研究 [J]. 江西农业大学学报, 20(1): 56-59.]
XIE ZQ, 1999. Gapregeneration of Cathaya argyrophylla forests [J]. Acta Ecol Sin, 19(6): 775-779. [谢宗强, 1999. 银杉(Cathaya argyrophylla)林林窗更新的研究 [J]. 生态学报, 1999, 19(6):775-779.]
XIONG M, TIAN S, ZHANG ZR, et al, 2014. Population genetic structure and conservation units of Sinomanglietia glauca (Magnoliaceae) [J]. Biodivers Sci, 22(4): 476-484 . [熊敏, 田双, 张志荣, 等, 2014. 华木莲居群遗传结构与保护单元 [J]. 生物多样性, 22(4): 476-484.]
YANG H, CHEN YY, XU YX, et al, 2011. Gene flow dynamics of exsitu conservation populations in two endangered Isoetes species: Genetic implications for reintroduction, conservation and management [J]. Plant Sci J, 29(3):319-330. [楊慧, 陈媛媛, 徐永星, 等, 2011. 两种濒危水韭植物迁地保护居群的基因流动态及回归重建保育遗传管理策略 [J]. 植物科学学报, 29(3):319-330.]
YANG QJ, XU H, YAN ZG, et al, 2006. Natural resources and conservation of Taiwania cryptomerioides in Hubei Province [J]. Guihaia, 26(5): 551-556 . [杨琴军, 徐辉, 严志国, 等, 2006. 湖北省原生台湾杉资源及其保护 [J]. 广西植物, 26(5): 551-556.]
YU GF, YI H, YANG J, et al, 2007. Study on the phenology of blooming and fruiting of Manglietia decidua Q. Y. Zheng [J]. Hunan For Sci & Technol, 33(6): 35-37. [余格非,易宏, 等, 2007. 永顺落叶木莲开花与结实物候学研究 [J]. 湖南林业科技, 33(6): 35-37.]
YU ZX, LI ZQ, 1999. Primary analysis on the growth process of Sinomanglietia glauca Z. X. Yu et Q. Y. Zheng [J]. Acta Agric Univ Jiangxi, 21(1): 95-98. [俞志雄 李志強, 1999. 华木莲生长过程的初步分析 [J]. 江西农业大学学报, 21(1):95-98.]
YU ZX, LIAO J, LIN XC, et al, 1999. Ecological studies on communities of Sinomanglietia glauca Z. X. Yu et Q. Y. Zheng [J]. Acta Agric Univ Jiangxi, 21(2): 213-217. [俞志雄,廖军,林新春,等, 1999. 华木莲植物群落的生态学研究 [J]. 江西农业大学学报, 21(2): 213-217.]
YU ZX, ZHENG QY, 1999. Wood structure of Sinomanglietia Z. X. Yu of Magnoliaceae [J]. Acta Agric Univ Jiangxi, 21(1): 91-94. [俞志雄, 郑庆衍, 1999. 木兰科华木莲属木材结构 [J]. 江西农业大学学报, 21(1):91-94.]
YU ZX, 1994. Sinomanglietia—a new genus of magnoliaceae from China [J]. Acta Agric Univ, 16(2): 202-204. [俞志雄, 1994. 华木莲属——木兰科一新属 [J]. 江西农业大学学报, 16(2):202-204.]
ZHAN P, XU WJ, CHEN JN, et al, 2013. Analysis of growth and genetic diversity of Manglietia decidua in Xiaoxi National Nature Reserve [J]. Hunan For Sci & Technol, 40(1): 57-60 . [詹鹏, 徐万吉, 陈介南, 等, 2013. 小溪国家级自然保护区落叶木莲生长与遗传多样性分析 [J]. 湖南林业科技, 40(1): 57-60.]
ZHANG ZR, LUO LC, WU D, et al, 2009. Two genetically distinct units of Sinomanglietia glauca (Magnoliaceae) detected by chloroplast PCRSSCP [J]. J Syst Evol, 47(2): 110-114.
ZHANG ZX, LIU P, LIU CS, et al, 2008. The structure characteristics and dominant population regeneration types of Tsuga tchekiangensis communities in the Jiulongshan National Natural Reserve of Zhejiang Province [J]. Acta Ecol Sin, 28(9): 4547-4558 . [张志祥,刘鹏, 刘春生,等, 2008. 浙江九龙山南方铁杉(Tsuga tchekiangensis)群落结构及优势种群更新类型 [J]. 生态学报, 28(9): 4547-4558.]
