张广才岭北部三大硬阔树木生长-气候关系的时空变异
2017-05-25苏金娟王晓春
苏金娟, 王晓春
东北林业大学生态研究中心, 哈尔滨 150040
张广才岭北部三大硬阔树木生长-气候关系的时空变异
苏金娟, 王晓春*
东北林业大学生态研究中心, 哈尔滨 150040
树木年轮在时空尺度上的比较可以更好地反映环境变化对树木生长的影响,在认识气候变化对森林生态系统的影响上具有重要意义。采用树木年代学方法分析了张广才岭北部地区阔叶红松林中主要阔叶树种-水曲柳(Fraxinusmandshurica)、黄菠萝(Phellodendronamurense)和胡桃楸(Juglansmandshurica)径向生长与气候关系的时空变异。结果表明,在同一地点树种间气候响应差异明显,胡桃楸受降水和最低温度共同作用,而黄菠萝和水曲柳则主要受最低温度限制,这表明树木生长与气候因子的关系具有一定的物种特异性。随着温度和降水格局的改变,三大硬阔年轮与气候关系在空间水平上存在差异,方正和西大圈样点的胡桃楸与6—8月最低温度呈显著正相关(P<0.05),而凤凰山样点与5、6月最低温度和降水呈正相关(P<0.05);黄菠萝和水曲柳径向生长随着降水空间格局的变化,其生长季末期相关性程度逐渐减弱。1980年后张广才岭北部出现气温显著升高,在升温前三大硬阔的生长趋势相对一致,而在升温后黄菠萝和水曲柳树轮宽度随温度升高呈上升趋势,但胡桃楸却出现随温度升高而生长下降的“分异现象”。如果未来增温趋势持续发生或者加重,可以推断在张广才岭北部胡桃楸可能受干旱胁迫加剧,其可能出现生长衰退,但增温可能更有利于黄菠萝和水曲柳的生长。
树木年轮;阔叶红松林;干旱胁迫;全球变暖;径向生长
近百年来,地球气候正经历着一次以变暖为主要特征的显著变化。全球变暖的影响在北半球中高纬度地区最为突出,尤其是在内陆生态系统[1- 4]。越来越多的数据表明中国东北地区自1970年以来增温幅度显著提高,这种明显的增温导致从1980年开始进入一种暖干化趋势。这种变暖会在不同尺度上影响森林生态系统的生产力,已有研究表明气候变暖后森林生产力下降[5- 7],也有研究显示气候变暖会改变森林树木的生长趋势[8]、物候和分布地区[9]。张广才岭北部的阔叶红松林,也是气候变化最为显著的地区之一。三大硬阔-水曲柳、黄菠萝和胡桃楸是阔叶红松林中的重要树种,也是红松(Pinuskoraiensis)的主要伴生树种,它们在森林环境形成、红松更新及阔叶红松林的结构与功能动态中起着非常重要的作用。此外,由于树木生理特性的限制,树轮的清晰程度有所不同,阔叶树种树轮研究较针叶树种更为困难,阔叶树种树轮研究相对较少,但是阔叶树种年轮可能含有与针叶树种不一样的信息[10-11]。因此,分析三大硬阔径向生长与气候关系,是探索整个阔叶红松林生态系统对气候变化响应的重要组成部分。
树木年代学在时间和空间尺度上的研究能够为树木径向生长时空变化提供可靠的信息[12]。目前,国外对阔叶树种树轮-气候响应时空变化研究已有一些报道,例如,Huang等[13]发现加拿大美洲山杨(Populustremuloides)和北美白桦(Betulapapyrifera)在南部主要受夏季湿度影响,而在北部主要受温度影响。Pierre等[9]发现随着经度变化,夏季降水减少,温度增加,无梗花栎(Quercuspetraea)对5、6月份温度和降水的相关性也减弱。另外,在中欧和法国西部对栎树以及其它阔叶树种的研究均表明海拔、经度、纬度变化对树木的生长有不同的影响[14- 16]。在国内张寒松等[17]用水曲柳年表重建了长白山过去240年降水变化;而王蔚蔚等[18]发现长白山水曲柳对温度信号更敏感,重建了长白山地区秋季温度;李牧等[19]在敦化地区用红松和三大硬阔年表重建了生长季最低温度。这些研究都集中在空间分析和重建上,然而过去几十年研究表明,在我国青藏高原和东北地区树木生长对气候响应是不稳定的,在气候变暖后出现了明显的转型[20- 22],即树木生长分异现象[23]。那么在全球变暖的背景下在张广才岭北部三大硬阔的树木生长分异现象时空格局会发生变化呢?
本文以张广才岭北部三大硬阔树轮为研究对象,探讨“三大硬阔树轮与气候关系在空间格局上的响应变化以及气候变暖后在时间上是否具有稳定性”这一科学问题,研究结果旨在揭示三大硬阔树轮生长对气候变化的不同响应特点,以期了解未来气候变暖对于森林生态系统的影响。
1 材料和方法
1.1 采样点概况
在黑龙江省张广才岭北部地区沿经度选取3个人为干扰较少的地区(图1):方正县双子山国家森林公园、勃利县西大圈森林公园和鸡东县凤凰山国家级自然保护区,作为本研究的采样点(表1)。研究区域属于长白山系支脉张广才岭北部,该区域属于温带大陆性气候,四季较为分明,雨热同期,年均温2.8—5℃,无霜期100—160 d,年总降水量400—600 mm(图2,表2),土壤多为暗棕色森林土。张广才岭地貌形态复杂,地形以山地为主,自然条件丰富,植物种类众多,植被区系类型以温带分布为主。主要森林植被类型为针阔叶混交林,主要包括白桦(Betulaplatyphylla)林、落叶阔叶林、杂木林、阔叶红松林以及蒙古栎(Quercusmongolica)林等多种类型。乔木树种以红松、黄菠萝、水曲柳、胡桃楸、白桦、紫椴(Tiliaamurensis)和蒙古栎等为主;灌木植物以金银忍冬(Loniceramaackii)、毛榛子(Corylusmandshurica)、胡枝子(Lespedezabicolor)等为优势种;草本植物尖齿蹄盖蕨(Athyriumdentilobum)、小叶芹(Aegopodiumalpestre)、木贼(Equisetumhyemale)等为主。张广才岭北部是东亚针阔叶混交林分布中心,三大硬阔——胡桃楸、黄菠萝、水曲柳广泛分布于张广才岭,是张广才岭北部山区顶极群落(阔叶红松林)的主要阔叶树种,并且采样地区受人为干扰较少,是研究树轮-气候关系的理想区域。
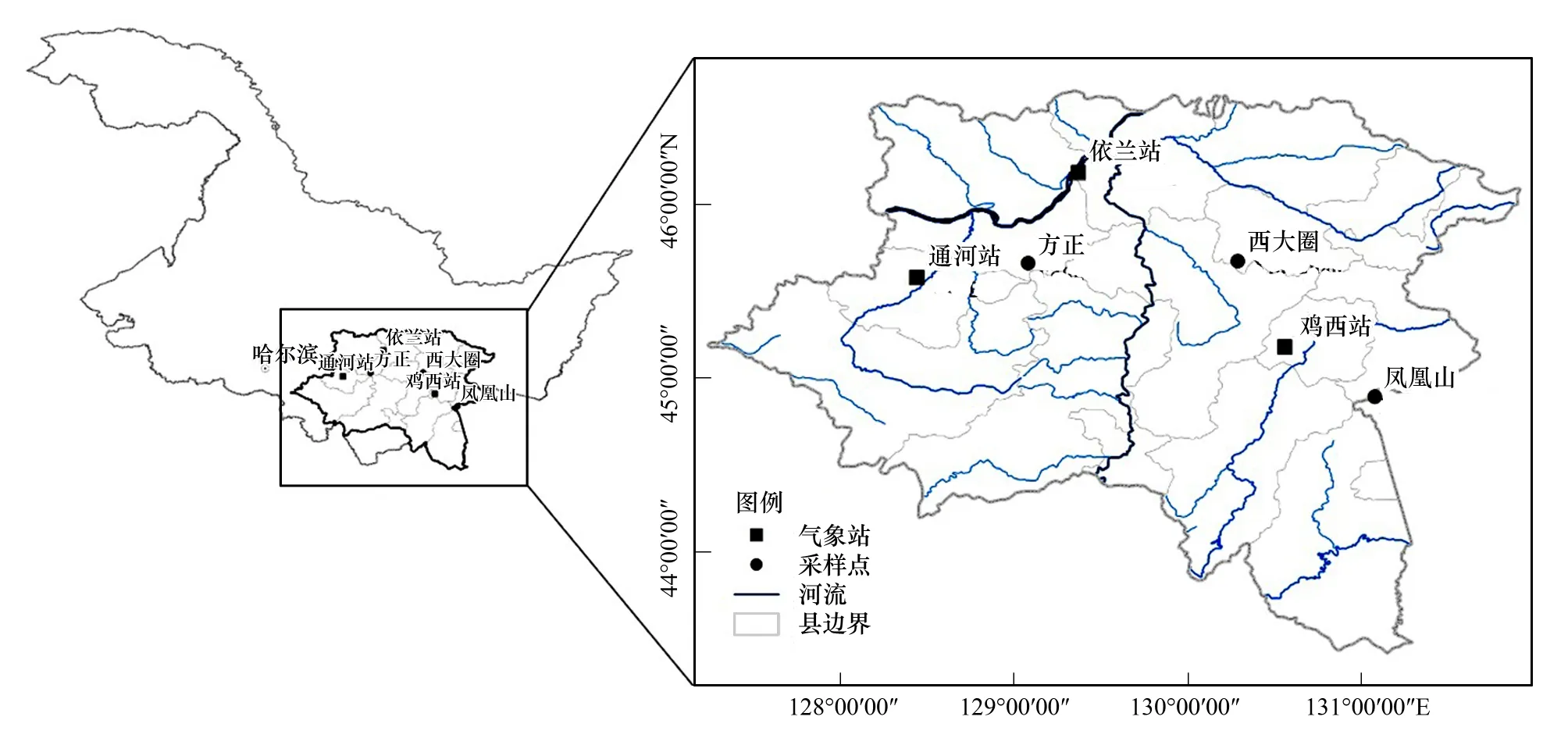
图1 三大硬阔树木年轮采样点和气象站分布Fig.1 Map of sampled sites for three hardwood tree rings and weather station near its sampled site
1.2 样本采集及年表建立

图2 研究地区气象站月平均气温和月总降水量变化 Fig.2 Monthly mean temperature and total precipitation at each study area
2014年7月在方正县双子山国家森林公园(FZ),勃利县西大圈森林公园(XDQ)和鸡东县凤凰山国家级自然保护区(FHS)分别在北坡下坡位进行树轮采样(表1,图1)。选择生长良好的树木,在胸高处(1.3 m)用生长锥钻取树轮样本。将取到的样本放入塑料管内,进行编号,并记录每棵树的生境。每个树种采集15—20棵树,每棵树采集2个树轮样芯。
将样品带回实验室进行固定、干燥、打磨后,用骨架示意图方法进行交叉定年[24],之后用Velmex年轮分析仪进行测量,精确到0.001 mm。测量后用COFECHA程序[25]对交叉定年和测量结果进行检验,以确保其结果准确性。经交叉定年后的树轮序列,利用ARSTAN程序进行去趋势和标准化[26],去趋势方法采用负指数或线性函数进行,降低因年龄生长速率不同而带来的影响。用双重平均法进行年轮曲线标准化,最终建立3个采样点黄菠萝、胡桃楸和水曲柳9个标准年表(图3)用以进行年轮-气候响应分析。
1.3 统计分析
选择离采样较近的通河和鸡西气象站分别对应方正(FZ)和凤凰山(FHS)采样点(表1),由于西大圈与依兰和鸡西气象站距离相等,本研究将依兰站和鸡西站气象站平均作为西大圈气象数据。气象数据由中国气象科学数据共享服务网(http://cdc.cma.gov.cn/)提供(表2)。气候因子主要包括:月均最低温(Tmin)、月均最高温(Tmax)、月均温(T)、月总降水量(P)、相对湿度(RH),其中相对湿度是由空气中的绝对湿度与同温度下的饱和绝对湿度的比值,得出一个百分比。由于上一年气候因子可能影响当年树木径向生长[24],因此分析了上一年10月到当年9月共12个月的气候因子与黄菠萝、胡桃楸和水曲柳标准年表的相关性。由于气候变化对树木径向生长有累积和长期的影响[22],本研究分析了年际、季节与树木径向生长的关系。结合生理生态意义将季节划分为:冬季(前一年12月至当年2月)、春季(3—5月)、夏季(6—8月)、秋季(9—11月)。
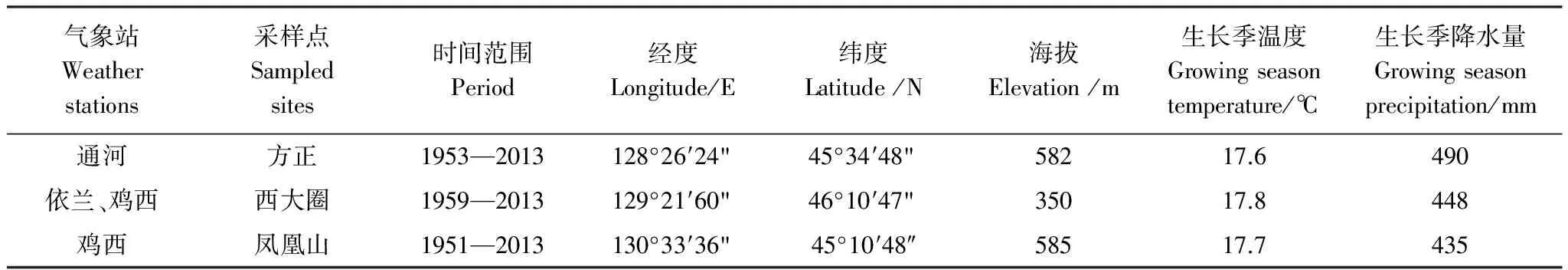
表2 不同采样点对应气象站点信息
根据相关分析结果分别将同一地点水曲柳、胡桃楸和黄菠萝提取同一地点主成分年表和不同地点同一树种的主成分年表用于分析最低温度对不同地点间树种的影响。采用SPSS19.0完成主成分提取及相关统计分析;采用Sigmapolt 12.5等软件完成图表制作。
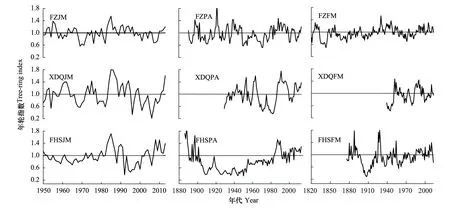
图3 不同地点三大硬阔标准年表年轮指数变化Fig.3 Variations of the standard chronologies for three hardwoods tree species at different sitesFZ、XDQ、FHS分别代表方正、西大圈、凤凰山
2 结果与分析
2.1 年表特征
在9个三大硬阔标准年表中,不同样点间三大硬阔年龄差异较大,其中方正水曲柳的树龄最大(208a),凤凰山胡桃楸的树龄最小(64a)。平均敏感度反映了年轮宽度对气候变化的敏感程度,胡桃楸的平均敏感度普遍高于黄菠萝和水曲柳,这说明了胡桃楸对气候信号更敏感。一阶自相关反映了上一年气候状况对当年年轮宽度生长的影响,一阶自相关都较高,说明了气候变化对三大硬阔生长影响的“滞后效应”较为明显。9个标准年表的样本总体代表性都高于0.85(表3),表明这些样本可以代表种群的总体水平。总体来说,9个标准年表的平均敏感度、信噪比和样本代表性都很高,年表质量较好,这说明这些年表适合用于树木年轮学气候研究。

表3 三大硬阔标准年表特征的主要统计特征
9个年表间在整体上具有较高的一致性,但局部也存在差异(表4)。方正水曲柳和西大圈黄菠萝年表间相关性最高(r=0.64),凤凰山水曲柳与方正胡桃楸和方正水曲柳年表间相关系数均最低(r=-0.03, 0.03),这可能与采样点空间距离、树种有关。另外,同一地点不同树种间的相关系数也存在差异,凤凰山胡桃楸与黄菠萝年表间的相关系数是0.39,而与水曲柳年表的相关为0.15,这表明树木的生理特征差异也会在一定程度影响年表对气候的响应。
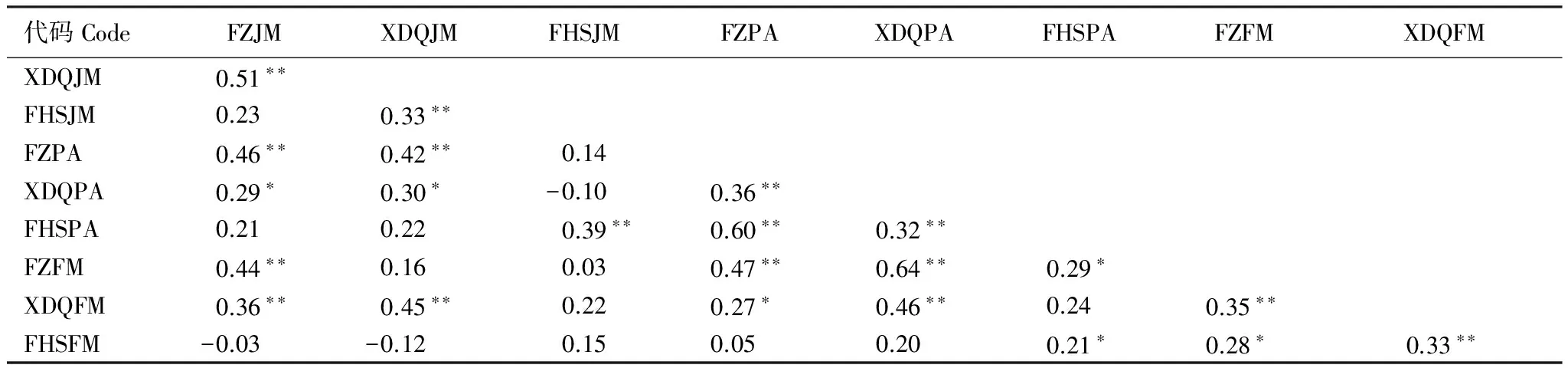
表4 不同地点、树种标准年表间相关系数(公共区间1949—2013年)
2.2 三大硬阔树轮与气候关系的空间变异
从9个三大硬阔标准年表与气候因子相关分析结果来看(图4),不同样点三大硬阔与气候因子响应关系存在差异,方正胡桃楸径向生长与生长季(6—9月)最低温度和降水均呈明显正相关,其中年表序列与6、7月最低温度及6、7、9月降水均呈显著正相关;西大圈胡桃楸与生长季最低温度和降水相关性较弱,仅与9月份降水显著正相关;凤凰山胡桃楸在生长季前(3—5月)及生长季与最低温度均呈正相关,其中5、6月最低温度呈显著正相关,而8、9月降水则呈负相关。黄菠萝和水曲柳对降水相关性不明显,在各个样点均表现较弱程度的正相关,各样点黄菠萝和水曲柳与最低温度呈明显的正相关,且凤凰山样点年表序列与最低温度的相关程度明显高于西大圈和方正样点。

图4 三大硬阔标准年表与月最低温度、降水的相关系数Fig.4 Correlation coefficients between standard tree-ring chronologies and monthly climate variables (monthly mean minimum temperatures and monthly total precipitation) for different sites and tree species-表示前一年

图5 三大硬阔标准年表与高温、均温、相对湿度的相关性Fig.5 Correlation coefficients between the standard tree-ring chronologies and monthly climate variables (monthly mean maximum temperature, mean temperature, and monthly mean relative humidity) for three hardwoods at different sites
从图5可以看出,方正胡桃楸径向生长与4、7月最高温负相关,与生长季相对湿度呈正相关;西大圈胡桃楸径向生长与温度、相对湿度相关性明显相反,与温度呈现一致的负相关,而相对湿度则呈现正相关,其中7—9月相对湿度呈显著正相关;凤凰山胡桃楸与最高温、均温以及相对湿度相关性较弱,仅与7月最高温、相对湿度达到显著水平。不同样点黄菠萝径向生长与最高温、均温、相对湿度差异明显,方正、西大圈样点相对湿度均呈正相关,而凤凰山样点则表现明显的负相关,另外凤凰山样点与均温相关程度明显高于方正、西大圈。3个样点水曲柳径向生长与生长季相对湿度均呈正相关,最高温表现明显不同,方正样点在生长季末期最高温明显正相关,而凤凰山和西大圈则表现较弱的负相关。
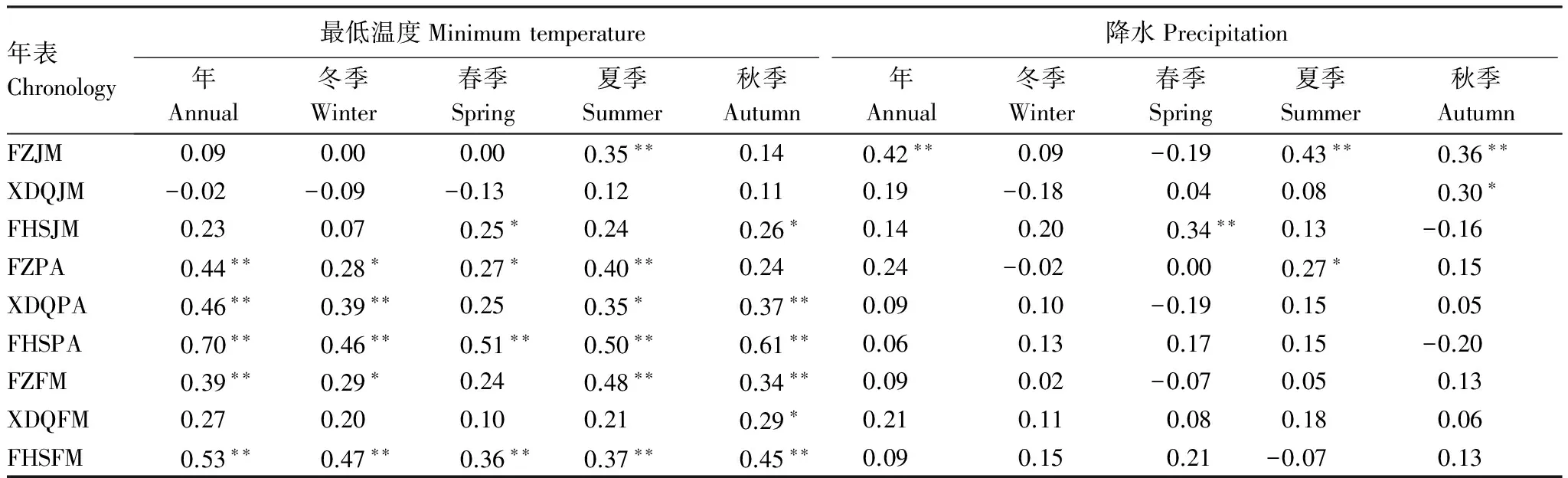
表5 三大硬阔标准年表与季节气候因子相关性
*P<0.05,**P<0.01
从表6中可以看出,方正年际、季节温度和降水对胡桃楸径向生长均呈正相关,其中夏季的最低温度和降水对胡桃楸的径向生长具有极显著的促进作用;西大圈胡桃楸对最低温度和降水则表现较弱程度的正相关,仅秋季的降水达到了显著水平(P<0.05);凤凰山胡桃楸较方正和西大圈对最低温度、降水的相关性程度有所增强,与春季和秋季的最低温度都达到了显著正相关,与春季降水达到极显著水平。3个样点的黄菠萝和水曲柳年际、季节相关结果表明(表6),年最低温度与黄菠萝和水曲柳都达到了极显著正相关(P<0.01),而降水相关性均呈较弱的正相关关系。这说明了最低温度是限制黄菠萝和水曲柳径向生长的主要限制因子。此外3个采样点从东到西降水在减少,其黄菠萝和水曲柳与降水相关性程度随样点变化逐渐减弱。
2.3 三大硬阔树轮-气候关系的时间变异
3个采样点温度和降水变化表明(图6),3个样点在1980年左右发现温度显著升高,1980年前平均最低温度缓慢上升甚至下降,但是1980年后平均最低温度均以0.03℃/a速率持续上升。降水变化则不明显,3个样点1980年前后一致的表现为下降趋势。
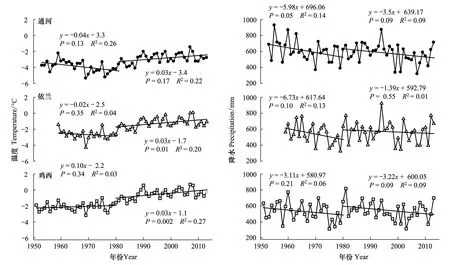
图6 1950—2013年研究区域年最低温度和总降水量变化Fig.6 Variation of annual mean minimum temperature and total precipitation from 1950 to 2013
三大硬阔不同树种升温前后年轮指数变化较大(图7),升温后3个样点的胡桃楸生长速率较升温前均呈下降趋势;黄菠萝和水曲柳年轮指数升温前后变化波动较小,其变化规律基本一致,生长速率持续上升。提取同一地点不同树种的第一主成分年表以及不同地点同一树种第一主成分年表与最低温度关系比较(图8)结果显示,3个样点间年轮指数变化差异不明显,1980年气候变暖后生长速率均是上升趋势,其上升幅度较升温前均有所下降;但是不同树种间差异较大,胡桃楸随温度升高年轮指数下降,而黄菠萝和水曲柳具有与温度升高相一致的趋势。
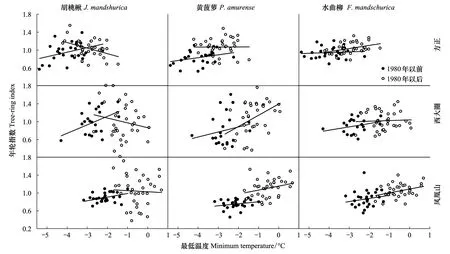
图7 升温(1980年)前后不同地点三大硬阔年轮指数随最低温度变化趋势Fig.7 Changes of tree-ring index for three hardwoods with annual minimum temperature at different sites before and after rapid warming (1980)
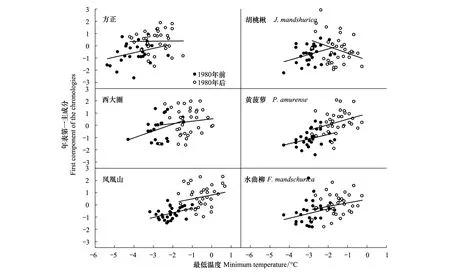
图8 升温(1980年)前后同一地点不同树种以及不同地点相同树种第一主成分随最低温度变化趋势Fig.8 Changes of the first components of chronologies for the same tree species and sites with annual mean minimum temperature before and after rapid warming (1980)
3 讨论
3.1 气候因子和空间环境变化对三大硬阔径向生长的影响
三大硬阔径向生长主要受温度和降水限制,最低温度是水曲柳和黄菠萝径向生长的主要限制因子,胡桃楸径向生长受最低温度和降水共同限制,这与敦化地区三大硬阔研究结果一致[19]。不同树种径向生长响应差异可能与其遗传特性有关,水曲柳具有一定的耐阴性,喜湿润环境并且光合速率最强[27-28],而温度对树木生理生态的最直接影响是改变光合速率,故水曲柳受最低温度的限制作用最强。从对水分需求的生理特性来看,胡桃楸耗水量和萎蔫系数均大于黄菠萝和水曲柳[28-29],并且水曲柳和黄菠萝能够适应一定的干旱胁迫,而胡桃楸在干旱胁迫中不能生长[30-31]。因此,胡桃楸对于水分的限制作用大于黄菠萝和水曲柳。彭剑锋等[32]在天山地区对天山云杉(Piceaschrenkiana)和西伯利亚落叶松(Larixsibirica)的研究发现遗传特性是造成树木对同一生境响应不同的原因。
张广才岭北部属于湿润、半湿润地区,但由于地理空间位置的差异,3个样点的环境条件并不完全一致,其不同样点温度、降水与树木径向生长响应有所差异。方正样点胡桃楸与生长季6、7、9月最低温度、降水显著正相关。方正样点位于最西端,气候较湿润,温度和降水共同调控胡桃楸径向生长,6—9月是树木生长最旺盛时期,良好的水热条件能够促进胡桃楸快速生长,体现了胡桃楸对水热条件的同步正响应。西大圈样点胡桃楸与生长季最低温度、降水相关程度减弱,仅与9月最低温度、降水呈显著正相关,主要是由于西大圈位于中间,其气候条件较温暖,加剧了水分的蒸发,此外9月份是生长季末期,过多的降水不利于营养物质的积累以及木质化的进程,导致水分对胡桃楸的促进作用下降,两者对胡桃楸径向生长的正响应均降低。凤凰山胡桃楸与最低温度和降水显著相关性则出现在5、6月,这可能与凤凰山地理位置更靠东部有关,气候条件为暖干,5月处于生长期早期,树木对水分的需求量大,较高的温度加速了冰雪融化和土壤温度升温,促进了根系的活动和地上部分的萌发,从而有利于生长季早期树木生长,故凤凰山胡桃楸提前表现显著的正相关。黄菠萝和水曲柳与降水相关性程度在生长季随样点变化逐渐减弱,这是因为在半湿润地区, 高温可以加快植物蒸腾和土壤蒸发, 加剧水分胁迫作用, 从而很大程度上抑制树体的生长,当降水量低于某一值时, 温度与树木生长呈现明显负相关, 降水量越少, 这种关系越明显与桑卫国[33]等对天山云杉研究结果一致。这似乎可以说明三大硬阔径向生长随着样点(经度)的变化,不同的水热条件对其径向生长影响有一定变化,但具体机制有待于进一步研究。
3.2 气候变暖对三大硬阔年轮生长的影响
20世纪80年代东北地区温度显著升高[34- 36],其中最低气温增高更明显[37],最低温升温对植物的生长影响是复杂的,有研究证明最低温度升高对植物生长有促进作用[38- 40],也有人认为最低温度升高对植物生长有抑作用[7]。最低温度升高对三大硬阔径向生长在不同树种间的差异明显。1980年前三大硬阔生长趋势基本一致(图7),1980年后黄菠萝和水曲柳径向生长随着温度升高而上升,但胡桃楸在升温后并未随温度升高而呈现上升趋势,反而出现下降趋势,胡桃楸的主成分年表结果也呈现这一趋势,胡桃楸径向生长与最低温度升高出现了“分异现象”。响应分异现象广泛存在于北半球中高纬度地区[20-21,41]。胡桃楸径向生长受降水和最低温度共同作用,最低温度升高加快植物蒸腾作用和土壤蒸发,从而导致干旱胁迫抑制胡桃楸径向生长。从胡桃楸年轮与气候相关分析可以推测,升温造成的干旱胁迫可能是胡桃楸出现分异现象主要原因,而且随着自西向东降水增多,其响应分异程度也随着经度梯度逐渐减弱,这似乎证明了这一结论。这与长白山鱼鳞云杉(Piceajezoensisvar.komarovii)上限树轮与温度出现的分异现象一致[39]。李宗善等[42]对川西地区岷江冷杉(Abiesfaxoniana)研究发现日照时数的变化是引起分异现象的原因,川西地区属于青藏高原气候,海拔高、空气稀薄、日照时数长对植物的影响较大。黄菠萝和水曲柳主要受最低温度限制,降水对其影响较小,升温导致的干旱加剧可能没有成为主要的限制因素,即使夏季最低温度有一定程度上升,可能目前并没有超过这两种树种的温度界限,因此并没有发生显著的变化,尚华明等[43]也证明了这一结论。另外,最低温度升高没有影响到黄菠萝和水曲柳的同时,也可能降低了其对环境变化的敏感性,即增强了其对高温胁迫的抵抗力,使其在显著升温后没有出现响应分异现象。虽然1980年后的显著升温,黄菠萝和水曲柳径向生长没有显著变化,但是如果持续变暖,尤其是夏季最低温度升高,超出其耐受温度或者引起干旱胁迫,也会对其生长趋势有一定影响。
4 结论
张广才岭北部阔叶红松林是气候变化最为显著的地区之一,本研究通过对阔叶红松林中水曲柳、黄菠萝和胡桃楸径向生长与气候关系分析表明:在空间尺度上,方正生长季同步的水热条件促进了胡桃楸径向生长,西大圈胡桃楸在生长季与最低温度、降水则表现较弱的正相关,凤凰山胡桃楸与5、6月最低温度、降水呈现正相关。黄菠萝和水曲柳径向生长与经度变化吻合,随着从西向东降水增多,其年表序列与降水的正相关作用逐渐减弱。在时间尺度上,20世纪80年代温度显著升高后,三大硬阔的径向生长在不同树种间出现明显差异,黄菠萝和水曲柳在显著升温后生长趋势没有明显变化,但胡桃楸生长趋势出现与升温相反的“分异现象”。未来在持续变暖的环境下胡桃楸可能出现衰退,而升温对黄菠萝和水曲柳的影响不大。三大硬阔与气候关系的时空变异机理尚不完全清楚,将来可通过三大硬阔的树轮解剖学或生理生态响应实验进一步分析其时空变异机制。
[1] Serreze M C, Walsh J E, Chapin III F S, Osterkamp T, Dyurgerov M, Romanovsky V, Oechel W C, Morison J, Zhang T, Barry R G. Observational evidence of recent change in the northern high-latitude environment. Climatic Change, 2000, 46(1/2): 159- 207.
[2] Andreu L, Gutiérrez E, Macias M, Ribas M, Bosch O, Camarero J J. Climate increases regional tree-growth variability in Iberian pine forests. Global Change Biology, 2007, 13(4): 804- 815.
[3] Diego Galván J, Büntgen U, Ginzler C, Grudd H, Gutiérrez E, Labuhn I, Camarero J J. Drought-induced weakening of growth-temperature associations in high-elevation Iberian pines. Global and Planetary Change, 2015, 124: 95- 106.
[4] IPCC. Summary for policymakers of climate change 2013: the physical science basis//Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Xia Y, Bex V, Midgley P M. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2014.
[5] 朱建华, 侯振宏, 张治军, 罗云建, 张小全. 气候变化与森林生态系统: 影响、脆弱性与适应性. 林业科学, 2007, 43(11): 138- 145.
[6] 刘国华, 傅伯杰. 全球气候变化对森林生态系统的影响. 自然资源学报, 2001, 16(1): 71- 78.
[7] Clark D A, Piper S C, Keeling C D, Clark D B. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984- 2000. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(10): 5852- 5857.
[8] D′Arrigo R D, Kaufmann R K, Davi N, Jacoby G C, Laskowski C, Myneni R B, Cherubini P. Thresholds for warming-induced growth decline at elevational tree line in the Yukon Territory, Canada. Global Biogeochemical Cycles, 2004, 18(3): GB3021, doi: 10.1029/2004GB002249.
[9] Mérian P, Bontemps J D, Bergès L, Lebourgeois F. Spatial variation and temporal instability in climate-growth relationships of sessile oak (Quercuspetraea[Matt.] Liebl.) under temperate conditions. Plant Ecology, 2011, 212(11): 1855- 1871.
[10] 高露双, 王晓明, 赵秀海. 长白山阔叶红松林共存树种径向生长对气候变化的响应. 北京林业大学学报, 2013, 35(3): 24- 31.
[11] Takahashi K, Okuhara I. Comparison of climatic effects on radial growth of evergreen broad-leaved trees at their northern distribution limit and co-dominating deciduous broad-leaved trees and evergreen conifers. Ecological Research, 2012, 27(1): 125- 132.
[12] Tardif J, Camarero J J, Ribas M, Gutiérrez E. Spatiotemporal variability in tree growth in the Central Pyrenees: climatic and site influences[J]. Ecological Monographs, 2003, 73(2): 241- 257.
[13] Huang J G, Tardif J C, Bergeron Y, Denneler B, Berninger F, Girardin M P. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Global Change Biology, 2010, 16(2): 711- 731.
[14] Lara A, Villalba R, Wolodarsky-Franke A, Aravena J C, Luckman B H, Cuq E. Spatial and temporal variation inNothofaguspumiliogrowth at tree line along its latitudinal range (35°40′- 55°S) in the Chilean Andes. Journal of Biogeography, 2005, 32(5): 879- 893.
[15] Friedrichs D A, Trouet V, Büntgen U, Frank D C, Esper J, Neuwirth B, Löffler J. Species-specific climate sensitivity of tree growth in Central-West Germany. Trees, 2009, 23(4): 729- 739.
[17] 张寒松, 韩士杰, 李玉文, 张军辉. 利用树木年轮宽度资料重建长白山地区240年来降水量的变化. 生态学杂志, 2007, 26(12): 1924- 1929.
[18] 王蔚蔚, 张军辉, 戴冠华, 王秀秀, 韩士杰, 张寒松, 王云. 利用树木年轮宽度资料重建长白山地区过去240年秋季气温的变化. 生态学杂志, 2012, 31(4): 787- 793.
[19] 李牧, 王晓春. 敦化三大硬阔、红松年轮-气候关系及生长季低温重建. 南京林业大学学报:自然科学版, 2013, 37(3): 29- 34.
[20] Zhang Y X, Wilmking M. Divergent growth responses and increasing temperature limitation of Qinghai spruce growth along an elevation gradient at the northeast Tibet Plateau. Forest Ecology and Management, 2010, 260(6): 1076- 1082.
[21] Zhang Y X, Wilmking M, Gou X H. Changing relationships between tree growth and climate in Northwest China. Plant Ecology, 2009, 201(1): 39- 50.
[22] Yu D P, Liu J Q, Benard J L, Zhou L, Zhou W M, Fang X M, Wei Y W, Jiang S W, Dai L M. Spatial variation and temporal instability in the climate-growth relationship of Korean pine in the Changbai Mountain region of Northeast China. Forest Ecology and Management, 2013, 300: 96- 105.
[23] Briffa K R, Schweingruber F H, Jones P D, Osborn T J, Shiyatov, S G, Vaganov E A. Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature, 1998, 391(6668): 678- 682.
[24] Fritts H C. Relationships of ring widths in arid-site conifers to variations in monthly temperature and precipitation. Ecological Monographs, 1974, 44(4): 411- 440.
[25] Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69- 78.
[26] Cook E, Holmes R L. User Manual for Program ARSTAN, Laboratory of Tree-Ring Research. Tucson: University of Arizona, 1986.
[27] 马建路. 水曲柳、胡桃楸、黄波罗和紫椴适生立地条件的对比研究. 林业资源管理, 1996, (4): 52- 56.
[28] 王晶英, 王展, 赵雨森. 水曲柳等树种光合、耗水及萎蔫系数比较研究. 林业科技, 2010, 35(3): 10- 13.
[29] 孙慧珍, 孙龙, 王传宽, 周晓峰. 东北东部山区主要树种树干液流研究. 林业科学, 2005, 41(3): 36- 42.
[30] 梁淑娟, 潘攀, 孙志虎, 王庆成. 坡位对水曲柳及胡桃楸生长的影响. 东北林业大学学报, 2005, 33(3): 18- 19.
[31] 肖冬梅, 王淼, 姬兰柱. 水分胁迫对长白山阔叶红松林主要树种生长及生物量分配的影响. 生态学杂志, 2004, 23(5): 93- 97.
[32] 彭剑峰, 勾晓华, 陈发虎, 李金豹. 天山云杉和西伯利亚落叶松的树轮气候记录. 生态环境, 2005, 14(4): 460- 465.
[33] 桑卫国, 王云霞, 苏宏新, 陆兆华. 天山云杉树轮宽度对梯度水分因子的响应. 科学通报, 2007, 52(19): 2292- 2298.
[34] 王绍武, 赵宗慈, 唐国利. 中国的气候变暖. 国际政治研究, 2009, (4): 1- 11, 194- 194.
[35] 周秀杰, 王凤玲, 吴玉影, 那济海, 潘华盛, 王勇. 近60年来黑龙江省与东北及全国气温变化特点分析. 自然灾害学报, 2013, 22(2): 124- 129.
[36] 李腾, 何兴元, 陈振举. 东北南部蒙古栎径向生长对气候变化的响应——以千山为例. 应用生态学报, 2014, 25(7): 1841- 1848.
[37] 孙凤华, 杨修群, 路爽, 杨素英. 东北地区平均、最高、最低气温时空变化特征及对比分析. 气象科学, 2006, 26(2): 157- 163.
[38] 于大炮, 王顺忠, 唐立娜, 代力民, 王庆礼, 王绍先. 长白山北坡落叶松年轮年表及其与气候变化的关系. 应用生态学报, 2005, 16(1): 14- 20.
[39] 李广起, 白帆, 桑卫国. 长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应. 植物生态学报, 2011, 35(5): 500- 511.
[40] 王淼, 白淑菊, 陶大立, 单建平. 大气增温对长白山林木直径生长的影响. 应用生态学报, 1995, 6(2): 128- 132.
[41] Driscoll W W, Wiles G C, D′Arrigo R D, Wilmking M. Divergent tree growth response to recent climatic warming, Lake Clark National Park and Preserve, Alaska. Geophysical Research Letters, 2005, 32(20): L20703, doi: 10.1029/2005GL024258.
[42] 李宗善, 刘国华, 傅伯杰, 张齐兵, 胡婵娟, 罗淑政. 川西卧龙国家级自然保护区树木生长对气候响应的时间稳定性评估. 植物生态学报, 2010, 34(9): 1045- 1057.
[43] 尚华明, 魏文寿, 袁玉江, 喻树龙, 张同文, 瓦合提·艾则买提, 李新建. 阿尔泰山南坡树轮宽度对气候变暖的响应. 生态学报, 2010, 30(9): 2246- 2253.
Spatio-temporal variations in climate-growth relationships of three hardwood tree species across the north Zhangguangcai Mountains, northeast China
SU Jinjuan, WANG Xiaochun*
CenterforEcologicalResearch,NortheastForestryUniversity,Harbin150040,China
Forests play a critical role in the global ecological balance. One of the major uncertainties in global climate change predictions is how the spatio-temporal dynamics of forests will be affected by global warming. Tree rings at different spatial and temporal scales can reflect the impacts of environmental change on tree growth, and can be critical for understanding the influence of climate change on forest ecosystems. Here, we investigated the spatio-temporal patterns of variation in climate-growth relationships of three common broadleaved tree species (Fraxinusmandshurica,Phellodendronamurense, andJuglansmandshurica) in a broadleaved Korean pine mixed forest across the north Zhangguangcai Mountains, northeast China. Tree-ring chronologies from three sample sites were established, transformed into principal component analysis (PCA), and analyzed through correlation analysis. With these analyses, climate factors limiting the radial growth were identified, as well as the detailed spatial radial growth-climate associations. The responses of the tree species at similar sites to climate factors differed significantly. Radial growth ofJ.mandshuricawas limited by monthly total precipitation and minimum temperature, whereasP.amurenseandF.mandshuricawere mainly affected by monthly mean minimum temperature. This suggests that climate-growth relationships are affected by species-specific characteristics. With changes in temperature and precipitation patterns, the three hardwoods differed in their spatial growth response to climate factors. The radial growth ofJ.mandshuricain Fangzheng and Xidaquan was positively correlated with minimum June to August temperatures (P<0.05). In contrast, it was positively correlated in Fenghuangshan with minimum temperature and precipitation in May and June (P<0.05). The radial growth ofP.amurenseandF.mandshuricadecreased from west to east with precipitation changes, since the correlation coefficients between tree-ring index and precipitation decreased from west to east. Temperatures in the north Zhangguangcai Mountains have increased since 1980, and the radial growth of the three hardwoods was consistent before 1980. Because climate factors affect growth differently, these boreal tree species showed inconsistent spatio-temporal responses to the recent warming, withJ.mandshuricabeing probably the most sensitive species. Radial growth ofP.amurenseandF.mandshuricaincreased with rapid warming since 1980, whereas tree growth ofJ.mandshuricadecreased after 1980. Therefore,J.mandshuricamay be subjected to drought stress and a subsequent decline in radial growth in the north Zhangguangcai Mountains if global warming continues or exacerbates. In contrast,P.amurenseandF.mandshuricamay benefit from increased temperatures through enhanced radial growth. For future research, the spatio-temporal variation of growth-climate relationship of the three hardwoods could be explored further by investigating the tree ring anatomical and physiological responses.
tree rings; broadleaved Korean pine mixed forest; drought stress; global warming; radial growth
国家自然科学基金面上项目(41471168, 31370463);教育部新世纪优秀人才支持计划(NCET- 12-0810);黑龙江省留学归国科学基金(LC2012C09)共同资助
2015- 09- 25;
日期:2016- 07- 13
10.5846/stxb201509251973
*通讯作者Corresponding author.E-mail: wangxc-cf@nefu.edu.cn
苏金娟, 王晓春.张广才岭北部三大硬阔树木生长-气候关系的时空变异.生态学报,2017,37(5):1484- 1495.
Su J J, Wang X C.Spatio-temporal variations in climate-growth relationships of three hardwood tree species across the north Zhangguangcai Mountains, northeast China.Acta Ecologica Sinica,2017,37(5):1484- 1495.
