水稻斑点叶突变体splZ97的生理特性及其基因定位
2017-05-24韦荔全罗延敏王文强池长程黄福灯程方民
韦荔全 罗延敏 王文强 池长程 黄福灯 向 珣 程方民 潘 刚,*
水稻斑点叶突变体spl的生理特性及其基因定位
韦荔全1,**罗延敏1,**王文强1池长程1黄福灯2向 珣1程方民1潘 刚1,*
1浙江大学农业与生物技术学院, 浙江杭州310058;2浙江省农业科学院作物与核技术利用研究所, 浙江杭州310021
通过EMS诱变籼稻恢复系珍97获得一个稳定遗传的褐色斑点叶突变体spl(ottedeaf, spl)。大田条件下, 突变体spl的斑点叶性状始于分蘖期, 此后由叶缘向叶中下部迅速扩散, 直至整个叶片, 严重时叶片部分或整体枯死, 从而致使突变体株高、每穗粒数及结实率极显著低于野生型对照。生理分析表明, 与野生型珍97相比, 孕穗期突变体spl剑叶、倒二叶和倒三叶的叶绿素含量极显著降低, 而POD (peroxidase, POD)活性、O2܋含量及MDA (malondialdehyde, MDA)含量升高; 突变体spl倒二叶和倒三叶的CAT (catalase, CAT)活性和可溶性蛋白含量极显著降低, 而SOD (superoxide dismutase, SOD)活性则极显著增加。组织化学分析进一步证实, 突变体spl的叶片明显累积O2܋。此外, 突变体spl苗期经盐胁迫处理后, 其株高及根长明显受到抑制。遗传分析表明, 突变体spl的斑点叶性状受一对隐性核基因控制, 借助图位克隆技术将该基因定位于第12染色体长臂的RM28466与RM28485两个SSR标记之间, 物理距离为189 kb, 该结果为进一步克隆SPL基因并研究其功能奠定了基础。
水稻;spl; 斑点叶; 生理特性; 基因定位
植物斑点叶(spotted-leaf)、类病变(lesion mimic)或类病斑(lesion simulating disease)突变体是指突变体植物在没有受到明显逆境、机械和农药损伤或病原菌侵染的条件下, 植物叶片、叶鞘甚至茎秆等部位自发形成类病原菌侵染或类过敏反应的程序性细胞死亡现象[1-2]。研究表明, 斑点叶突变体还与植物抗病防卫反应密切相关[3]。因此, 斑点叶突变体是研究植物程序性细胞死亡及病害响应机理的理想材料[2-3]。
关于斑点叶的形成机理, 基于不同突变体的分子生理生化研究, 认为斑点叶的形成不仅受外因, 如光[4]和温度[5]的诱导; 更受内因, 如激素和ROS (reactive oxygen species, ROS)等信号因子[6-8], 以及基因的精细调控[7-8]。利用正向和反向遗传学手段, 迄今已从水稻中克隆并经遗传转化或T-DNA插入突变体验证的叶片类病变形成的相关基因有18个, 根据基因功能可以将其分成6类, 包括转录因子, 如[9]和[10]; 蛋白酶或激酶基因, 如/[11][5]; 参与代谢加工降解的基因, 如[12]、/[13]、/[14]、[15]、/[16]、[17]、[18]、[19]等; 物质转运相关基因, 如[20]、[21]、[22]等; 蛋白或RNA结合蛋白基因, 如[23]和[24]; 生物合成相关基因, 如[4]。
本课题组利用EMS诱变籼稻恢复系珍97, 获得一个隐性斑点叶突变体, 暂命名为spl(spotted leaf Z97, spl)。该突变体斑点叶表型始于水稻分蘖期(约6~7叶期), 之后随着叶龄的增加, 除上部1~2片展开叶及心叶外, 其他叶片均不同程度出现斑点叶性状; 抽穗开花期除剑叶外所有叶片均出现斑点叶性状。本文对突变体spl基本表型、生理变化及基因定位等方面的研究, 为进一步克隆该基因并揭示其斑点叶分子生理机理奠定了基础。
1 材料与方法
1.1 试验材料
EMS诱变籼稻恢复系珍97, 从诱变后代中获得斑点叶突变体spl, 经浙江杭州和海南连续6代回交和自交, 获得斑点叶性状稳定株系。之后以spl为母本, 分别与原始野生型对照珍97、粳稻品种02428和秀水110杂交获得F1, F1自交获得的F2群体用于遗传分析及基因定位。所有材料均种植于浙江大学紫金港农业试验站。2014年和2015年在水稻成熟后, 分别取spl与珍97各20株, 调查其株高、穗长、有效穗数、每穗粒数、结实率和千粒重等主要农艺性状。2015年在水稻生长至孕穗期, 取突变体spl及其野生型珍97的剑叶、倒二叶和倒三叶, 测定有关生理指标; 同时, 对F1及其F2群体分单株观察其斑点叶表型并取样, 提取DNA用于后续基因定位分析。
1.2 盐处理
选取突变体spl及其野生型对照珍97的成熟种子并脱壳, 经75%酒精消毒1 min及10%次氯酸钠溶液消毒20 min, 之后灭菌水洗涤5次, 每次2 min。将消毒好的种子接种在含0、100和200 mmol L–1NaCl的1/2 MS[25]固体培养基上, 于水稻组织培养室培养6 d后统计根长及株高并拍照。培养条件为光照16 h, 黑暗8 h, 温度为28℃。
1.3 生理指标测定
选取突变体spl及其野生型对照珍97各10株处于孕穗期(剑叶叶枕与倒二叶叶枕平齐的分蘖)的剑叶、倒二叶和倒三叶, 分别测定叶绿素、可溶性蛋白、过氧化氢(H2O2)、超氧阴离子(O2܋)、MDA含量以及CAT、SOD和POD活性。用80%丙酮于黑暗条件下浸泡叶片至发白, 将浸提液稀释5倍, 以80%丙酮为空白, 在波长663 nm和646 nm下测定其光密度, 计算叶绿素含量; 采用考马斯亮蓝G-250染色法测定可溶性蛋白; 采用碧云天生物技术有限公司的过氧化氢检测试剂盒(s0038)测定H2O2含量; 参考《植物生理学实验技术》的相关方法测定其他生理指标[26]。采用日本岛津公司UV-2450紫外分光光度计, 每个指标测定5个生物学重复。
1.4 细胞组织化学染色
选取分蘖盛期突变体spl及其野生型对照珍97主茎倒二叶中上部3~5 cm长叶片, 分别参考Kariola等[27]和Mahalingam等[28]的方法进行NBT及DAB染色。每个试验重复3次。
1.5 基因遗传分析与定位
大田栽培条件下, 首先分别观察spl/珍97、spl/02428和spl/秀水110的杂种F1叶片表型, 确定控制突变体spl斑点叶性状的显隐性; 其次, 根据F2群体中具有正常叶色的水稻单株数与具有斑点叶表型的单株数的比例, 确定控制斑点叶性状的基因数量。同时, 分单株剪取spl/02428的F2定位群体中具有斑点叶性状的单株、spl、02428以及F2群体中10株正常植株的叶片, 采用CTAB法提取基因组总DNA。利用均匀分布于水稻12条染色体的500对SSR标记, 其引物序列来自Gramene数据库(http://www.gramene.org/), 以及Shen等[29]所开发的50对InDel标记进行基因定位。PCR总反应体系为20 μL, 内含0.8 UDNA聚合酶, 1 × PCR buffer (Mg2+Plus), 1 mmol L–1dNTP Mixture, 50 ng DNA, 上下游引物各0.25 μmol L–1。PCR反应条件为95℃预变性5 min; 94℃, 30 s; 55℃, 30 s; 72℃, 30 s; 共35个循环; 72℃, 10 min。PCR产物经8%非变性聚丙烯酰胺凝胶电泳, 快速银染后观察[30]。
1.6 遗传图谱构建
从spl/02428 F2群体中的853个单株中获得207株具有斑点叶性状的单株组成基因定位群体, 利用上述筛选到的基因连锁分子标记进行定位群体的遗传分析, 获得每个标记的重组交换单株数。同时在RAP-DB数据库(http://rapdb.dna.affrc.go.jp/)查找基因连锁分子标记在染色体上的具体物理位置以确定其排列顺序, 结合每个标记的重组交换单株数确定SPL基因与连锁标记的顺序, 构建基因定位图谱。
2 结果与分析
2.1 突变体spl的表型及主要农艺性状
突变体spl叶片在分蘖期(6~7叶期)就表现斑点叶性状(图1-A), 而后除上部1~2片展开叶及心叶外, 其他叶片的褐色斑点始于叶尖及边缘, 并向叶中部及下部迅速蔓延, 严重时叶中上部枯死。而且自分蘖开始至抽穗期, 相对于野生型珍97, 突变体spl叶片的叶色明显更黄(图1-B, C)。突变体spl生长至抽穗开花前期, 除剑叶外所有叶片均不同程度出现斑点叶症状(图1-D, E, F)。由于叶片出现类病变症状, 导致突变体spl的主要农艺性状, 如株高、穗长、每穗粒数和结实率分别比野生型对照珍97下降25.16%、12.78%、47.29%和42.60% (表1)。
2.2 突变体spl对盐胁迫的响应特性
正常条件下, 突变体spl与野生型珍97之间的株高和根长均无明显差异; 而在盐胁迫条件下, 两者的均受到抑制, 但与野生型相比, 突变体spl的根长和株高抑制更明显(图2-A)。与突变体spl对照相比, 经100 mmol L–1NaCl处理后的突变体株高和根长分别降低19.70%和21.76%; 而经200 mmol L–1NaCl处理的则分别下降26.71%和56.68% (图2-B, C)。
A: 分蘖期; B: 抽穗期; C: 抽穗期叶片; D: 开花期; E: 开花期叶片; F: 开花期倒二叶; 1~4代表剑叶至倒四叶。Bar=20 cm。
A: tillering stage; B:heading stage; C:leaves at heading stage; D:flowering stage; E:leaves at flowering stage; F: the 2nd leaf from top at the flowering stage; 1–4 mean the 1st leaf to 4th leaf from top. Bar=20 cm.
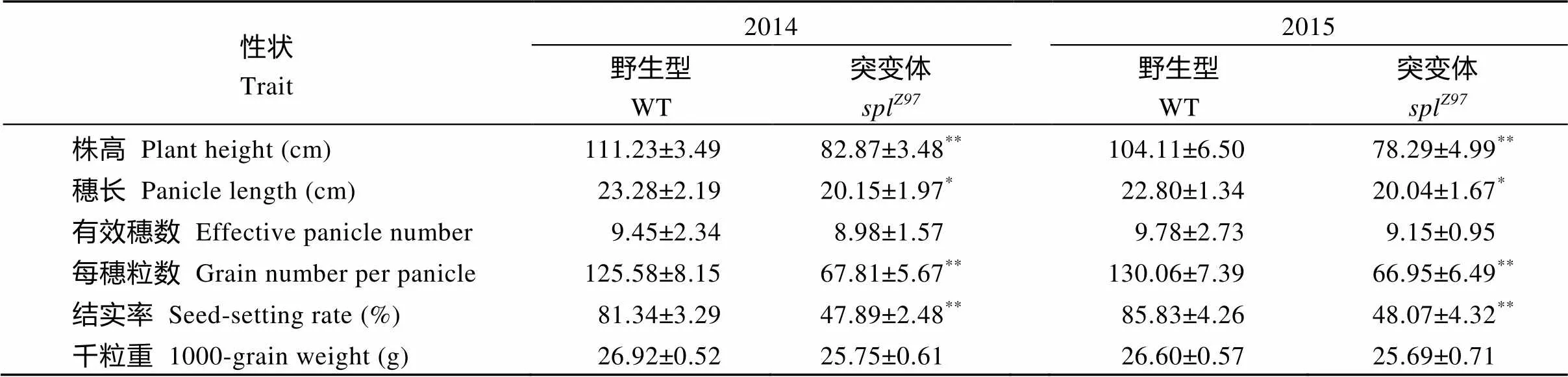
表1 突变体及其野生型的农艺性状
*在0.05水平上差异显著;**在0.01水平上差异显著。
*Significantly different at< 0.05;**significantly different at< 0.01 (-test).
A: 突变体spl及其野生型的表型, Bar=5 cm; B: 芽长; C: 根长。*在0.05水平上差异显著;**在0.01水平上差异显著。
A: phenotype ofspland its WT plants, Bar=5 cm; B: Shoot lengths; C: primary root lengths of seedlings.*Significantly different at<0.05;**Significantly different at<0.01 (-test).
2.3 突变体spl的生理分析
2.3.1 叶绿素含量 孕穗期突变体spl的剑叶、倒二叶和倒三叶的叶绿素(图3-A)、叶绿素(图3-B)、叶绿素总含量(图3-C)及叶绿素/比值(图3-D)依次下降且显著低于野生型, 其中突变体spl的倒二叶和倒三叶的总叶绿素含量分别比其剑叶下降7.47%和39.46% (图3-C)。与野生型相比, 突变体spl的剑叶、倒二叶和倒三叶的总叶绿素含量分别下降43.18%、47.62%和58.59% (图3-C)。
2.3.2 POD、CAT和SOD的活性 图4-A显示, 相对于野生型对照珍97, 突变体spl的剑叶、倒二叶与倒三叶的POD活性均极显著增加, 分别为113.49%、982.69%和929.22%。图4-B显示, 野生型对照珍97的剑叶与倒二叶间的CAT活性无显著性差异, 但倒三叶则分别比剑叶与倒二叶下降11.72%和8.58%; 而突变体spl的剑叶、倒二叶与倒三叶间的CAT活性则依次极显著下降, 且其倒二叶与倒三叶的CAT活性极显著低于野生型珍97, 分别下降21.16%和37.00%。图4-C显示, 突变体spl的剑叶、倒二叶与倒三叶间的SOD活性无显著性差异, 而野生型珍97则依次下降。与野生型珍97相比, 突变体spl的倒二叶与倒三叶的SOD活性分别增加14.75%和23.13%。
1: 剑叶; 2: 倒二叶; 3: 倒三叶。*在0.05水平上差异显著;**在0.01水平上差异显著。
1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top.*Significantly different at<0.05;**significantly different at<0.01 (-test).
1: 剑叶; 2: 倒二叶; 3: 倒三叶。**在0.01水平上差异显著。
1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top.**Significantly different at<0.01 (-test).
2.3.3 超氧阴离子(O2܋)和过氧化氢(H2O2)的累积
图5-A显示, 野生型对照珍97和突变体spl倒二叶和倒三叶间的O2܋含量差异不显著, 但显著高于剑叶。与野生型相比, 突变体spl剑叶、倒二叶和倒三叶的O2܋含量分别增加15.71%、17.31%和13.66%。图5-B结果显示, 野生型及突变体的剑叶、倒二叶和倒三叶间的H2O2含量无显著差异。同时, 抽穗期突变体spl剑叶的部分组织细胞明显被NBT染成更多更深的蓝色(图5-C), 进一步证实其体内累积O2܋; 而突变体spl及其野生型对照叶片组织均没有被DAB染成明显的棕红色(图5-D), 证实两者之间的H2O2含量无明显差异。
2.3.4 MDA和可溶性蛋白含量 图6-A显示, 与野生型相比, 突变体剑叶、倒二叶和倒三叶的MDA含量分别增加29.63%、34.48%和56.25%, 达极显著水平; 且突变体spl倒二叶和倒三叶的MDA含量分别比剑叶增加11.42%和42.86%。图6-B显示, 野生型和突变体剑叶、倒二叶和倒三叶间的可溶性蛋白含量依次降低, 尤其是两者倒三叶的含量极显著低于其剑叶和倒二叶(图6-B), 与野生型相比, 除剑叶外, 突变体倒二叶和倒三叶的可溶性蛋白含量分别下降12.26%和16.74%。
2.4 遗传分析及基因定位
spl/珍97、spl/02428和spl/秀水110三个杂种F1单株的叶片均表现正常, 说明突变体spl的斑点叶性状由隐性位点控制。而这3个杂种F1的自交F2后代的田间结果表明, 具正常叶片表型的单株与具斑点叶表型的单株数之比均符合孟德尔3∶1的分离比(表2), 证实突变体spl的斑点叶性状受一对隐性核基因控制。
1: 剑叶; 2: 倒二叶; 3: 倒三叶。**在0.01水平上差异显著。 C: NBT染色。D: DAB染色。
1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top.**Significantly different at<0.01 (-test). C: NBT staining. D: DAB staining.
利用spl/02428的F2群体中的207株具斑点叶性状的单株作为定位群体。选取均匀分布于水稻12条染色体上的500对SSR标记及50对InDel标记逐条对02428和突变体spl进行亲本多态性分析,利用筛选到的多态性标记分析正常基因池和突变基因池。发现在突变体基因池和正常基因池水稻第12染色体上的SSR标记RM511、RM1246和RM1300 (引物序列见附表1)存在明显偏分离, 利用207个突变体单株验证以上标记, 初步把SPL定位在RM511和RM1300之间(图7)。为进一步定位SPL基因, 利用RM511和RM1300之间的5对在两亲本之间具有多态性的SSR标记(包括RM28204、RM1246、RM28466、RM28485和RM28502)(引物序列见附表1)将该基因定位在RM28466与RM28485之间, 物理距离约189 kb, 横跨AL935072、AL928753、BX072546和AL713908等4个BAC (图7), 其间有EST支持的22个ORF (http://rapdb.dna. affrc.go.jp/viewer/gbrowse/irgsp1/)(表3)。
3 讨论
斑点叶突变体最明显的外在表现是叶片上出现坏死斑点或斑块, 而内在表现则为程序性细胞死亡(programmed cell death, PCD), 并由此带来叶绿体降解、蛋白质降解和ROS (reactive oxygen species, ROS)累积等众多生理生化变化[7-8]。研究表明, 叶绿体降解不仅引起ROS累积, 也介导PCD形成[7], 因此叶绿素含量及叶绿素的比值是衡量细胞死亡的重要生理指标。利用EMS诱变获得斑点叶突变体spl, 其斑点叶症状始于分蘖期(图1-A), 孕穗期后除剑叶外所有叶片均不同程度出现褐色斑点(图1-B, C, D, E, F)。叶绿素含量测定表明, 突变体剑叶、倒二叶和倒三叶的叶绿素/比值均低于其野生型珍97且依次显著下降, 说明突变体叶肉细胞已发生PCD, 而且其叶绿素比叶绿素下降更快, 究其原因可能是倒二叶和倒三叶中的ROS累积, 特别是O2܋的累积显著高于剑叶(图5-A), 导致叶绿素对活性氧离子的敏感性高于叶绿素[31]。至于孕穗期突变体光合色素含量明显低于野生型的原因, 可能是突变体基因的突变影响叶绿素的合成[32], 从而造成突变体叶色偏黄(图1-B, C)。
1: 剑叶; 2: 倒二叶; 3: 倒三叶。**在0.01水平上差异显著。
1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. ** Significantly different at<0.01 (-test).
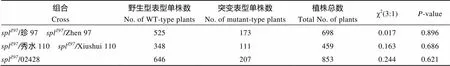
表2 突变体splZ97的遗传分析
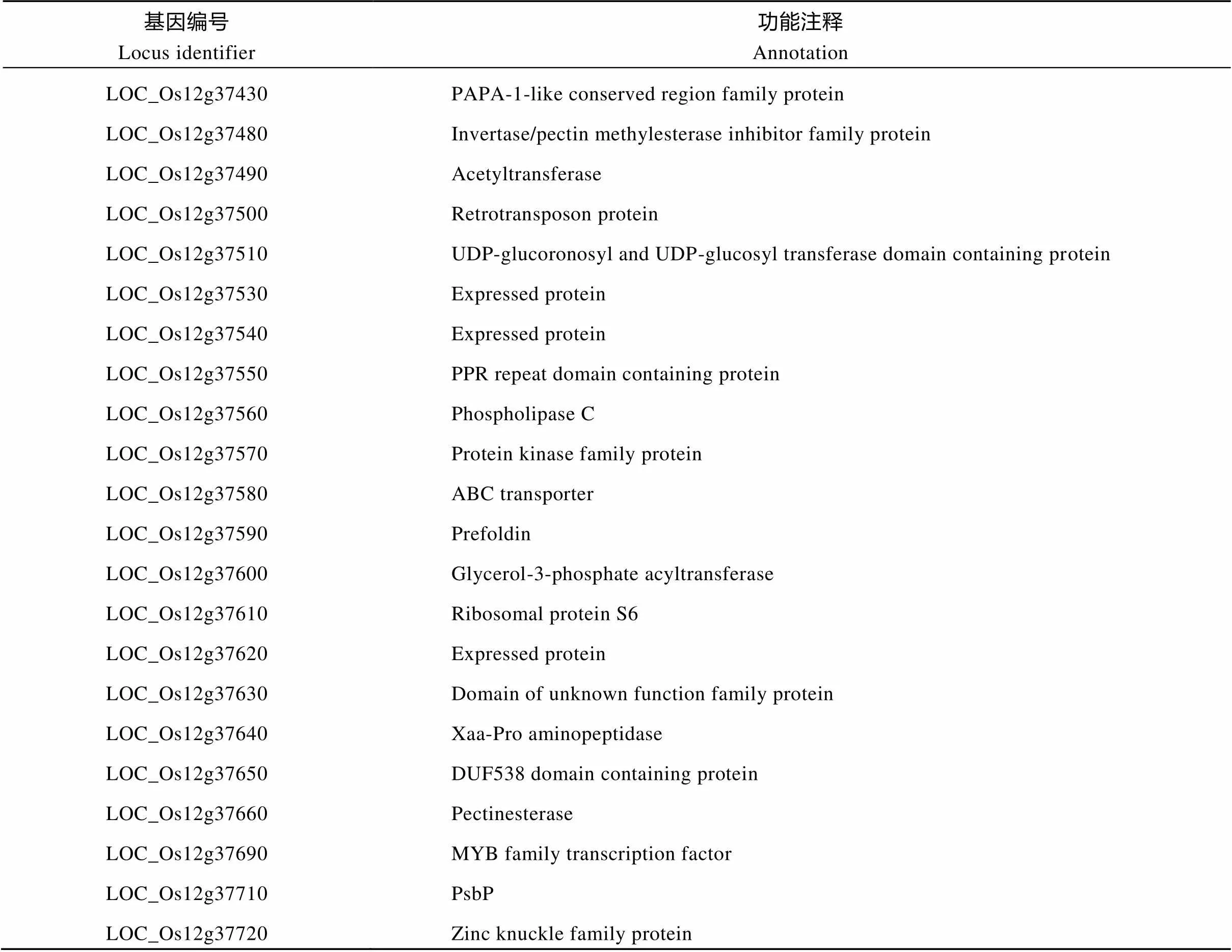
表3 定位区间内的基因及功能注释
引起叶片斑点叶形成的原因极其复杂, 它是由植物自身的生理生化原因及外界环境因素共同作用决定的。其中, 作为植物体内重要信号分子ROS, 在抵御各种逆境胁迫、调控植物生长发育中起重要作用, 其重要来源是叶绿体, 伴随叶片叶绿体的降解或解体, 叶绿体中的电子传递链受到抑制, 致使1O2、H2O2和O2܋大量形成[33]。研究显示, O2܋是引起拟南芥突变体类病斑形成的重要因素, 外源SOD能有效抑制该突变体叶片出现类病斑症状[34]。本研究中野生型珍97的剑叶、倒二叶和倒三叶间的ROS(主要包括H2O2和O2܋)无显著性差异, 但突变体的O2܋含量则极显著高于野生型且依次增加(图5-A), 叶片NBT组织化学染色进一步证实突变体的O2܋明显累积(图5-C)。因此推测O2܋是引起spl斑点叶形成的重要因素。同时, 由于叶片中大量累积ROS, 导致细胞膜脂质过氧化而产生大量MDA (图5-A), 加剧膜损伤并致使细胞坏死[35]。当然, 在正常植株体内, 存在重要的抗氧化酶系统, 如SOD、POD和CAT, 调控ROS的产生和降解, 维持ROS含量处于一个动态平衡而使细胞免于氧化胁迫伤害[36]。其中, CAT专一作用于H2O2, 将其分解成无毒的H2O和O2。本研究中突变体倒二叶和倒三叶的CAT活极显著低于剑叶, 致使倒三叶细胞中存在H2O2累积(图4-B)。至于突变体叶片, 尤其是倒二叶和倒三叶的POD活性(图4-A)和SOD活性(图4-C)极显著高于野生型且依次显著升高的原因, 可能是斑点叶形成初期细胞自我保护, 诱导SOD和POD合成相关基因的表达而使SOD和POD活性增加, 该结果进一步证实了汪媛[37]和赵晨晨等[38]的研究。
此外, 任何与叶片生理生化代谢相关基因的突变, 均有可能破坏细胞内部的生理生化平衡, 导致ROS累积及内源激素的失衡, 引起细胞程序性死亡并最终造成斑点叶的形成。研究表明,、和是植物中最重要的三大转录因子, 调控各种分子生理生化过程, 进而影响植物生长发育[39-42]。过量表达基因, 将导致转基因拟南芥出现类病斑症状, 究其原因, 可能是该基因的过量表达促进了SA合成途径中基因的上调表达, 进而导致SA的累积而造成斑点叶的形成[43-44]。而编码水稻热激转录因子的基因的突变将导致突变体从水稻分蘖期到抽穗期, 整个叶片表面出现红棕色小斑点[9]。此外, 在高粱中表达水稻蛋白激酶基因, 将导致转基因后代表现类病斑症状且对盐敏感[45]。迄今为止, 在第12染色体长臂上已定位或克隆[15]和[46]等斑点叶相关基因, 而这些基因均不在本研究的定位区间, 因此SPL是一个新的斑点叶相关基因。与报道相关的导致植物类病斑形成的基因则仅有蛋白激酶基因(LOC_ Os12g37570)和MYB转录因子(LOC_Os12g37690)。基于突变体的MDA含量、CAT活性、O2–含量及H2O2含量等结果初步认为spl基因与ROS密切相关, 而依据突变体对盐敏感的结果则进一步将候选基因指向蛋白激酶基因(LOC_Os12g37570)。当然,SPL基因的最终确定要依赖于候选基因的测序分析及遗传互补验证。
4 结论
spl是一个新的斑点叶且对盐敏感的突变体, 其斑点叶症状始于分蘖期, 此后由叶边缘向叶片内部扩散, 直至遍布整个叶片。与野生型对照相比, 突变体剑叶、倒二叶和倒三叶的叶绿素含量极显著降低, ROS明显累积, MDA含量、SOD和POD活性极显著升高; 而CAT活性显著下降, 清除H2O2的能力也显著下降。spl斑点叶性状受1对隐性核基因控制, 位于第12染色体长臂的RM28466和RM28485两个标记的189 kb区间, 本研究结果为进一步克隆SPL基因并揭示斑点叶形成的分子生理机制奠定了基础。
References
[1] 孙惠敏, 张春娇, 李保同, 潘晓华. 水稻类病斑突变体的研究进展. 上海农业学报, 2014, 30: 142–147 Sun H M, Zhang C Q, Li B T, Pan X H. Advances of study on rice lesion mimic mutants., 2014, 30: 142–147 (in Chinese with English abstract)
[2] Moeder W, Yoshioka K. Lesion mimic mutants: a classical, yet still fundamental approach to study programmed cell death., 2008, 3: 764–767
[3] Wu C, Bordeos A, Madamba M R, Baraoidan M, Ramos M, Wang G L, Leach J E, Leung H. Rice lesion mimic mutants with enhanced resistance to diseases., 2008, 279: 605–619
[4] Sun C, Liu L, Tang J, Lin A, Zhang F, Fang J, Zhang G, Chu C., encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice., 2011, 38: 29–37
[5] Zhang H,Cao Y,Zhao J,Li X,Xiao J,Wang S. A pair of orthologs of a leucine-rich repeat receptor kinase-like disease resistance gene family regulates rice response to raised temperature., 2011, 11: 160
[6] Yang D L, Yang Y, He Z. Roles of plant hormones and their interplay in rice immunity., 2013, 6: 675–685
[7] Bruggeman Q, Raynaud C, Benhamed M, Delarue M. To die or not to die? Lessons from lesion mimic mutants., 2015, 6: 24
[8] Lorrain S, Vailleau F, Balagué C, Roby D. Lesion mimic mutants: keys for deciphering cell death and defense pathways in plants., 2003, 8: 263–271.
[9] Yamanouchi U, Yano M, Lin H, Ashikari M, Yamada K. A rice spotted leaf gene,, encodes a heat stress transcription factor protein., 2002, 99: 7530–7535
[10] Wang L, Pei Z, Tian Y, He C. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation., 2005, 18: 375–384
[11] Wang S H, Lim J H, Kim S S, Cho S H, Yoo S C, Koh H J, Sakuraba Y, Paek N C. Mutation of SPOTTED LEAF3 () impairs abscisic acid-responsive signaling and delays leaf senescence in rice., 2015, 66: 7045–7059.
[12] Jiang H, Chen Y, Li M, Xu X, Wu G. Overexpression ofresults in oxidative stress and lesion-mimic cell death in rice seedlings.2011, 53: 375–387
[13] Wang Z, Wang Y, Hong X, Hu D, Liu C, Yang J, Li Y, Huang Y, Feng Y, Gong H, Li Y, Fang G, Tang H, Li Y. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defense responses in rice., 2015, 66: 973–987
[14] Fujiwara T, Maisonneuve S, Isshiki M, Mizutani M, Chen L, Wong H L, Kawasaki T, Shimamoto K. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice., 2010, 285: 11308–11313
[15] Zeng L R, Qu S, Bordeos A, Yang C, Baraoidan M, Yan H, Xie Q, Nahm B H, Leung H, Wang G L. Spotted leaf 11, a negative regulator of plant cell death and defense, encodes a U-box/ armadillo repeat protein endowed with E3 ubiquitin ligase activity., 2004, 16: 2795–2808
[16] Mori M, Tomita C, Sugimoto K, Hasegawa M, Hayashi N, Dubouzet J G, Ochiai H, Sekimoto H, Hirochika H, Kikuchi S. Isolation and molecular characterization of a spotted leaf 18 mutant by modified activation-tagging in rice., 2007, 63: 847–860
[17] Jiao B B, Wang J J, Zhu X D, Zeng L J, Li Q, He Z H. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice., 2012, 5: 205–217
[18] Chen X, Hao L, Pan J, Zheng X, Jiang G, Jin Y, Gu Z, Qian Q, Zhai W, Ma B., a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice., 2012, 30: 939–949
[19] Tamiru M, Takagi H, Abe A, Yokota T, Kanzaki H, Okamoto H, Saitoh H, Takahashi H, Fujisaki K, Oikawa K, Uemura A, Natsume S, Jikumaru Y, Matsuura H, Umemura K, Terry M J, Terauchi R. A chloroplast-localized protein LESION AND LAMINA BENDING affects defense and growth responses in rice., 2016, 210:1282–1297
[20] Yang X, Gong P, Li K, Huang F, Cheng F, Pan G. A single cytosine deletion in thegene encoding vacuolar-type H+-ATPase subunit A1 leads to premature leaf senescence and seed dormancy in rice., 2016, 67: 2761–2776
[21] Qiao Y, Jiang W, Lee J, Park B, Choi M S, Piao R, Woo M O, Roh J H, Han L, Paek N C, Seo H S, Koh H J.encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice ()., 2010, 185: 258–274
[22] Fekih R, Tamiru M, Kanzaki H, Abe A, Yoshida K, Kanzaki E, Saitoh H, Takagi H, Natsume S, Undan J R, Undan J, Terauchi R. The rice (L.) LESION MIMIC RESEMBLING, which encodes an AAA-type ATPase, is implicated in defense response., 2015, 290: 611–622
[23] Manosalva P M, Bruce M, Leach J E. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance., 2011, 68: 777–787
[24] Undan J R, Tamiru M, Abe A, Yoshida K, Kosugi S, Takagi H, Yoshida K, Kanzaki H, Saitoh H, Fekih R, Sharma S, Undan J, Yano M, Terauchi R. Mutation in, a gene encoding a protein with two double-stranded RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (L.)., 2012, 87: 169–179
[25] Murashige T, Skoog F. A revised medium for rapid growth and bio assays with tobacco tissue cultures., 1962, 15: 473–497
[26] 张治安, 陈展宇. 植物生理学实验技术. 长春: 吉林大学出版社, 2008. p 7 Zhang Z A, Chen Z Y. Experiment Technology of Plant Physiology. Changchun: Jilin University Press, 2008. p 7 (in Chinese)
[27] Kariola T, Brader G, Li J, Palva E T. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants., 2005, 17: 282–294
[28] Mahalingam R, Jambunathan N, Gunjan S K, Faustin E, Weng H, Ayoubi P. Analysis of oxidative signalling induced by ozone in., 2006, 29: 1357–1371
[29] Shen Y J, Jiang H, Jin J P, Zhang Z B, Xi B, He Y Y, Wang G, Wang C, Qian L L, Li X, Yu Q B, Liu H J, Chen D H, Gao J H, Huang H, Shi T L, Yang Z N. Development of genome-wide DNA polymorphism database for map-based cloning of rice genes., 2004, 135: 1198–1205
[30] Panaud O, Chen X, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSR) in rice (L.)., 1996, 252: 597–607
[31] 伍泽堂. 超氧自由基与叶片衰老时叶绿素破坏的关系. 植物生理学通讯, 1991, 27: 277–279 Wu Z T. Relationship between superoxide radical and destruction of chlorophyll during leaf senescence., 1991, 27: 277–279 (in Chinese with English abstract)
[32] Wang F, Wu W, Wang D, Yang W, Sun J, Liu D, Zhang A. Characterization and genetic analysis of a novel light-dependent lesion mimic mutant,, showing adult-plant resistance to powdery mildew in common wheat., 2016, 11: e0155358
[33] Hideg E, Kalai T, Kos P B, Asada K, Hideg K. Singlet oxygen in plants: its significance and possible detection with double (fluorescent and spin) indicator reagents., 2006, 82: 1211–1218
[34] Dietrich R A, Richberg M H, Schmidt R, Dean C, Dangl J L. A novel zinc finger protein is encoded by thegene and functions as a negative regulator of plant cell death., 1997, 88: 685–694
[35] 华春, 王仁雷. 杂交稻及其三系叶片衰老过程中SOD、CAT活性和MDA含量的变化. 西北植物学报, 2003, 23: 406–409 Hua C, Wang R L. Changes of SOD and CAT activities and MDA content during senescence of hybrid rice and three lines leaves., 2003, 23: 406–409 (in Chinese with English abstract)
[36] You J, Chan Z. ROS regulation during abiotic stress responses in crop plants., 2015, 6: 1092
[37] 汪媛. 水稻叶片衰老过程生理变化及蛋白质降解与蛋白酶活性变化研究. 扬州大学硕士学位论文, 江苏扬州, 2010Wang Y. The Research of Physiological Changes, Protein Degradation and Protease Activity in the Process of Leaf Senescence in Rice. MS Thesis of Yangzhou University, Yangzhou, China, 2010 (in Chinese with English abstract)
[38] 赵晨晨, 黄福灯, 龚盼, 杨茜, 程方民, 潘刚. 水稻叶片早衰突变体的生理特征与基因定位. 作物学报, 2014, 40: 1946–1955 Zhao C C, Huang F D, Gong P, Yang X, Cheng F M, Pan G. Physiological characteristics and gene mapping of a leaf early-senescence mutantin rice., 2014, 40: 1946–1955 (in Chinese with English abstract)
[39] Lindemose S, O’Shea C, Jensen M K, Skriver K. Structure, function and networks of transcription factors involved in abiotic stress response., 2013, 14: 5842–5878
[40] Shao H, Wang H, Tang X.transcription factors in plant multiple abiotic stress responses: progress and prospects., 2015, 6: 902
[41] Ambawat S, Sharma P, Yadav N R, Yadav R C.transcription factor genes as regulators for plant responses: an overview., 2013, 19: 307–321
[42] Bakshi M, Oelmüller R.transcription factors: Jack of many trades in plants., 2014, 9: e27700
[43] Raffaele S, Rivas S, Roby D. An essential role for salicylic acid in-mediated control of the hypersensitive cell death program in., 2006, 580: 3498–3504
[44] Vailleau F, Daniel X, Tronchet M, Montillet J L, Triantaphylidès C, Roby D. Agene,, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack.2002, 99: 10179–10184
[45] Mall T K,Dweikat I,Sato S J,Neresian N,Xu K,Ge Z,Wang D,Elthon T,Clemente T. Expression of the ricein sorghum: molecular and phenotypic analyses., 2011, 75: 467–479
[46] 宋莉欣, 黄奇娜, 奉保华, 施勇烽, 张晓波, 徐霞, 王惠梅, 李小红, 赵宝华, 吴建利. 水稻斑点叶突变体的鉴定与基因定位. 作物学报, 2015, 41: 1519–1528 Song L X, Huang Q N, Feng B H, Shi Y F, Zhang X B, Xu X, Wang H M, Li X H, Zhao B H, Wu J L. Characterization and gene mapping of a spotted-leaf mutantin rice (L.)., 2015, 41: 1519–1528 (in Chinese with English abstract)
附表1 用于SPL基因定位的SSR标记
Supplementary table 1 SSR markers used forSPLgene mapping
Physiological Characters and Gene Mapping of a Spotted-leaf Mutantsplin Rice
WEI Li-Quan1,**, LUO Yan-Min1,**, WANG Wen-Qiang1, CHI Chang-Cheng1, HUANG Fu-Deng2, XIANG Xun1, CHENG Fang-Min1, and PAN Gang1,*
1College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058, China;2Institute of Crop and Nuclear Technology Utilization, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
A spotted-leaf mutantsplwas isolated from a mutant bank generated by EMS mutagenesis ofrestore line Zhen 97. Under field conditions, the brown lesion-mimics mutant splfirstly displayed at the tip and edge of leaf blade at tillering stage, and then gradually spread to whole leaf, resulting in the death of the whole blade when the symptom was severe. At the same time, the major agronomic traits including plant height, grain number per panicle and seed-setting rate were markedly affected. Compared with the wild-type the flag leaf, the second leaf from top and the third leaf from top at heading stage, chlorophyll contents in the mutantsplsignificantly decreased, while POD (peroxidase, POD) activity, O2܋level and MDA (malondialdehyde, MDA) content increased. In addition, CAT (catalase, CAT) activity and soluble protein content of the second leaf from top and the third leaf from top of the mutant decreased as compared with the wild type; on the contrary, the SOD (superoxide dismutase, SOD) activity significantly increased. The histochemical analysis further indicated thatO2܋accumulated in the leaf blade of the mutantspl. In addition, under salt stress at seedling stage, the shoot length and root length of the mutant splwere significantly shorter than these of the wild type. Genetic analysis and gene mapping showed thatsplwas controlled by a single recessive nuclear gene, which was mapped to a region of 189 kb flanked by two SSR markers RM28466 and RM28485 on the long arm of chromosome 12. These results achieved in the present study would further facilitate the cloning and functional analysis of the geneSPL.
Rice;spl; Spotted-leaf; Physiological characters; Gene mapping
10.3724/SP.J.1006.2017.00648
本研究由国家自然科学基金项目(31271691)和国家转基因生物新品种培育重大专项(2016ZX08001-002)资助。
This study was supported by the Natural Science Foundation of China (31271691) and the National Major Project for Developing New GM Crops (2016ZX08001-002)
(Corresponding author): 潘刚, E-mail: pangang12@126.com**同等贡献(Contributed equally to this work)
(收稿日期): 2016-08-13; Accepted(接受日期): 2017-01-21; Published online(网络出版日期):2017-02-17.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20170217.1001.012.html
