游离态氨基酸对羊角月牙藻的生物有效性
2017-05-22翟天恩霍守亮张靖天席北斗肖喆
翟天恩,霍守亮,张靖天,席北斗,肖喆
中国环境科学研究院地下水与环境系统创新基地,北京 100012
游离态氨基酸对羊角月牙藻的生物有效性
翟天恩,霍守亮*,张靖天,席北斗,肖喆
中国环境科学研究院地下水与环境系统创新基地,北京 100012
采用实验室模拟培养试验,以6种个体氨基酸——丝氨酸、天冬氨酸、谷氨酸、甘氨酸、丙氨酸和精氨酸为唯一氮源,采用一次性培养的方法,在无菌条件下,研究羊角月牙藻对不同种类个体氨基酸和不同浓度混合氨基酸的利用潜能。结果表明:藻细胞在生长过程中会释放氨基酸增加水体中氨基酸浓度,同时,藻类可迅速并直接利用多种游离态氨基酸快速生长。氨基酸浓度在培养周期的前2天减少90%以上,藻细胞数在第6~8天达到最大,说明藻细胞的生长滞后于游离氨基酸浓度的下降;在个体氨基酸试验中,藻细胞对氨基酸的利用程度为精氨酸>丙氨酸>谷氨酸>天门冬氨酸>丝氨酸>甘氨酸;在混合氨基酸试验中,藻细胞的比增殖速率明显高于个体氨基酸试验组,且随混合氨基酸浓度的增加,最大藻细胞生物量增加;羊角月牙藻在利用个体氨基酸的同时也会释放其他种类的氨基酸,其中,天冬氨酸和甘氨酸的释放浓度最大。羊角月牙藻在无机氮缺乏情况下,能迅速利用氨基酸。
游离态氨基酸;溶解性有机氮;羊角月牙藻;生物有效性
氮是水生态系统中重要的营养元素之一,参与浮游生物的新陈代谢活动,对浮游生物生长具有重要的作用。研究表明,细菌和藻类可直接利用无机氮,并且细菌可将一些有机氮转化为无机氮形式作为碳源和氮源供给生命体的新陈代谢活动[1]。近期研究发现,一些浮游植物也可以直接利用有机氮[1-3],并且在溶解性无机氮(DIN)浓度较低时,高浓度的溶解性有机氮(DON)会促进浮游植物的增殖[1,3]。氨基酸作为DON库中活跃的组分,广泛存在于自然水体、沉积物、大气中,包括游离态氨基酸(DFAA)和结合态氨基酸(DCAA)。湖泊水体中28%~46%的DON为氨基酸类有机氮[4]。近海表层沉积物氨基酸组成中[5],含量较高的氨基酸有天冬氨酸、谷氨酸、丙氨酸和赖氨酸,其平均摩尔分数均在80%以上。大气PM2.5样品中也检测到DFAA,主要以鸟氨酸、苏氨酸、甘氨酸、丝氨酸、甲胺和丙氨酸为主,占DFAA总量的81.1%[6]。在北太平洋西部气溶胶中发现的氨基酸主要以甘氨酸、丝氨酸、天冬氨酸和精氨酸为主,占DFAA总量的72%[7]。污水处理厂出水中也发现15%~35%的有机氮为氨基酸类物质[8],主要包括谷氨酸、丝氨酸、天冬氨酸和甘氨酸,以及少量的鸟氨酸、组氨酸、赖氨酸和半膀氨酸。
国内外学者对DON的生物有效性已开展多项研究:如Pehlivanoglu等[9]研究发现,在有菌条件下56%的DON可以被藻类所利用,同时发现接近30%的DON是小分子类含氮化合物(<1 kDa),但在无菌条件下,DON几乎不能被藻类所利用;Urgun-Demirtas等[10]研究发现,在有菌和无菌条件下,有18%~61%的DON能被藻类所利用;Liu等[11-12]将DON分为亲水、疏水组分,发现亲水组分可刺激藻类生长;Su等[13]进一步研究了细菌对DON不同极性组分的利用特性,发现细菌能够利用亲水、疏水组分中的氨基酸,并且在后期可通过水解作用将溶解性蛋白质物质分解为缩氨酸和自由态氨基酸,导致氨基酸浓度缓慢增长。
DFAA是重要的小分子有机氮,具有很强的活性,不仅能作为细菌生长的碳源和氮源,而且在DIN浓度较低时,可被浮游植物通过直接吸收或胞外酶分解间接利用[14]。依据氨基酸分子中所含氨基和羧基数目的不同,可将DFAA分为酸性氨基酸(天冬氨酸、谷氨酸)、碱性氨基酸(组氨酸、精氨酸)和中性氨基酸。目前,对藻类吸收利用氨基酸已开展了研究。徐宁等[15]对比研究了不同条件下4种海洋微藻对DFAA(试验浓度为100 μmolL)的利用特性,结果表明,海洋微藻对游离态氨基酸的利用存在显著的种间差异,东海原甲藻和球形棕囊藻可在无菌条件下以氨基酸为唯一氮源快速生长,而米氏凯伦藻和中肋骨条藻必须依赖细菌才能利用氨基酸维持生长。Zhang等[2]研究蛋白核小球藻(Chlorellapyrenoidosa)对18种个体氨基酸在高浓度(0.05和0.5 gL)条件下的利用潜能发现,有10种氨基酸均能促进小球藻的生长,特别是天冬氨酸、精氨酸和甘氨酸更能促进藻细胞的生长。John等[16]研究发现,丰迪亚历山大藻(Alexandriumfundyense)在低浓度(50 μmolL)条件下可以利用多种不同个体氨基酸并能在数小时内将氨基酸浓度降低50%左右。Dai等[17]研究发现,在无菌条件下,铜绿微囊藻(Microcystisaeruginosa)对100 μmolL浓度下不同种类氨基酸的利用程度存在明显差异,精氨酸、丙氨酸和亮氨酸可被藻细胞迅速利用并促进细胞增殖,而谷氨酸、天冬氨酸和赖氨酸尽管可以通过细胞膜进入胞内,但不能促进藻细胞的生长。Yan等[18]运用15N同位素示踪技术研究发现,铜绿微囊藻在精氨酸中的生长状况优于氨氮。
羊角月牙藻(Selenastrumcapricornutum)作为自然水体中常见的藻类,因其细胞特征明显,易于培养,且试验结果重现性较好,已广泛应用于DON的生物有效性研究上[9-12]。近年来以羊角月牙藻为测试藻种,从DON不同分子量分布以及亲疏水极性开展大量研究,但多从菌藻共存条件下出发,且对于DON中的已知组分(如氨基酸、尿素等)的生物有效性报道较少。同时,关于藻类利用游离态氨基酸的研究侧重于个体氨基酸作为氮源,所研究的氨基酸浓度大多大于50 μmolL,高于自然水体中的浓度,多为海洋微藻[15,19-20]或有毒藻种[17-18],不同藻种的生物利用特性存在一定差异,因而有必要展开羊角月牙藻对氨基酸的生物有效性研究。笔者拟选择环境中存在的6种典型个体氨基酸作为氮源,包括酸性、碱性和中性氨基酸,以淡水藻种羊角月牙藻作为试验藻种,研究在低浓度环境和无菌条件下,藻类对游离态氨基酸的利用特性,以期为藻类对DON的生物有效性研究提供理论依据。
1 材料与方法
1.1 试验藻种及培养
试验藻种为羊角月牙藻,由中国科学院水生生物研究所提供。试验前,选用BG11培养基,在藻细胞处于对数生长期反复接种3~4次,使细胞达到同步增长。接种前,培养基通过 121 ℃高温灭菌15 min后,移至超净台中操作。所有容器及试验耗材均经过高温灭菌后使用。藻细胞生长温度为(25±1)℃,光照强度为2 000~3 000 lx,光照周期为12 h∶12 h,在恒温光照培养箱中进行培养试验。
无氮培养:将处于对数生长期的藻细胞离心,用超纯水清洗2~3次,接种至无氮BG11培养基(即培养基中不添加NaNO3组分)中培养,每天按时测定藻细胞生物量,培养1周左右藻细胞生物量处于稳定状态,认为此时藻细胞体内贮存的氮消耗殆尽。
1.2 试验设计
1.2.1 个体氨基酸的生物有效性
选择丝氨酸(Ser)、天冬氨酸(Asp)、谷氨酸(Glu)、甘氨酸(Gly)、丙氨酸(Ala)和精氨酸(Arg)为研究对象,以无氮培养液做空白对照组,在实验室条件下开展藻类培养试验,评估个体氨基酸对羊角月牙藻的生物有效性,即培养过程中羊角月牙藻所能利用的个体氨基酸浓度占初始个体氨基酸浓度的比例。
1.2.2 混合氨基酸的生物有效性
以6种个体氨基酸的混合液作为藻类生长的氮源,以无氮培养液为参照组,评估不同浓度梯度混合氨基酸对藻类的生物有效性,即培养过程中羊角月牙藻所能利用的混合氨基酸浓度占初始混合氨基酸浓度的比例。
采用实验室一次性培养的方法,试验共设置12组,其中包括1组空白组,6组个体氨基酸试验组和5组混合氨基酸试验组,每组2个平行试验,试验周期为15 d,藻细胞初始接种密度为3.75×104cellsmL。试验设计如表1所示。
将上述配置好的不同试验组培养基置于121 ℃高温灭菌锅中灭菌30 min,冷却至室温后移至无菌超净台,接种无氮培养后的藻细胞并取水样作为初始培养基浓度,后期取样均在无菌操作台中进行,光照培养箱在放入试验组前用仪器自带紫外灯灭菌。
1.3 藻细胞生物量测定
每天对藻细胞生物量进行测定,用显微镜(NIKON,日本尼康株式会社)观察藻细胞培养是否出现细菌污染,并记录藻细胞生物量。比生长速率(μ)的计算公式如下:
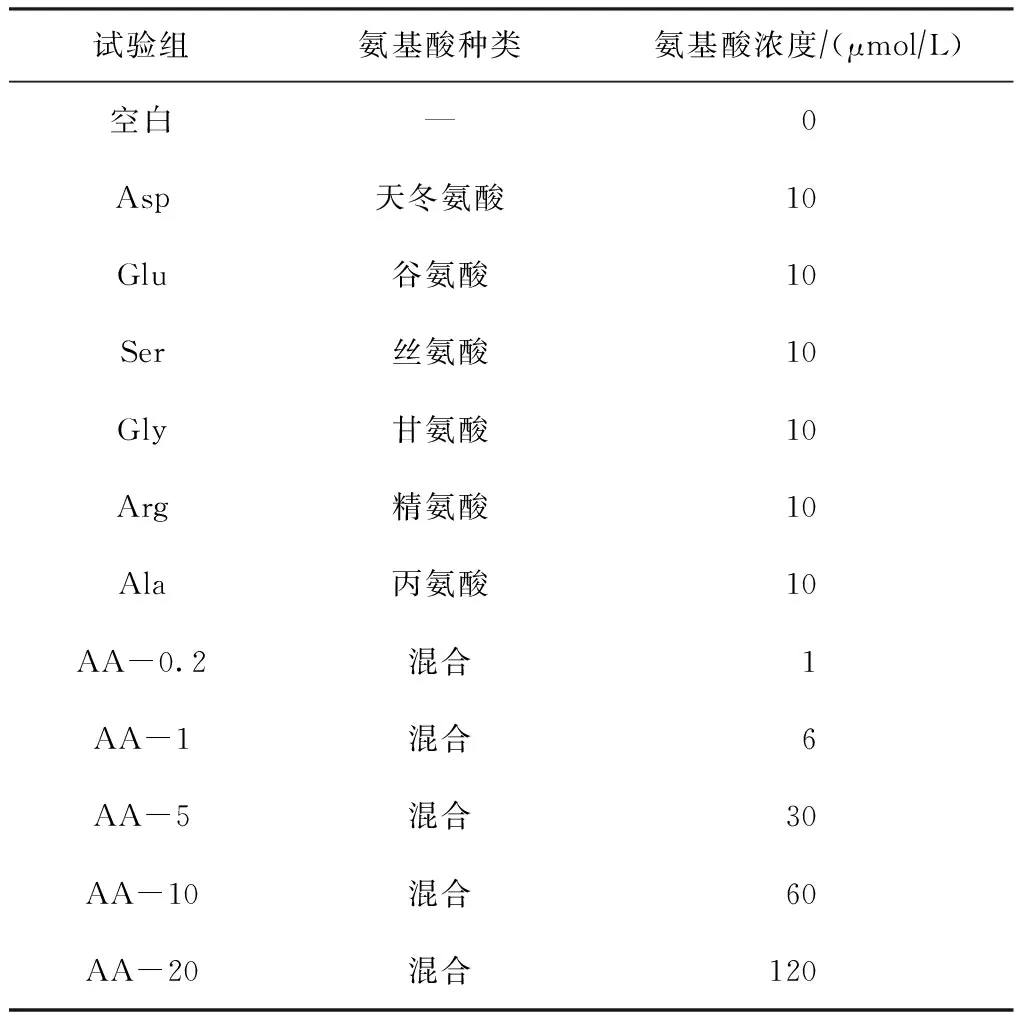
表1 各试验组培养基中的氨基酸种类及浓度
注:混合为天冬氨酸、谷氨酸、丝氨酸、甘氨酸、精氨酸和丙氨酸6种个体氨基酸混合液。
μ=(lnN1-lnN0)(T1-T0)
式中N1和N0分别为第T1天和T0天时的细胞数。
1.4 氨基酸的测定
1.4.1 试验仪器和试剂
试验仪器采用Agilent-1200液相色谱仪,配有在线脱气机、四元梯度泵、标准自动进样器(20 μL)、荧光检测器(fluorescence detector,FLD)及Agilent化学工作站。
氨基酸混合标准溶液包括天冬氨酸、丝氨酸、色氨酸(Trp)、谷氨酸、甘氨酸、组氨酸(His)、精氨酸、苏氨酸(Thr)、丙氨酸、脯氨酸(Pro)、胱氨酸(Cys)、酪氨酸(Tyr)、缬氨酸(Val)、蛋氨酸(Met)、异亮氨酸(Ile)、亮氨酸(Leu)及苯丙氨酸(Phe);个体氨基酸试剂包括天冬氨酸、谷氨酸、丝氨酸、甘氨酸、精氨酸和丙氨酸,试验中所使用的氨基酸以及衍生化试剂邻苯二甲醛(OPA,HPLC)和3-巯基丙酸(3-MPA,HPLC)均为Sigma (St Louis, MO, USA)生产;甲醇、乙腈和四氢呋喃(THF)为HPLC级试剂;试验用水是Mill-Q水(Millipore, USA);其他试剂均为优级纯。
色谱柱为Agilent Eclipse XDB C18柱(5 μm,4.6 mm×250 mm)。
1.4.2 试剂配制
氨基酸混合标准溶液不同浓度梯度配制:准确吸取一定量的氨基酸混合标准溶液,用Mill-Q水配置成0.005、0.010、0.025、0.050、0.100、0.250、0.500、1.000、2.000、5.000和10.000 μmolL不同浓度梯度溶液,冷藏备用。
OPA衍生化试剂配制:将13.4 mg OPA溶解于25 mL甲醇中;取出5 mL加入20 μL 3-巯基丙酸;用pH为9.9±0.05的硼酸钠缓冲溶液定容到25 mL,最终pH为9.3±0.05;该溶液避光陈化90 min以上,以降低试剂空白;保存在4 ℃冰箱中,最长使用9 d。
1.4.3 衍生化反应
将200 μL衍生化试剂添加到0.5 mL氨基酸混合标准溶液或试验样品中反应10 min后,以微量进样器吸取20 μL进样。
1.4.4 色谱条件的选择与设定
根据Confer等[21]研究设置以下试验条件:1)柱温为35 ℃;2)流动相的pH为7.2,流动相中THF的浓度为1.5%;3)将反应时间控制在10~15 min;4)梯度洗脱条件如表2所示;5)荧光扫描样品时间为31 min,并保持液相色谱后运行10 min使压力恢复到正常水平(即只进流动相的压力),检测波长Ex为337 nm,Em为454 nm。
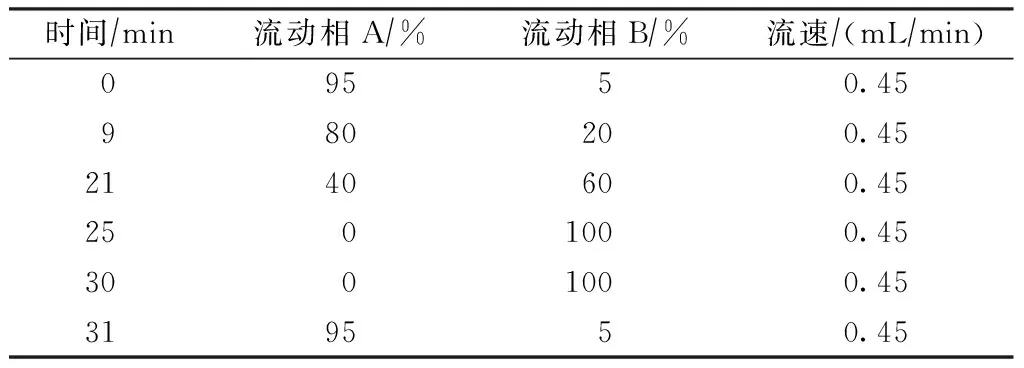
表2 流动相梯度洗脱
2 结果与讨论
2.1 氨基酸标准曲线的建立
利用个体氨基酸标准溶液的保留时间定性,外标法定量,利用不同浓度混合标准溶液建立的各氨基酸标准曲线相关系数均在0.996以上,大部分在0.999以上。
用保留时间法对样品中氨基酸定性,即在相同的色谱分析条件下,样品中氨基酸的色谱峰与氨基酸标准的色谱峰保留时间相同或相近,则认为样品中含有此氨基酸。采用峰面积法对水样中游离态氨基酸的浓度进行定量分析。氨基酸标准的液相色谱如图1所示。该分析方法进行一次氨基酸分析需要30 min,除了半胱氨酸在FLD下无信号,脯氨酸不能与OPA生成衍生物,Phe和Ile分离不太好外,其他氨基酸都得到较为满意的分离。
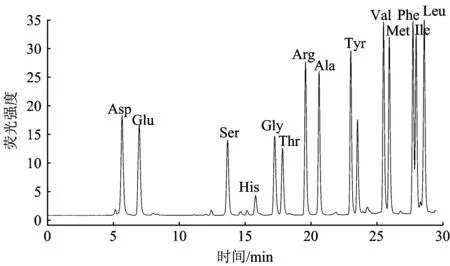
注:每个峰代表1.0 μmolL的氨基酸。图1 氨基酸标准液相色谱Fig.1 Chromatogram of the amino acid standards
在上述最佳试验条件下,所测得各氨基酸标准样品的保留时间、回归方程和相关系数见表3。从表3可以看出,在一定浓度范围内,氨基酸组分峰面积与浓度呈良好的线性关系。
2.2 试验结果
2.2.1 个体氨基酸对藻类的生物有效性
图2为个体氨基酸试验组培养过程中藻细胞生物量的变化和培养中游离态氨基酸浓度的变化。从图2可以看出,以个体氨基酸为唯一氮源时,羊角月牙藻均呈现快速生长态势。在培养的第2天,藻细胞增长速率达到最大,为3×104~8×104cells(mL·d);第2~8天,藻细胞处于稳定期,之后藻细胞生物量出现明显的下降趋势。从最大细胞生物量来看,藻细胞对氨基酸的生物可利用程度为精氨酸>丙氨酸>谷氨酸>天冬氨酸>丝氨酸>甘氨酸。以精氨酸为唯一氮源时,细胞生物量达到最大,为58×104cellsmL。精氨酸属于碱性氨基酸,其分子组成中氨基数目大于羧基数目,1个精氨酸分子中含有4个氮原子,是15种氨基酸中含氮原子数量最多的氨基酸之一,相同浓度的个体氨基酸中,精氨
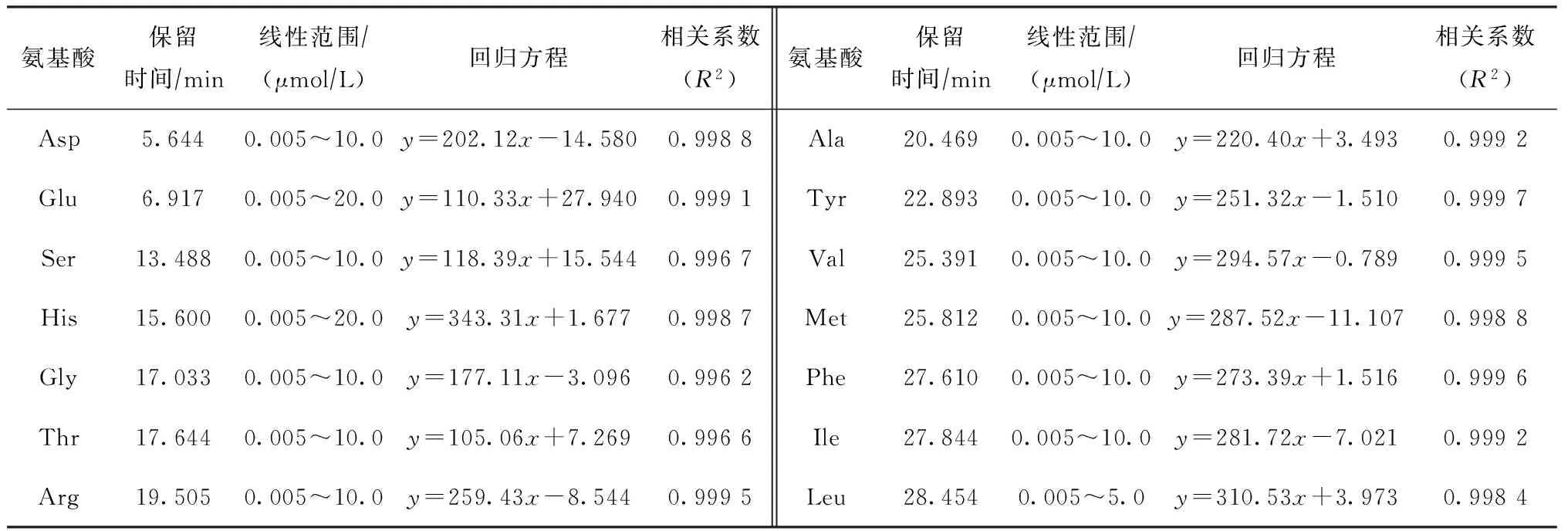
表3 各氨基酸标准样品的保留时间、线性范围、回归方程和相关系数
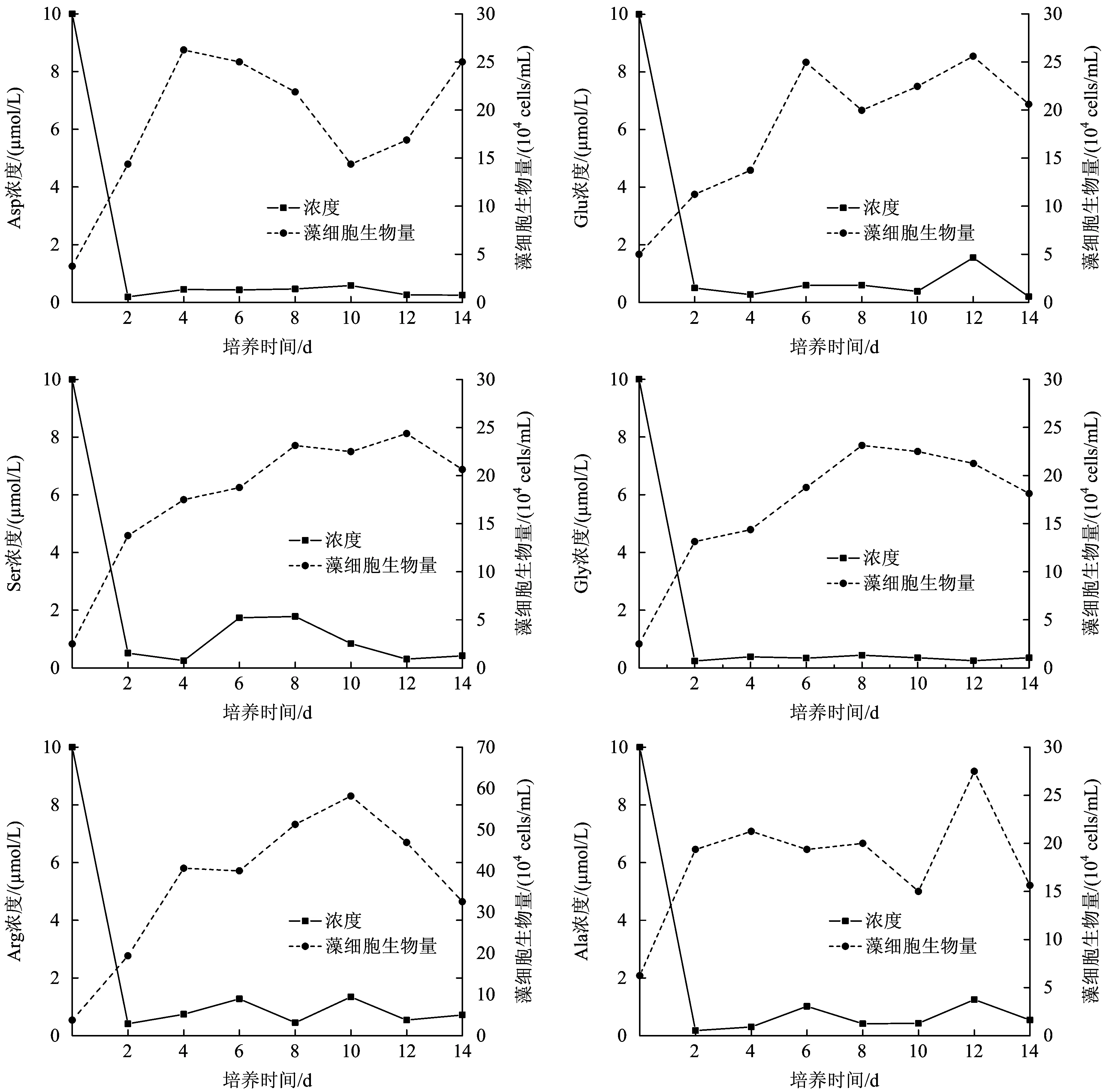
注:氨基酸浓度为培养液中总浓度。图2 个体氨基酸对羊角月牙藻细胞生物量的影响Fig.2 Growth curve of Selenastrum capricornutum and concentration change of amino acid in different individual amino acids situation
酸的氮含量最大,藻细胞生物量增长最多。最大比生长速率可表示藻细胞瞬间最大的生长潜力。以丝氨酸、甘氨酸和精氨酸为唯一氮源时,藻细胞的最大比生长速率分别为0.85、0.82、0.82 d-1;以谷氨酸为唯一氮源时,藻细胞的最大比生长速率为0.41 d-1。谷氨酸属酸性氨基酸,一般认为藻类对中性脂肪族氨基酸利用率较高,而对酸性氨基酸利用率较低,本试验结果进一步印证了该观点。
2.2.2 混合氨基酸对藻类的生物有效性
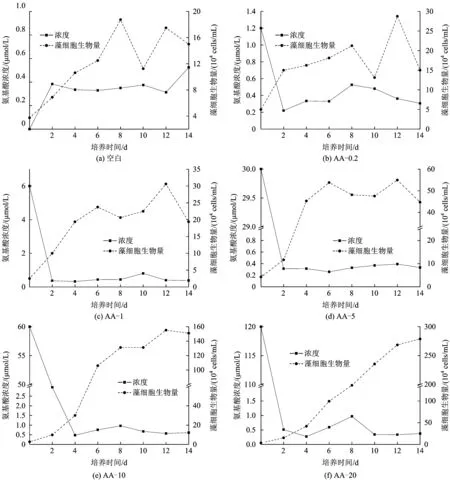
图3 不同浓度混合氨基酸对羊角月牙藻细胞生物量的影响Fig.3 Growth curve of Selenastrum capricornutum and concentration change of total amino acids in different concentrations situation
图3为不同浓度混合氨基酸对藻细胞生物量的影响。从图3可以看出,藻细胞生物量随培养基中氨基酸总浓度的增大而增加。藻细胞最大比生长速率基本出现在前2 d的培养过程中,AA-0.2、AA-1、AA-5、AA-10和AA-20试验组藻细胞最大比生长速率分别为0.55、0.69、0.58、0.65和0.75 d-1。除AA-20试验组外,其他4组试验,藻细胞生物量在第12天均出现不同程度的下降,说明藻细胞进入衰亡期;在AA-20试验组,试验培养的第14天,藻细胞还处于增长过程,说明培养液中还有供给藻细胞增殖的氮源存在。AA-0.2、AA-1、AA-5、AA-10和AA-20试验组的最大藻细胞生物量分别是接种初期的5.75、12.25、15.50、44.00和71.70倍;在个体氨基酸试验中,除精氨酸试验组外,天冬氨酸、谷氨酸、丝氨酸、甘氨酸和丙氨酸5个试验组最大藻细胞生物量分别是接种初期的7.00、5.13、9.75、9.25、15.50和4.40倍:说明混合氨基酸更能促进藻细胞的生长。John等[16]研究了丰迪亚历山大藻在无机氮和氨基酸不同氮源条件下的藻细胞生长情况,结果表明,氨基酸为氮源的环境下最大藻细胞生物量仅达到无机氮环境下的23;而Yan等[18]对铜绿微囊藻在氨氮和丙氨酸不同氮源条件下的研究发现,藻细胞在丙氨酸环境下增加的生物量约是氨氮环境下的3.5倍,其结果与Dai等[17]的基本一致。其差异性的原因首先是藻种类不同[15],其利用氮源的能力存在一定的差异;其次是藻对于不同氮源采用不同的摄入形式,并且对不同氮源的利用程度存在一定的差异[19-20,22]。
2.2.3 藻类生长过程中不同氨基酸变化
在藻类培养过程中,藻细胞会通过自身裂解或死亡产生碳水化合物、多肽、氨基酸等物质[12,17]。表4为羊角月牙藻在无氮培养基内生长过程中水体氨基酸的浓度变化。从表4可以看出,在无外源氨基酸引入的情况下,藻细胞在生长过程中也会释放各种氨基酸。第2天,藻细胞释放了较多的天冬氨酸和甘氨酸,而其他氨基酸的释放量较少。在培养过程中,谷氨酸、组氨酸、苏氨酸、亮氨酸和苯丙氨酸等氨基酸浓度较低,甚至检测不出;天冬氨酸和甘氨酸浓度相对较高,是羊角月牙藻释放的主要氨基酸组成组分。于茜等[23]研究了铜绿微囊藻在培养过程中氨基酸的释放特性,结果表明,藻细胞释放的氨基酸浓度整体呈增加趋势,与本试验结果相似。
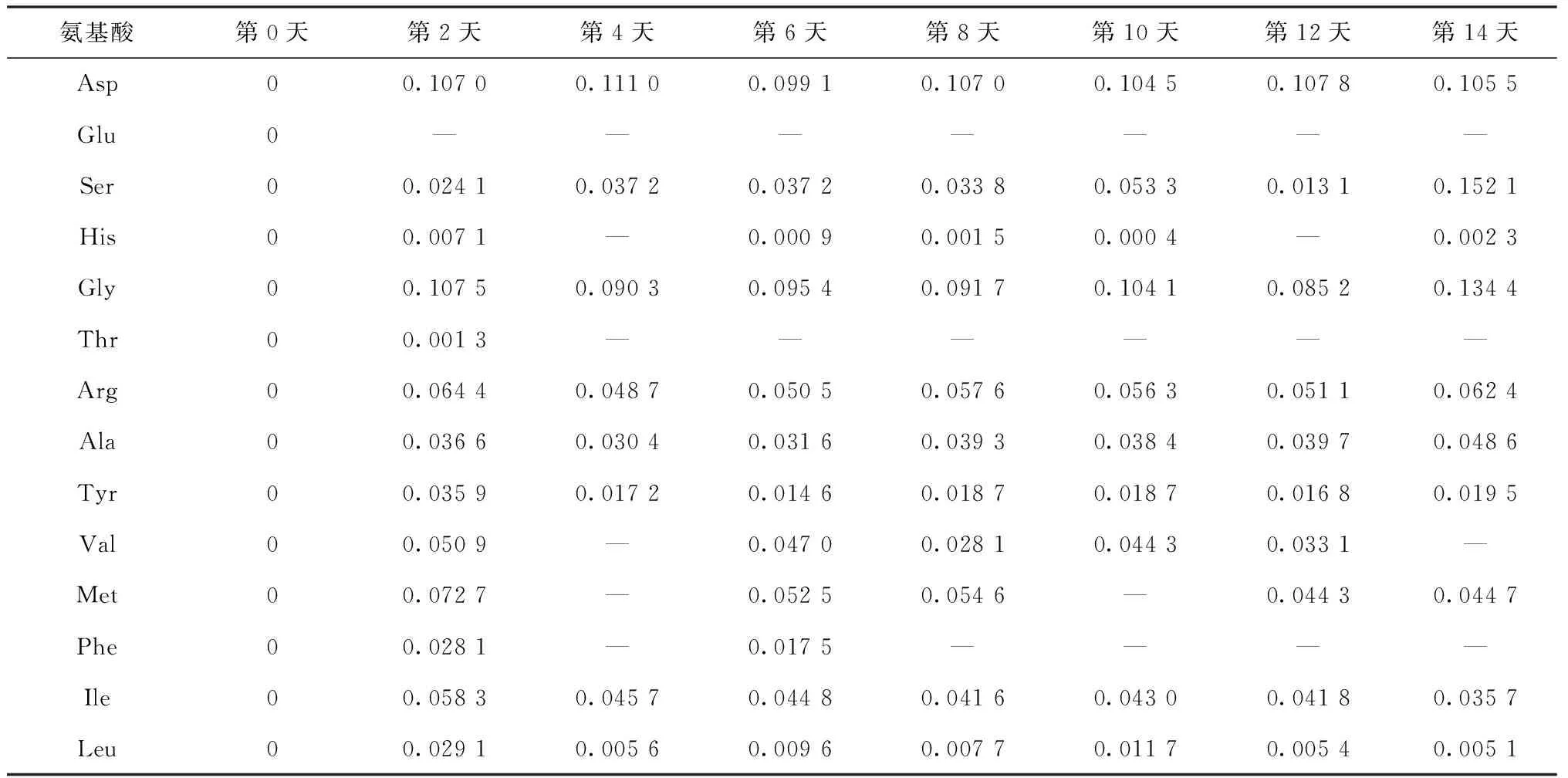
表4 羊角月牙藻无氮生长过程中水体氨基酸浓度变化
注:“—”表示氨基酸浓度低于仪器检测限。
表5给出了不同试验组羊角月牙藻生长过程中氨基酸的浓度变化。个体氨基酸试验组中,第2天因子氨基酸的浓度均发生明显下降,比初始(第0天)下降了99.0%~100%;从第2天开始,因子氨基酸的浓度基本保持不变。混合氨基酸试验组中,第2天因子氨基酸浓度下降了82.5%~99.6%。John等研究[16-18]发现,藻细胞内可能存在储存库和代谢库2个氨基酸库,吸收的氨基酸首先进入储存库,在短时间内可能不会影响代谢库的氨基酸量;随着藻细胞对代谢库中氨基酸的利用,储存库中氨基酸开始向代谢库中转移,这一过程对胞外因子氨基酸的浓度变化影响较小;之后藻细胞通过酶作用将摄取的氨基酸转化为生长所需的能量以及合成新的蛋白质等[24],因而藻细胞增长出现明显的滞后现象。Dai等[17]研究发现,铜绿微囊藻在12 h内可迅速降低培养基中天冬氨酸和谷氨酸的浓度,而对丙氨酸和亮氨酸的吸收速度较慢;培养24 h,天冬氨酸、精氨酸和谷氨酸基本耗尽;同时,不同试验组均检测到其他氨基酸的生成,且浓度变化不大。同样的,本研究发现,不同培养周期内,不同试验组中也检测到除添加的因子氨基酸外的其他氨基酸,且氨基酸种类和浓度与空白试验组基本相同。试验最初添加的藻细胞取自同一瓶藻液,且接种的生物量基本相同,这说明藻细胞释放的氨基酸种类和浓度与因子氨基酸浓度和藻细胞生物量关系不大,可能的原因是氨基酸的释放主要来自培养初期,因而培养过程中氨基酸的释放差别不大。
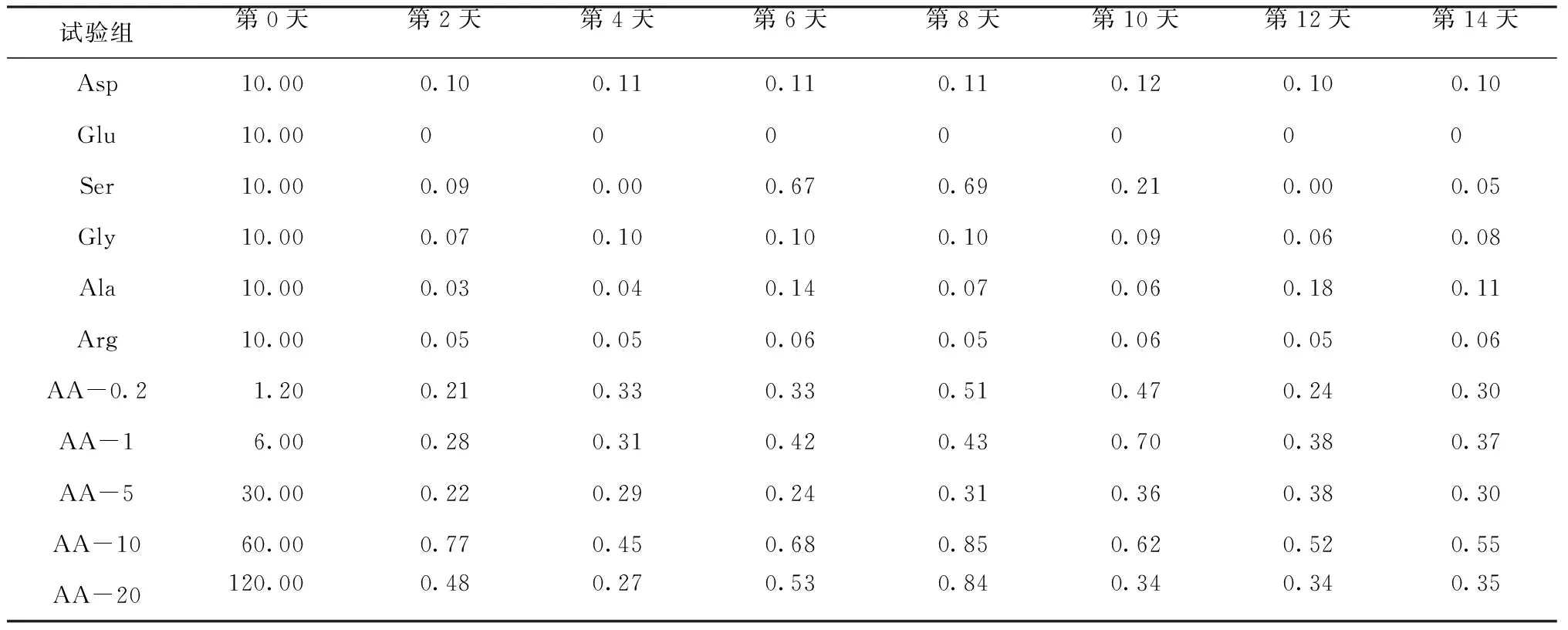
表5 藻类生长过程中水体因子氨基酸浓度变化
3 结论
(1)羊角月牙藻对多种个体氨基酸具有较强的利用能力,氨基酸的利用率达99.0%~100%,藻细胞对氨基酸的生物可利用程度为精氨酸>丙氨酸>谷氨酸>天门冬氨酸>丝氨酸>甘氨酸。
(2)羊角月牙藻在混合氨基酸中的生长状况优于个体氨基酸,且在低浓度(<120 μmolL)条件下,随着氨基酸浓度的增加,其细胞生物量也增加;藻细胞藻培养的前2天,氨基酸利用率达到82.5%~99.6%。
(3)在无菌条件下,藻细胞可直接利用氨基酸作为氮源供给自身生长;在生长过程中,羊角月牙藻会释放其他种类的氨基酸,且主要以天冬氨酸和甘氨酸为主,藻细胞释放氨基酸的种类和数量与添加的氨基酸种类以及生长过程中藻细胞生物量无关。
[1] BRADLEY P B,SANDERSON M P,FRISCHER M E,et al.Inorganic and organic nitrogen uptake by phytoplankton and heterotrophic bacteria in the stratified Mid-Atlantic Bight[J].Estuarine Coastal & Shelf Science,2010,88(4):429-441.
[2] ZHANG W,ZHANG Z,YAN S.Effects of various amino acids as organic nitrogen sources on the growth and biochemical composition ofChlorellapyrenoidosa[J].Bioresource Technology,2015,197:458-464.
[3] LINARES F.Effect of dissolved free amino acids (DFAA) on the biomass and production ofMicrophytobenthiccommunities[J].Journal of Experimental Marine Biology & Ecology,2006,330(2):469-481.
[4] YAO X,ZHU G,CAI L,et al.Geochemical characteristics of amino acids in sediments of Lake Taihu,a large,shallow,eutrophic freshwater lake of China[J].Aquatic Geochemistry,2012,18(3):263-280.
[5] 王丽玲,胡建芳,唐建辉.中国近海表层沉积物中氨基酸组成特征及生物地球化学意义[J].海洋学报,2009,31(6):161-169. WANG L L,HU J F,TANG J H.Amino acid composition and its biogeochemistry implications of surface sediments in coastal areas of China[J].Acta Oceanologica Sinica,2009,31(6):161-169.
[6] ZHANG Q,ANASTASIO C.Free and combined amino compounds in atmospheric fine particles (PM2.5) and fog waters from Northern California[J].Atmospheric Environment,2003,37(16):2247-2258.
[7] MATSUMOTO K,UEMATSU M.Free amino acids in marine aerosols over the Western North Pacific Ocean[J].Atmospheric Environment,2005,39(11):2163-2170.
[8] WESTRHOFF P,MASH H.Dissolved organic nitrogen in drinking water supplies:a review[J].Journal of Water Supply:Research and Technology,2002,51(8):415-448.
[9] PEHLIVANOGLU E,SEDLAK D L.Bioavailability of wastewater-derived organic nitrogen to the algaSelenastrumcapricornutum[J].Water Research,2004,38(14/15):3189-3196.
[10] URGUN-DEMIRTAS M,SATTAYATEWA C,PAGILLA K R.Bioavailability of dissolved organic nitrogen in treated effluents[J].Water Environment Research,2008,80(5):397-406.
[11] LIU H Z,JEONG J,GRAY H,et al.Algal uptake of hydrophobic and hydrophilic dissolved organic nitrogen in effluent from biological nutrient removal municipal wastewater treatment systems[J].Environmental Science and Technology,2012,46(2):713-721.
[12] QIN C,LIU H Z,LIU L,et al.Bioavailability and characterization of dissolved organic nitrogen and dissolved organic phosphorus in wastewater effluents[J].Science of the Total Environment,2015,511:47-53.
[13] SU M,ZHANG J,HUO S,et al.Microbial bioavailability of dissolved organic nitrogen (DON) in the sediments of Lake Shankou,Northeastern China[J].Journal of Environmental Sciences,2016,42:79-88.
[14] MULHOLL M R,GOBLER C J,CINDY L.Peptide hydrolysis,amino acid oxidation and nitrogen uptake in communities seasonally dominated byAureococcusanophagefferens[J].Limnology & Oceanography,2002,47(4):1094-1108.
[15] 徐宁,刘静雅,赖海燕,等.海洋微藻对游离氨基酸的利用特性研究[J].环境科学学报,2013,33(4):1058-1065. XU N,LIU J Y,LAI H Y,et al.Uptake and utilization characteristics ofProrocentrumdonghaiense,Phaeoecystisglobosa,KareniamikimotoiandSkeletonemacostatumfor dissolved free amino acids[J].Acta Scientiae Circumstantiae,2013,33(4):1058-1065.
[16] JOHN E H,FLYNN K J.Amino acid uptake by the toxic dinoflagellateAlexandriumfundyense[J].Marine Biology,1999,133:11-19.
[17] DAI R,LIU H,QU J H,et al.Effects of amino acids on microcystin production of theMicrocystisaeruginosa[J].Journal of Hazardous Materials,2009,161(2/3):730-736.
[18] YAN Y W,DAI R H,LIU Y,et al.Comparative effects of inorganic and organic nitrogen on the growth and microcystin production ofMicrocystisaeruginosa[J].World Journal of Microbiology and Biotechnology,2015,31(5):763-772.
[19] LINDEHOFF E,GRANÉLI E,GLIBERT P M.Nitrogen uptake kinetics ofPrymnesiumparvum(haptophyte)[J].Harmful Algae,2011,12:70-76.
[20] KILLBERG-THORESON L,MULHOLLAND M R,HEIL C A,et al.Nitrogen uptake kinetics in field populations and cultured strains ofKareniabrevis[J].Harmful Algae,2014,38:73-85.
[21] CONFER D R,LOGAN B E,AIKEN B S,et al.Measurement of dissolved free and combined amino acids in unconcentrated wastewaters using high performance liquid chromatography[J].Water Environment Research,1994,67(1):118-125.
[22] AHLUWALIA A S.Effect of amino acids and some chemicals on cellular differentiation inAnabaenaoscillarioides[J].Annals of Biology (India),1985,1:137-144.
[23] 于茜,朱元荣,王焕华,等.铜绿微囊藻培养过程中氨基酸的释放特征及其对水体有机质的贡献[J].环境科学研究,2016,29(3):360-367. YU X,ZHU Y R,WANG H H,et al.Release of amino acids fromMicrocystisaeruginosaand its contributions to organic matter[J].Research of Environmental Sciences,2016,29(3):360-367.
[24] BRONK D A,SEE J H,BRADLEY P, et al.DON as a source of bioavailable nitrogen for phytoplankton[J].Biogeosciences,2007,4(3):283-296.□
Bioavailability of dissolved free amino acids to algaSelenastrumcapricornutum
ZHAI Tianen, HUO Shouliang, ZHANG Jingtian, XI Beidou, XIAO Zhe
Innovation Base of Groundwater and Environmental System Engineering, Chinese Research Academy of Environmental Sciences,Beijing 100012, China
In order to understand the incubation process of algae on the low concentrations of dissolved free amino acids (DFAA), the growth ofSelenastrumcapricornutumwas studied in the laboratory. Six individual amino acids, i.e. serine, aspartic acid, glutamic acid, glycine, alanine and arginine, were used as the sole and mixed nitrogen sources for algal incubation under axenic conditions. The results showed that algae were able to utilize various amino acids for growth; meanwhile, algae could spread different amino acids in the incubation. DFAA concentration decreased with up to 90% in the 2 days, while the cells number of algae reached to the maximum in the 6-8 days, meaning that the growth of algae cell number lags behind the decrease of the of DFAA content. In the individual amino acids experiments, rank order of the use of amino acid was: arginine>alanine>glutamic acid>aspartic acid>serine>glycine. In the mixed amino acids experiments, the cell proliferation rate of algae was significantly higher than that of individual amino acid group; meanwhile with the increase of mixed amino acid concentration, the maximum number of algal cells was also increased.Selenastrumcapricornutumcan also release other kinds of amino acids in the use of the individual amino acid, among them, the main release amino acids are aspartic acid and glycine. Algae could use amino acid rapidly in the lack of inorganic nitrogen condition, which provided a theoretical basis to illustrate dissolved organic nitrogen promotes algae growth.
dissolved free amino acids (DFAA); dissolved organic nitrogen (DON);Selenastrumcapricornutum; bioavailability
2016-09-22
国家水体污染控制与治理科技重大专项(2012ZX07101-002)
翟天恩(1991—),男,硕士研究生,主要研究方向为溶解性有机氮的生物有效性,nyzhai@163.com
*通信作者:霍守亮(1981—),男,研究员,博士,主要从事湖泊营养物基准标准及氮磷迁移转化研究,huoshouliang@126.com
503.23
1674-991X(2017)03-0348-09
10.3969/j.issn.1674-991X.2017.03.049
翟天恩,霍守亮,张靖天,等.游离态氨基酸对羊角月牙藻的生物有效性[J].环境工程技术学报,2017,7(3):348-356.
ZHAI T E, HUO S L, ZHANG J T, et al.Bioavailability of dissolved free amino acids to algaSelenastrumcapricornutum[J].Journal of Environmental Engineering Technology,2017,7(3):348-356.
