生物耕作对崇明芋艿养分吸收及其分配规律的影响
2017-05-19李双喜何七勇郑宪清张娟琴袁大伟张翰林吕卫光
李双喜,何七勇,郑宪清,张娟琴,袁大伟,张翰林,吕卫光
(上海市农业科学院生态环境保护研究所,农业部上海农业环境与耕地保育科学观测实验站,上海市设施园艺技术重点实验室,上海市农业环境保护监测站,上海 201403)
生物耕作对崇明芋艿养分吸收及其分配规律的影响
李双喜,何七勇,郑宪清,张娟琴,袁大伟,张翰林,吕卫光
(上海市农业科学院生态环境保护研究所,农业部上海农业环境与耕地保育科学观测实验站,上海市设施园艺技术重点实验室,上海市农业环境保护监测站,上海 201403)
通过田间试验,研究了生物耕作(接种蚯蚓)对芋艿N、P、K、Ca、Mg吸收及在体内分配的影响。结果表明:不同耕作方式下,芋艿植株不同器官对N、P、K、Ca、Mg等元素的吸收和分配规律是一致的。与传统机械耕作(CK)相比,生物耕作(T2)可以大幅促进N、P、K、Ca、Mg等元素在芋艿体内的积累(P<0.05),幼苗期以及发棵期(出苗后60 d)的N、P、K分配均为叶柄>叶片>根,Ca、Mg分配为叶片>叶柄>根;发棵期(出苗90 d)以及球茎膨大期的N、P、K主要分配在球茎中,子芋和孙芋中N、P、K含量最高,Ca素和Mg素则主要分布在子芋和叶片内,其他器官较少。在传统的农业生态系统中,培育土壤有益动物(生物耕作)可提高养分利用效率,对实现芋艿产业可持续生产具有重要意义。
生物耕作;蚯蚓;芋艿;矿质养分;吸收分配
芋[Colocasia esculenta(L.)schott]是天南星科多年生草本植物,是我国长三角地区典型的特色蔬菜之一,具有较高的营养价值和保健价值。香酥芋在崇明县有较长的栽培历史,肉质细腻,煮而不糊,是崇明县十大特色农产品之一,深受上海市民喜爱。目前,上海崇明地区香酥芋的栽培面积有166 hm2左右,由于其具有较好的效益,种植面积逐年增加。前人关于芋的研究主要集中在种质资源分布、品种划分、遗传多样性分析、营养成分分析、芋艿繁殖以及相关传统栽培技术等方面[1-5],关于芋艿的养分利用及分配研究较少,如宋春凤等[6]研究了传统栽培条件下莱阳孤芋的氮磷钾养分吸收状况。目前芋艿栽培全凭农户的传统经验,由于长期连作栽培,芋艿田出现了连作障碍问题[7],研究人员发现生物耕作(释放蚯蚓)可以有效缓解芋艿的连作障碍问题(另文发表)。为此,本试验比较了不同耕作方式下芋艿对N、P、K、Ca、Mg等矿质养分的吸收分配规律,以期为优化崇明地区芋艿的生产方式提供依据。
1 材料与方法
1.1 试验区概况
试验区设置在上海市崇明县三星镇西新村,试验区所在的崇明岛地处北亚热带,具有显著的季风气候特征。年平均气温15.3℃,降水量1 003.7 mm,日照时数2 104 h,日照百分率47%,全年无霜期229 d。试验田主要的蔬菜茬口为芋艿-花椰菜模式。生物耕作接种的蚯蚓为‘沪地龙’,为威廉腔环蚓,每条鲜重4 g左右,接种蚯蚓为15条/m2。蚯蚓种源由上海富年药材有限公司提供[8]。
1.2 试验设计
供试芋艿材料为崇明本地种香酥芋。试验设机械旋耕(CK)、免耕(T1)以及生物耕作(T2)3个处理,每处理重复3次,小区面积134 m2,随机排列。免耕即不采取任何机械耕作措施;机械旋耕深度为15 cm;生物耕作接种蚯蚓为15条/m2。为了防止蚯蚓由接种小区向其他处理小区迁移,小区之间开有水沟做天然屏障[8]。试验小区所用肥料为练科牌商品有机肥(N+P2O5+K2O>5%),基肥施用量为30 t/hm2。试验跨度为6个月(2014年4—10月),采样从5月持续到10月。
1.3 调查及取样方法
芋艿出苗后每隔30 d取样1次,每次取5株,重复3次。样品洗净后,按叶、叶柄、根、母芋、子芋、孙芋等器官分开,105℃杀青15 min,在75℃下烘干至恒重。将干样磨碎过筛后,用凯氏定氮法测全氮,钼锑抗显色法测全磷,原子吸收分光光度计测全钾、全钙、全镁[9]。
1.4 数据处理
使用Excel 2007和SPSS 16.0软件进行统计分析,P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 不同耕作方式下芋艿植株不同器官氮素积累及分配特点
不同处理下,芋艿植株不同器官对氮素的吸收和分配规律是一致的。随着生育期的进程,氮素在叶片、叶柄、根、母芋、子芋以及孙芋中的积累量逐步增加。但在不同时期,各器官氮素的分配有一定的差异,表现为:幼苗期,叶柄>叶片>根;发棵期(出苗后60 d),叶片>叶柄>根;发棵期(出苗后90 d),子芋>叶片>叶柄>母芋>根>孙芋;球茎膨大期,子芋>孙芋>母芋>叶片>叶柄>根。
生物耕作(T2)各个时期植株不同器官中氮素含量与机械耕作(CK)均有显著差异。与免耕处理(T1)相比,在发棵期(出苗后90 d),生物耕作大幅促进了孙芋对氮素的吸收和分配,差异呈极显著。从表1可知,在发棵期,超过50%的氮素分配到芋艿叶片中,在球茎膨大期,地上部(叶片和叶柄)氮素分配率逐渐减少,子芋和孙芋含量逐渐增加,CK、T1、T2处理地上部(叶片和叶柄)氮素分配率依次为21.7%、21.8%和22.1%,3种处理芋球茎中氮素占植株氮素的比例依次为76.3%、76.33%和76.42%。生物耕作(T2)促进了芋艿对氮素的吸收和分配,可能是因为蚯蚓活动增加了包括土壤氨化和硝化细菌在内的微生物数量[10-11],土壤中氨化细菌数量的增加,使更多不能被植物所利用的有机含氮化合物转化为可给态氮,也为植物及其他微生物的繁殖和活动创造了良好的营养条件[12]。此外,氮素是叶绿素的重要组成成分,芋艿发育前期叶片进行光合作用满足代谢需求,因而发棵期(苗后60 d)之前都是叶片中含量最高;随着生育期的过程,叶片生长基本稳定,生长重心转移到地下部(子芋和孙芋),氮素等营养元素转移运输到生长最旺盛的部位,因而造成在球茎膨大期氮素分配的差异。

表1 不同耕作方式对各个生育期芋艿植株各器官氮素(N)养分分配的影响Table 1 Effect of different tillage ways on nitrogen distribution in different taro organs at different grow th stages mg·株-1
2.2 不同耕作方式下芋艿植株不同器官磷素积累及分配特点
芋艿植株对磷素的积累及分配与氮素有相似之处。磷素是细胞核的主要组成部分,同时参与植物生长所需的各种酶的组成。磷素首先向生长中心运输和转移,并且具有较大的移动性[13]。如表2所示,随着生育期的过程,不同处理磷素在叶片、叶柄、根、母芋、子芋以及孙芋中的积累量逐步增加。但是在不同时期,各器官磷素的分配有一定的差异,表现为:幼苗期,叶柄>叶片>根;发棵期(出苗后60 d),叶柄>叶片>根;发棵期(出苗后90 d),子芋>叶柄>叶片>母芋>孙芋>根;球茎膨大期,子芋>孙芋>母芋>叶片>叶柄>根。

表2 不同耕作方式对各个生育期芋艿植株各器官磷素(以P2O5计)养分分配的影响Table 2 Effect of different tillage ways on phosphorus distribution in different taro organs at different grow th stages mg·株-1
生物耕作(T2)各个时期植株不同器官中磷素含量与机械耕作(CK)均有显著差异。与免耕处理(T1)相比,在发棵期(出苗后90 d),生物耕作大幅促进了子芋对磷素的吸收和分配,差异呈极显著。从表2可知,在发棵期(出苗后60天),超过50%的磷素分配到芋艿叶片中,在发棵期至球茎膨大期,地上部(叶片和叶柄)磷素分配率逐渐减少,子芋和孙芋逐渐增加,说明生长中心下移,磷优先运往地下部位;球茎膨大后期叶片衰老,叶中磷素开始外运,尤以球茎膨大期(出苗后150 d)T2处理的子芋磷素增加幅度最为明显,分别比CK和T1处理多46 mg/株和22 mg/株,这是因为生物耕作增加了土壤中无机磷分解菌的数量[11],促使土壤中的不溶性无机磷转化为可溶性磷盐,从而促进了磷素被植物吸收[14]。
2.3 不同耕作方式下芋艿植株不同器官钾素积累及分配特点
钾是植物生长所必需的一种成分。植物通过根系从土壤中选择性地吸收土壤中的水溶态钾离子,钾元素比较集中地分布在植物代谢最活跃的器官和组织中。如表3所示,随着生育期的进程,不同处理钾素在叶片、叶柄、母芋、子芋以及孙芋中的积累量逐步增加,根中的钾素积累量呈现先增加后减少的特点。但是在不同时期,各器官钾素的分配有一定的差异,表现为:幼苗期以及发棵期(出苗后60 d),叶柄>叶片>根;发棵期(出苗后90 d),叶柄>子芋>叶片>母芋>根>孙芋;球茎膨大期,孙芋>子芋>叶柄>叶片>母芋>根。
生物耕作(T2)处理除球茎膨大期(出苗后150 d)根中钾素差异不显著外,其他时期以及不同器官内钾素含量均与机械耕作(CK)差异显著。由表3可知,钾素在幼苗期以及发棵期(出苗后60 d)多分布在叶片和叶柄中,以叶柄为主,占全株钾素含量的60%以上。随着生育期的延长,尤其是发棵期(出苗后90 d)子芋中钾素急剧增加,至球茎膨大期,孙芋中钾素含量最高,分配最多,生物耕作(T2)处理表现的尤其明显。

表3 不同耕作方式对各个生育期芋艿植株各器官钾素(以K2O计)养分分配的影响Table 3 Effect of different tillage ways on potassium distribution in different taro organs at different grow th stages mg·株-1
2.4 不同耕作方式下芋艿植株不同器官钙素积累及分配特点
钙是形成果胶钙的主要成分,在作物体内果胶钙有助于细胞壁的发育,能促进作物体内细胞的分裂,并对碳水化合物的转化和氮素代谢有良好效果。如表4所示,随着生育期的进程,不同处理钙素在叶片、母芋以及子芋中的积累量逐步增加,叶柄、根以及孙芋中的钙素积累量呈现先增加后减少的特点,均在出苗后120 d达到最高。不同时期各器官钾素的分配有一定的差异,表现为:幼苗期以及发棵期(出苗后60 d),叶片>叶柄>根;发棵期(出苗后90 d),叶片>叶柄>子芋>根>母芋;球茎膨大期(出苗后120 d),叶片>叶柄>子芋>孙芋>母芋>根;球茎膨大期(出苗后150 d),叶片>叶柄>子芋>母芋>孙芋>根。
在球茎膨大期生物耕作(T2)处理叶片中钙素基本保持不变,而免耕(T1)和机械耕作(CK)呈增加趋势,在球茎膨大期(出苗后120 d)生物耕作处理叶片中的Ca含量最高达到419 mg,显著高于T1和CK处理;此外,从出苗后30 d到成熟期,芋艿吸收的钙素有45%以上分配在叶片中,其次分配在叶柄中,子芋比母芋和孙芋分配的多。
2.5 不同耕作方式下芋艿植株不同器官镁素积累及分配特点
镁是叶绿素的主要成分,能促进磷酸酶和葡萄糖转化酶的活化,有利于单糖的转化,因而在碳水化合物代谢过程中起着很重要的作用。如表5所示,随着生育期的进程,不同处理镁素在子芋和孙芋中的积累量逐步增加,叶片、叶柄、根以及母芋中的镁素积累量呈现先增加后减少的特点,均在出苗后120 d达到最高。不同时期各器官钾素的分配有一定的差异,表现为:幼苗期以及发棵期(出苗后60 d),叶片>叶柄>根;发棵期(出苗后90 d),叶片>叶柄>子芋>母芋>根;球茎膨大期(出苗后120 d),叶片>叶柄>子芋>孙芋>母芋>根;球茎膨大期(出苗后150 d),子芋>叶片>孙芋>叶柄>母芋>根。
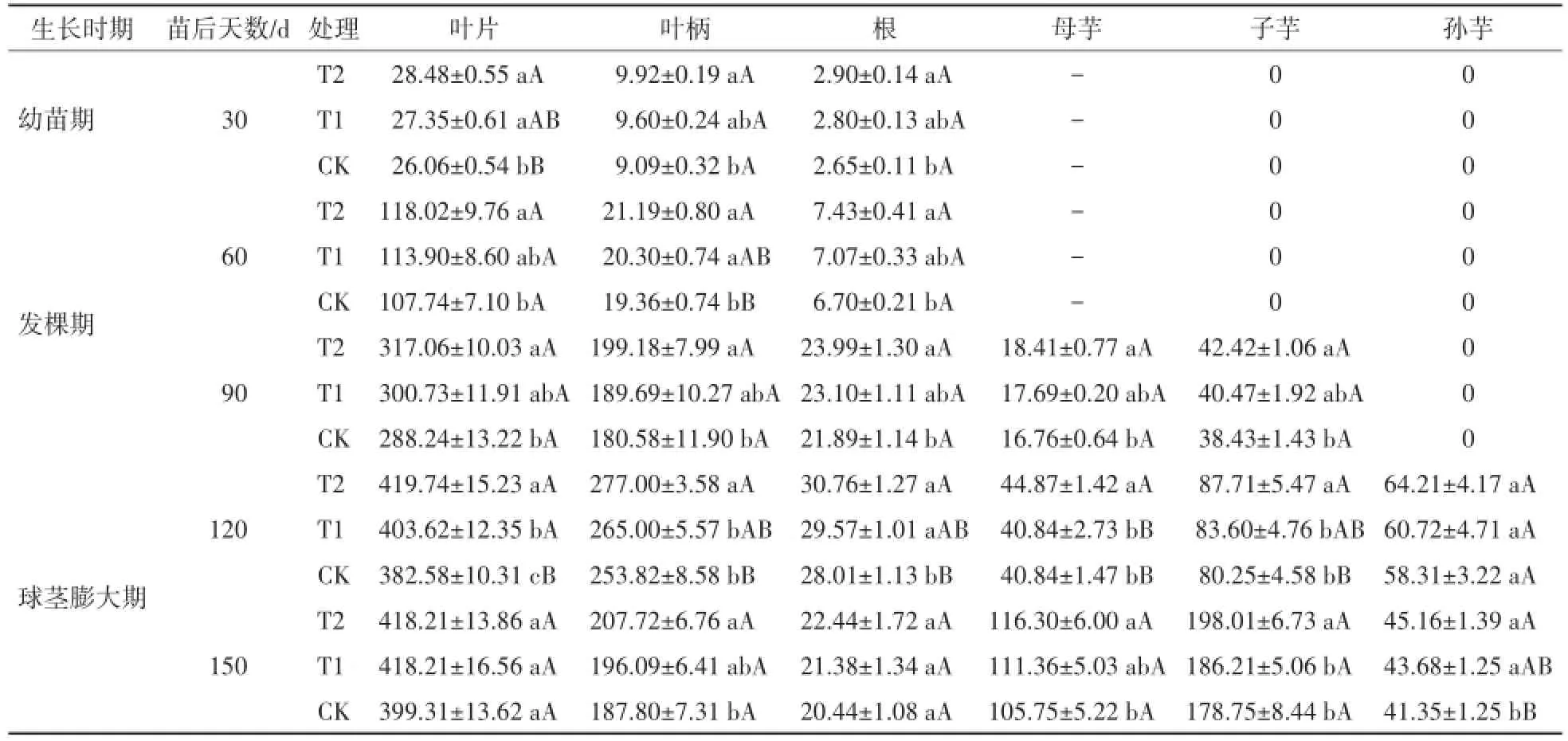
表4 不同耕作方式对各个生育期芋艿植株各器官钙素(Ca)分配的影响Table 4 Effect of different tillageways on calcium distribution in different taro organs at different grow th stages mg·株-1

表5 不同耕作方式对各个生育期芋艿植株各器官镁素(M g)分配的影响Table 5 Effect of different tillagewaysonmagnesium distribution in different taro organs at different grow th stages mg·株-1
在各个时期,生物耕作(T2)处理芋艿各个器官的镁素含量与机械耕作(CK)均有显著差异,而与免耕(T1)处理差异不显著。芋艿吸收镁素主要分配在叶片中,收获期子芋中分配比叶片多,生物耕作能够大幅提升芋艿各个器官对镁素的吸收和积累。
3 结论与讨论
本研究显示,在不同的耕作方式下,N、P、K、Ca、Mg等元素在芋艿不同器官内的含量和分配随生育期的延长有很大变化。与传统机械耕作(CK)相比,生物耕作(T2)可以大幅促进N、P、K、Ca、Mg等元素在芋艿体内的积累与分配,促进芋艿的生长发育。
就分配特点和规律而言,3种耕作方式处理下N、P、K、Ca、Mg等元素的分配规律是一致的。但是不同器官的分配规律有一定的差异,叶片、叶柄、母芋、子芋以及孙芋的N、P、K养分含量保持增加趋势,但是不同生育期Ca和Mg差异较大。幼苗期以及发棵期(出苗后60 d)的N、P、K分配均为叶柄>叶片>根,Ca、Mg分配为叶片>叶柄>根;发棵期(出苗90 d)以及球茎膨大期的N、P、K分配主要表现为地下部>地上部,根系分配最少,Ca素含量则表现为地上部>地下部,Mg素则主要分布在子芋和叶片内,其他器官较少。
合理的耕作措施可以提高土壤的养分,调节土壤pH、水分等,对植物的生长有很大促进作用[8]。前期的研究显示,释放蚯蚓可以改善土壤微生物活性、酶活性等,土壤酶积极参与土壤中氮、磷等矿化过程,从而促进其吸收利用[15-16]。释放蚯蚓(生物耕作)后,它的代谢产物与土壤微生物相互作用促进氮素的转化吸收和利用,Ca素和Mg素的吸收与氮素有很大的相关性,本研究也证明了生物耕作(T2)处理下芋艿各器官的Ca素和Mg素含量均高于T1和CK处理。因此,释放有益土壤动物(蚯蚓)进行生物耕作,可以更高效地利用土壤养分,改善传统芋艿生产过程中养分利用效率低下的问题,从而实现芋艿产业的可持续发展。
[1]黄新芳.中国芋种质资源研究进展[J].植物遗传资源学报,2005,6(1):119-123.
[2]沈镝.芋种质资源的耐藏性调查[J].长江蔬菜,2002(12):37-38.
[3]李庆典,杨永平,李颖.中国芋种质资源的遗传多样性及分类研究[J].湖南农业大学学报(自然科学版),2004,30(5):424-428.
[4]IRWIN S V,KAUFUSIP,BANKS K,et al.Molecular characterization of taro(Colocasia esculenta L.schott)using RAPD markers[J].Euphytica,1998,99:183-189.
[5]CAILLON S,QUERO-GARCIA J,LESCUREIJP,et al.Nature of taro(Colocasia esculenta(L.)schott)genetic diversity prevalent in a Pacific Ocean island,Vanua Lava,Vanuatu[J].Genetic Resources and Crop Evolution,2006,53:1273-1289.
[6]宋春凤,徐坤.芋对氮磷钾吸收分配规律的研究[J].植物营养与肥料学报,2004,10(4):403-406.
[7]杜红梅,黄丹枫.芋艿品种退化的原因及解决途径[J].中国蔬菜,2002(6):58-60.
[8]李双喜,郑宪清,袁大伟,等.生物耕作对菜田土壤理化特性、酶活性及青花菜生长和品质的影响[J].中国生态农业学报,2012,20(8):1018-1023.
[9]中国科学院南京土壤研究所.土壤理化分析[M].上海:上海科学技术出版社,1981:62-142.
[10]胡锋,王霞,李辉信.等.蚯蚓活动对稻麦轮作系统中土壤微生物量碳的影响[J].土壤学报,2005,42(6):965-969.
[11]李双喜,郑宪清,袁大伟,等.生物耕作对菜田土壤微生物区系及细菌生理类群的影响[J].华北农学报,2012,27(增刊):339-344.
[12]BILAL R.Associative of nitrogen-fixing plant growth promoting rhisobacteria(PGFPR)with kallar grass rice[J].Plant and Soil,1997,194:37-44.
[13]白宝璋,田纪春,王清连.植物生理学[M].北京:中国农业科技出版社,1996.
[14]CHEN Y P,REKHA P D,ARUN A B,et al.Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities[J].Applied Soil Ecology,2006,34:33-41.
[15]CECCANTIB,GARCIA C.Coupled chemical and biochemicalmethodologies to characterize a composting process and the humic substances[M]∥Senesi N,Miano T.Humic substances in the global environment and its implication on human health.New York:Elsevier,1994.
[16]胡佩,刘德辉,胡锋,等.蚓粪中的植物激素及其对绿豆插条不定根发生的促进作用[J].生态学报,2002,22(8):1211-1214.
(责任编辑:闫其涛)
Effects of biological tillage on nutrient absorption and distribution of Chongm ing taro
LIShuang-xi,HE Qi-yong,ZHENG Xian-qing,ZHANG Juan-qin,YUAN Da-wei,ZHANG Han-lin,LYUWei-guang
(Institute of Eco-Environment and Plant Protection,Shanghai Academy of Agricultural Sciences;Shanghai Scientific Observation and Experimental Station for Agricultural Environment and Land Conservation,Ministry of Agriculture;Shanghai Key Laboratory of Protected Horticultural Technology;Shanghai Environmental Protection Monitoring Station of Agriculture,Shanghai201403,China)
The effect of biological tillage(earthworm)on N,P,K,Ca,Mg absorption and distribution of taro plant[Colocasia esculenta(L.)schott]was studied by field experiment.The results showed that the absorption and distribution of N,P,K,Ca,Mg in different organs of taro plant were similar under different tillage ways.Compared with CK,biological tillage(T2)could greatly promote the accumulation of N,P,K,Ca,Mg in taro plant(P<0.05).In seedling stage and tiller stage(60 days after emergence),the distribution of N,P and K were petiole>leaf>root,Ca,Mg were leaf>petiole>root.In tiller stage(90 days after emergence)and corm expansion stage,N,P and K weremainly distributed in the corm,the content of N,P and K were the highest in the first grade taro and the second grade taro;Ca and Mg were mainly distributed in the first grade taro and leaves,less in the other organs.In the traditional agricultural ecosystem,cultivation of soil beneficial animal(biological tillage)could improve the efficiency of nutrient utilization,which was has important in realizing sustainable production of taro industry.
Biological tillage;Earthworm;Taro;Mineral nutrition;Absorption and distribution
S632.3
:A
1000-3924(2017)02-037-06
10.15955j.issn1000-3924.2017.02.07
2015-11-04
上海市农口青年成长项目[沪农青字(2014)第1-25号];上海市长三角国内合作项目(14395810602);上海市科技兴农推广项目(沪农科推字(2013)第4-1号);国家科技支撑计划项目(2010BAK69B18)
李双喜(1986—),男,在读博士,助理研究员,主要从事生态农业、设施农田土壤障碍研究与防治工作。E-mail:lsx1986@126.com
