猪akirin2基因的组织表达谱及序列分析
2017-05-17郭豫杰高会贞李宏基王月影李奎杨
郭豫杰 高会贞 李宏基 王月影 李奎 杨国宇
摘要:克隆猪的akirin2基因,并对该基因在猪的18个组织中的分布进行研究。猪akirin2基因编码区全长为612 bp(GenBank登录号KCl40110.1),推测的氨基酸序列包含203个氨基酸,为酸性蛋白。利用BLAST工具对克隆到的猪akirin2基因序列与人、大鼠、小鼠、绵羊和牛的序列进行同源性比较。采用软件对氨基酸序列进行分析,用相关序列的比对结果进行系统进化树分析。组织分布结果显示,猪akirin2基因在脑组织中高表达,在淋巴、睾丸中表达量相对较低,在心脏、十二指肠、直肠、皮肤、胸腺中几乎检测不到该基因的表达。序列分析结果显示,猪akirin2基因与绵羊和牛的同源性最高,均为96%,与人、小鼠、大鼠的同源性分别为95%、91%、89%。氨基酸序列分析发现,存在1个10个肽的核定位信号序列19-PASPKRRRCA-28,推测的氨基酸序列的二级结构中含有2个a-螺旋。在重建的系统进化树上,几个不同物种的akirin2蛋白分成3支,结果表明猪akirin2与人、牛和羊的亲缘关系比其与小鼠和大鼠的近,与鸡的亲缘关系最远。
关键词:猪;akirin2基因;克隆;组织分布;序列分析;基因功能
中图分类号:S828.2 文献标志码:A 文章编号:1002—1302(2016)01—0025—04
在畜牧业中,肌肉的产量和质量是肉用畜禽的重要经济性状,对畜禽饲养业的经济效益起决定性作用。肌细胞的发育开始于成肌细胞或前体细胞。成肌细胞经过分化和融合过程形成肌管,其随后进一步分化成肌纤维。肌肉抑制素(my-ostatin,MSTN)(或生长分化因子8,GDF-8)是一种调节肌肉生长和发育的主要因子,MSTN表现出对肌肉生长的负调节。肌肉抑制素基因中11 bp的缺失在牛中导致比利时蓝(belgian blue)或双臀肌(double-muscled)表型,比利时蓝牛在肌肉质量上增加20%~30%。尽管在遗传分析中已经揭示出MSTN的重要功能,然而对MSTN信号通路下游调控肌肉发生的相关基因的了解还不多。akirin是Miner等通过小鼠消减抑制杂交文库筛选出的MSTN下游调控基因。这一新核蛋白基因的发现,填补了MSTN调控肌肉生成的上下游基因的空缺。在脊椎动物中akirin基因非常保守,至少存在akirin1和akirin2等2个同源基因。然而,在鸟类和爬行动物中没有akirin1,只有akirin2基因,在硬骨鱼中akirin基因家族有2~8个成员,在细菌、植物和酵母中均未发现有akirin的存在。张志强等首次克隆到猪的mighty(akirin1)基因,并对该基因在猪不同组织中的表达谱进行分析。本研究克隆猪的akirin2基因,并对其组织分布及序列特征进行分析,为进一步研究该基因的功能奠定基础。
1材料与方法
1.1样品采集
取健康猪的心、肝、脾、肺、肾、空肠、十二指肠、回肠、直肠、大脑、肌肉、皮肤、脂肪、胸腺、淋巴、下颌、子宫、睾丸等18个组织,用锡箔纸包好迅速冷冻于液氮中,置于-80℃低温冰箱保存备用。
1.2主要试剂与仪器
Trizol(Invitrogen);DEPC(Serva);Reverse TranscriptaseM-MLV(RNase H_)、RNase inhibitor、dNTP-mix、Premix TaqVersion 2.0、DL2000 DNA Marker、凝胶回收试剂盒(TaKaRa Agarose Gel DNA Purification Kit Ver.2.0)、pMDl9-T Vector均购自宝生物工程(大连)有限公司;大肠杆菌(Escherichiacoli)DH5ct菌株为笔者所在实验室保存。台式高速离心机(Eppendorf,Germany);温度梯度PCR仪(sensoquest,Germa-ny);凝胶成像系统(Alphalmager HP,USA);微量核酸蛋白检测仪NanoDrop-100(Thermo,USA)。
1.3方法
1.3.1引物设计 根据NCBI上GenBank中小鼠和人的aki-rin2基因序列,在GenBank数据库中对猪的ESTs和UniGene数据库进行搜索,对搜索到的ESTs序列进行拼接,结合软件BIOXM2.6及引物设计软件,设计1对引物[由生工生物工程(上海)股份有限公司合成]:akirin2-F,5′-ATGGCGTGCG-GAGCTACTCTG-3′;akirin2-R,5′-TCATGAAACATAAC-TAGCAGGCTG-3′。
1.3.2总RNA的提取及cDNA的合成 取50~100 mg经液氮处理过的大脑组织,用Trizol法从中提取总RNA。取2μL样品,采用ND-1000分光光度计检测总RNA的浓度及纯度,取8μL进行甲醛变性胶电泳检测,其余总RNA样品-80℃冰箱中保存备用。反转录体系:0.5μg/μL Oligo(dT)151μL、总RNA 2μg、M-MLV Buffer(5×)5.0 IxL、10 mmol/LdNTP 6.0 txL、40 U/μL RNase-Inhibitor 0.7 txL、200 U/μLM-MLV Reverse Transcriptase 1.0μL、DEPC H2O补足至25.0μL。反应程序:25℃10 min,42℃60 min,72℃15 min。反应结束后,迅速冰浴2 min,得到cDNA,置于-20℃保存。
1.3.3目的基因豬akirin2的扩增 以上述cDNA稀释5倍作为模板,进行PCR扩增。PCR扩增体系:Premix Taq聚合酶12.0 μL、10μmoL/μL akirin2-F 1.0 μL、10μmol/μLakirin2-R 1.0μL、cDNA 2.0 txL,用灭菌的蒸馏水补足至总体积为25.0μL,混匀,瞬时离心,立即置于PCR仪上。反应程序:95℃预变性5 min;95℃30 s,60℃30 s,72℃40 s,30个循环;72℃延伸15 min。反应结束后,取5μLPCR产物用
1.2%琼脂糖凝胶电泳进行检测。
1.3.4猪akirin2基因的克隆 PCR产物经1.2%琼脂糖凝胶电泳进行切胶纯化,产物纯化回收采用TaKaRa Agarose Gel DNA Purification Kit Ver.2.0试剂盒进行。将回收的产物与pMD19-T载体直接进行连接,连接产物转化至感受态细胞DH5α,涂布于含有50μg/mL Amp的LB平板上,37℃培养过夜。挑取3~5个单克隆作菌液PCR鉴定,经初步鉴定为阳性的克隆送往生工生物工程(上海)股份有限公司测序。
1.3.5组织分布 提取采集的猪心、肝、脾、肺、肾、空肠、十二指肠、回肠、直肠、大脑、肌肉、皮肤、脂肪、胸腺、淋巴、下颌、子宫、睾丸等18个组织的总RNA,经RT-PCR后得到相应的cDNA。选择猪的β-actin基因作为内参,引物序列:β-actin-F,5′-GGACTTCGAGCAGGAGATGG-3′;β-actin-R,5′-AGGAAGGAGGGCTGGAAGAG-3′,产物片段预计为138 bp。用于扩增猪akirin2基因的引物序列:akirin2-F2,5′-TCCATCAGCAACGTCCTCAC-3′;akirin2-R2,5′-AACTAGCAGGCTGTTCTCCA-3′,扩增片段预计为218 bp。各取5μLPCR产物经1.2%琼脂糖凝胶电泳进行检测,分析猪akirin2在以上各组织中的分布及表达情况。
1.3.6序列分析 利用ncbi在线分析服务器软件Blast(http:∥www.ncbi.nlm.nih.gov/BLAST)进行序列同源性比对。应用ExPASy(http:∥expasy.ch/tools/protparam.html)在线分析推测氨基酸序列的理化特征。信号肽序列的预测采用signalP 3.0在线软件(http:∥www.cbs.dtu.dk/services/Signalp)。应用SABLE(http:∥sable.cchmc.org)在线软件对蛋白质的二级结构进行预测分析。
为了研究akirin基因在进化上的特点,选取NCBI中绵羊、牛、人、小鼠、大鼠以及鸡这几个物种的相关序列进行多序列比对,并用MEGA 5.0软件,选择邻接法(Neighbor-Join-ing)并自举(Bootstrap)1000次计算置信值,建立系统进化树。
2结果与分析
2.1总RNA的提取
肌肉组织总RNA采用Trizol法提取后,采用ND-1000分光光度计检测总RNA的浓度及纯度。D260nm/D2280nm为1.83,电泳条带以28S rRNA和18S rRNA为主(图1)。
2.2猪akirin2基因的克隆
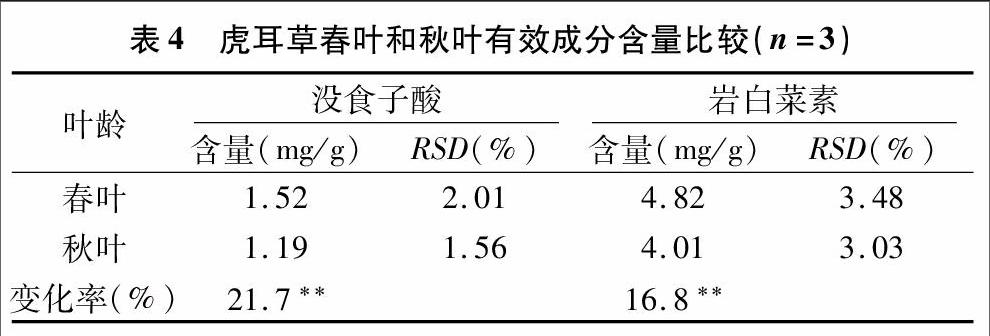
总RNA经反转录后,以得到的cDNA为模板,经PCR扩增后,取5μL PCR产物,经1.2%琼脂糖凝胶电泳检测,电泳结果显示,扩增得到1个约为609 bp的片段(图2),该片段大小与预期目的基因大小位置接近。
2.3菌液PCR鉴定
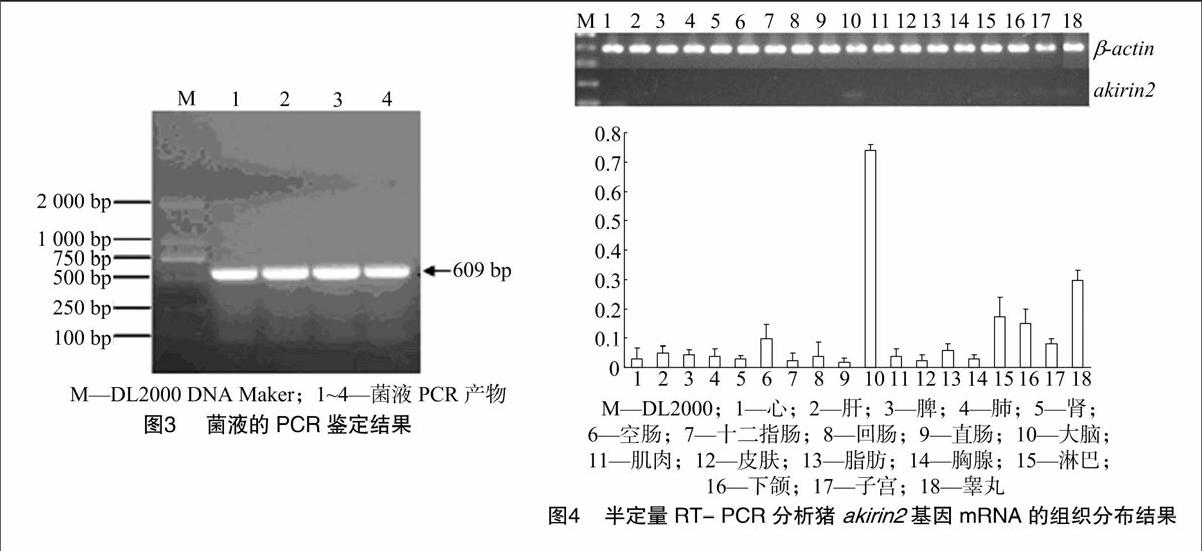
经切胶纯化回收后,将该DNA片段与pMD19-T载体进行连接、转化、涂板及挑斑后,经菌液PCR对阳性克隆进行初步筛选,菌液PCR结果(图3)显示,在609 bp处有预期大小的目的条带。
2.4 RT-PCR分析猪akirin2基因组织表达谱
为了分析猪akirin2基因mRNA在不同组织中的表达及分布情况,本研究提取猪18个组织的总RNA,采用半定量RT-PCR的方法进行分析。目的基因猪akirin2的光密度值经内参基因β-actin进行修正,采用二者的比值作为目的基因的相对表达水平。结果显示,猪akirin2基因在猪不同组织中的表达和分布明显不同(图4),在有些组织如心脏、十二指肠、直肠、皮肤、胸腺等中几乎检测不到该基因的表达,在脑组织中该基因有相对较高的表达水平,在淋巴、睾丸等组织中可以检测到akirin2基因低水平的表达。
2.5序列分析
在GenBank中利用Blast工具对克隆到的猪akirin2基因进行同源性比较,猪akirin2基因与绵羊(Ovis aries,NM_001252176.1)和牛(Bos taurus,NM_001110087)的akirin2基因同源性最高,均为96%,与人(Homo sapiens,NM_018064.3)的同源性为95%,与小鼠(Mus musculus,NM_001007589.3)的同源性为91%,与大鼠(Rattus norvegicus,NM_001039914.2)的同源性为89%。从比对结果可知,该基因在不同物种间的同源性较高。
通过ExPASy软件在线分析预测到该蛋白质相对分子量为50 900.1,理论等电点为5.12,为酸性蛋白。采用signalP3.0在线预测信号肽软件分析发现,该多肽链序列无信号肽,不是分泌蛋白,序列分析发现存在1个10个肽的核定位信号序列19-PASPKRRRCA-28(图5),由此推测akirin2可能在细胞核中发挥功能,这在鼠和果蝇akirin2的研究中已经得到证实。
应用SABLE在线软件对猪Akirin2蛋白质的二级结构进行预测分析,结果显示,二级结构中含有2个α-螺旋(图6)。其中,靠近C-端的α-螺旋在猪、牛、羊、人、小鼠、大鼠及雞中都有。
为了研究akirin2基因在进化中的特点,选取NCBI中绵羊、牛、人、小鼠、大鼠以及鸡这几个物种的相关氨基酸序列进行多序列比对,并用MEGA 5.0软件,选择邻接法(Neighbor-Joining)并自举(Bootstrap)1000次计算置信值,建立系统进化树(图7)。在重建的系统进化树上,这几个不同物种的Akirin2蛋白分成3支,牛和羊先聚在一起,然后又与猪和人聚成其中一支,小鼠和大鼠聚成另外一支,鸡则单独在一支上。结果显示,猪Akirin2蛋白与人、牛和羊的亲缘关系比其与小鼠和大鼠的近,与鸡的亲缘关系最远。
3结论与讨论
目前已经报道了akirin基因在免疫、发育及肌肉再生等方面发挥了重要的生物学功能。本研究克隆了猪akirin2基因的编码区全序列,该序列包含612个核苷酸,由此推测编码203个氨基酸。人、牛、羊的akirin2基因的ORF的核苷酸序列也都是612 bp,编码203个氨基酸;小鼠和大鼠的akirin2基因的ORF包含609个核苷酸,编码201个氨基酸,鸡的akirin2基因的ORF仅有573 bp,编码191个氨基酸,比猪、牛、羊和人的少36 bp,共计12个氨基酸。大鼠和小鼠的akirn2连续缺少6 bp,氨基酸序列中连续缺少2个氨基酸残基,鸡连续缺失36 bp、12个氨基酸,缺失部分都不在α-螺旋区。由于氨基酸的缺失使得猪Akirin2氨基酸的序列同源性與大鼠和小鼠较近;而鸡Akirin2氨基酸残基缺失,导致猪与鸡的同源性最远,而氨基酸数目同样多的猪、牛、羊和人的同源性更近。人、牛、羊、大鼠、小鼠和鸡的Akirin2蛋白在其N-端都有一段保守的核定位信号序列,序列分析发现,猪Akirin2蛋白与其他物种的Akirin2蛋白一样,也含有1个核定位信号序列,这与报道的Akirin2是在细胞核中发挥功能的核蛋白相符合。此外,这一核定位信号序列在猪Akirin1蛋白中也能找到。应用SABLE在线软件对猪Akirin2蛋白质的二级结构进行预测分析,结果显示,猪Akirin2的二级结构中含有2个α-螺旋,靠近C-端的α-螺旋在猪、牛、羊、人、小鼠、大鼠及鸡中都有,该区域的保守性显示此区域可能对Aki-rin2发挥功能具有重要作用。
从猪akirin2基因组织分布的试验结果中可以看出,该基因在不同组织中的分布和表达水平明显不同。检测的18个组织中,心脏、十二指肠、直肠、皮肤、胸腺等组织中几乎检测不到该基因的mRNA表达,只在大脑中检测到相对较高的表达。Man等在研究不同日龄鸡的akirin2基因的组织表达谱时发现,在雏鸡和成年鸡16个组织中的akirin2基因表达存在差异,在3日龄鸡的11个组织肝脏、胸腺、肾脏、肌胃、心脏、皮肤、大脑、小肠、脾脏、肺和肌肉中该基因均有明显的表达,然而在成年鸡的某些组织如心脏、肝脏、大肠、腹脂和胰腺中则几乎检测不到该基因的表达,说明该基因的表达具有一定的时间特异性。该试验在雏鸡和成年鸡的大脑中都检测到了较高水平的akirin2 mRNA,这与本研究在猪的脑组织中检测到较高水平akirin2基因的表达结果一致。此外,猪akirinl(mighty)基因在脑中也显示有较高水平的表达。脑中akirin基因相对较高的表达水平具有的意义仍需要试验进行深入研究。与猪akirin1(mighty)基因在猪不同组织中的表达谱相比较发现,akirin1基因在各种组织中都有表达,而akirin2只在少数几个组织中检测到有表达,由于本研究没有检测蛋白水平的表达情况,导致猪akirin2基因表达差异的可能原因较多,由此推测猪akirin2在不同组织中的分布差异可能与该基因在不同组织中的功能有关。本研究成功克隆了猪的akirin2基因,并对其序列进行分析,检测猪akirin2基因在18个组织中的分布和表达情况,这些数据为进一步研究猪akirin2基因的功能奠定基础。
