凋落物对土壤呼吸的贡献研究进展①
2017-05-15吕富成王小丹
吕富成,王小丹
(1 中国科学院成都山地灾害与环境研究所,成都 610041;2 中国科学院大学,北京 100049)
凋落物对土壤呼吸的贡献研究进展①
吕富成1,2,王小丹1*
(1 中国科学院成都山地灾害与环境研究所,成都 610041;2 中国科学院大学,北京 100049)
土壤呼吸是土壤碳库输出的主要途径,凋落物是影响土壤呼吸的重要因素。明确凋落物对土壤呼吸的贡献,有助于准确评估植物-土壤-大气三个碳库之间的碳收支过程。本文综述了近年来国内外有关凋落物对土壤呼吸贡献的研究成果,阐明了凋落物对土壤呼吸的贡献机理,讨论了凋落物对土壤呼吸贡献率及其存在的时空分异特征,在此基础上,对该领域研究前景进行了展望。
凋落物;土壤呼吸;碳循环
土壤是陆地生态系统最大的碳库,碳储量达1.5×103Pg(以单质C计,下同),约为大气碳储量的2倍,陆地植物碳储量的3倍[1–2]。土壤呼吸是指土壤碳以 CO2的形式向大气释放的过程,是土壤碳库输出的主要途径[3]。土壤每年向大气输出碳约68 Pg,约占大气CO2输入量的20% ~ 40%,所以土壤呼吸轻微的扰动对全球大气CO2浓度的变化将产生重大影响[4–8]。
凋落物是陆地生态系统地上部分和地下部分物质和能量交换的重要“纽带”[9]。作为土壤碳库输入的主要来源之一[10],凋落物还为土壤微生物活动提供物质基础,调节土壤表层温度和湿度,改善土壤微环境,进而影响土壤呼吸[11]。全球变化和人类活动引起的气温升高、氮沉降以及土地利用/覆被变化等问题,势必会对陆地生态系统的生产力[12]、凋落物质量[13]、凋落物量[14]产生影响。凋落物这个碳库的扰动,则会对全球气候变化产生反馈作用[15–16]。因此,厘清连接土壤和大气纽带的凋落物对土壤呼吸的贡献是十分必要的。本文在综述了国内外相关研究成果基础上,分析了凋落物对土壤呼吸的贡献机理,并总结了凋落物对土壤呼吸贡献率存在的时空规律及其调控因子,在此基础上,对该领域的研究前景提出了展望,以期为该领域今后的研究提供参考。
1 凋落物对土壤呼吸的贡献机理
凋落物对土壤呼吸的贡献机理包含了一系列的生物学过程,它可以通过直接或者间接途径影响土壤呼吸。凋落物对土壤呼吸的直接贡献主要是指凋落物分解过程中释放的 CO2;间接贡献是指凋落物通过物理过程、化学过程和生物过程屏蔽气体传输、改变土壤水热条件、增加或者减少土壤碳源,影响根系生长和微生物群落的结构与功能等,进而作用于土壤呼吸。
1.1 凋落物对土壤呼吸的直接贡献
凋落物对土壤呼吸最直接的贡献是在微生物的参与下,凋落物通过自身分解释放CO2。但许多研究表明,凋落物释放CO2通量低于凋落物对土壤呼吸的贡献量[17–19]。例如,鼎湖山3种植被季风常绿阔叶林、针阔叶混交林、马尾松林凋落物年平均直接排放的CO2通量分别为597±129、736±187、582±181 g/(m2⋅a) (以单质C计,下同),明显低于凋落物对土壤呼吸的贡献量[17]。另外,原始林凋落物释放的 CO2通量高于更新林,这主要是林地凋落物多寡及各层所占比例不同所致[18]。这表明凋落物自身分解释放 CO2量并不能完全代表凋落物对土壤呼吸的贡献。
1.2 凋落物对土壤呼吸的间接贡献
凋落物对土壤呼吸的间接贡献包括物理过程、化学过程和生物过程,3种过程相互关联,共同作用。
1) 物理过程。凋落物地上部分是大气与土壤进行物质与能量交换的缓冲区,其物理过程包括两方面:第一,凋落物层降低了大气对土壤内部环境的扰动,特别是有效降低土壤温度和湿度的波动幅度[20],有利于土壤微生物和真菌的生存。研究证实,去除凋落物降低了Q10值,削弱土壤对环境变化的抗干扰能力[17,21]。因此,凋落物层是土壤碳库抵御外界气候变化重要的组成部分,也是凋落物对土壤呼吸贡献的物理表现形式。第二,覆被于地表的凋落物层减缓了土壤呼吸向外释放CO2的过程,被称为“屏蔽效应”[18–19,22],造成这种效应的原因:一是某些生态系统凋落物覆盖层很厚,在土壤呼吸中扮演屏障角色,物理减慢了 CO2的外溢过程;二是土壤微生物中好气性细菌比例很高,去除凋落物层后活动异常旺盛所致[19];三是凋落物以CO2形式排放的碳量相当低[18],主要以别的形式转化和迁移。当屏蔽作用阻挡的气体量超过凋落物分解释放量,凋落物对土壤呼吸的贡献就会出现负值[23–24],如三江平原退耕还湿地的凋落物对土壤呼吸的贡献率为 –5%[24]。
2) 化学过程。凋落物对土壤呼吸间接贡献的化学过程在于:凋落物是土壤有机质的主要来源,在微生物等分解者的参与下,凋落物转化为供植物吸收利用的矿质养分,促进土壤有机碳的化学氧化和植物根系呼吸,影响根系生长和分配。例如,Sayer等[25–26]对热带森林的研究发现,增加凋落物量导致植物细根量显著减少,土壤呼吸反而增加,原因是新鲜凋落物的输入带来更为有效的底物碳源[27–28]或者加倍凋落物产生了正“激发效应”[29–30]。凋落物的化学性质也会影响凋落物的分解速率,凋落物的质量(碳、氮含量,酚含量,碳/氮,木质素/氮,氮/磷等)是决定凋落物分解速率的重要因素[31–32],继而对土壤呼吸有重要的影响。其中,凋落物分解与碳/氮、木质素/氮值关系最为密切[31]。一般而言,凋落物碳/氮比越高,氮含量越低,木质素含量越高,凋落物分解越困难[32]。但是氮与木质素对凋落物分解的限制作用在不同分解阶段上有所差异,初期起主导作用的是氮,后期起主导作用的是木质素[33]。另外,研究发现凋落物碳/氮比值低的森林土壤中微生物活性较高[26],这是由于低的碳/氮比值有利于微生物的分解。
3) 生物过程。凋落物对土壤呼吸贡献的生物过程表现为凋落物地上部分和地下部分的输入会影响土壤微生物群落的结构与功能[34–36]。微生物是土壤养分循环的重要参与者和推动者,不同凋落物的输入均能影响土壤微生物的活性和代谢能力[37]。研究认为,去除和添加凋落物处理会改变土壤真菌和细菌比例[38–39]。Subke等[40]利用同位素标记技术对挪威云杉进行环剥和凋落物去除实验,结果显示在无环剥保留凋落物的条件下,真菌生物量和微生物碳含量最大,对土壤呼吸产生“激发效应”[29]。Li 等[41]测定去除凋落物使风铃木次生林和松树人工林土壤微生物量分别减少67% 和69%,微生物的活性也相对降低。相对于物理过程和化学过程,凋落物对土壤呼吸贡献的生物过程研究较少。
2 凋落物对土壤呼吸的贡献率
凋落物对土壤呼吸的贡献率是衡量凋落物在土壤碳循环中作用的重要指标之一,地上/地下凋落物对土壤呼吸的贡献率在一定程度上可以反映植物地上部分与地下部分碳分配机制。因此,深入研究凋落物对土壤呼吸的贡献率对于认识凋落物在植物与土壤之间物质循环和养分平衡中的纽带作用具有十分重要的意义。
2.1 地上凋落物对土壤呼吸的贡献率
目前,地上凋落物对土壤呼吸贡献率的研究通常采用DIRT(detritus input removal and transfer)方法[39],即在尽量相邻两块样地中,分别做保留地上凋落物和去除地上凋落物的处理并测定两块样地土壤呼吸总量,二者之差即为地上凋落物对土壤呼吸的贡献量,然后其贡献量除以保留地上凋落物的土壤呼吸量即为地上凋落物对土壤呼吸的贡献率,其计算方法为:
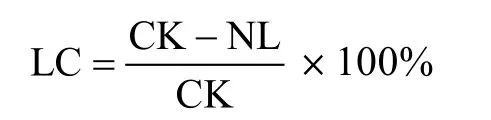
式中:LC为地上凋落物对土壤呼吸的贡献率,CK和NL分别为保留和去除地上凋落物处理的土壤呼吸(g/m2)。
综合当前的研究成果发现,地上凋落物对土壤呼吸的贡献率为7.6% ~ 40.8% 不等(表1),且地上凋落物对土壤呼吸的贡献率存在明显的时空变异,并受到水热条件[47]、凋落物质量[57]、凋落物输入量[58]、土壤动物和微生物[49]等多种因素的综合调控。
首先,地上凋落物对土壤呼吸的贡献具有明显的季节动态,且不同地区的水热调控因素存在差异。例如,邓琦等[17]对 3种主要类型的森林凋落物研究得出,季风常绿阔叶林、针阔叶混交林、马尾松林地上凋落物对土壤呼吸贡献的高峰期都出现在雨季(4—9月),分别占全年贡献量的 71.9%、61.9%、70.4%。在意大利地中海混交栎树林的调查也发现,受夏季高温干旱的影响,地上凋落物对土壤呼吸的贡献仅为15%,低于降水丰富的秋季(29%)[47]。这是因为在干湿分明的生态系统中,水分的胁迫作用往往会比温度更大,甚至居于主导地位[60]。研究表明,干燥环境会导致土壤微生物休眠或形成孢子[61],甚至细胞脱水[62],而适度增加水分可以提高土壤微生物的活性,增加土壤呼吸速率。例如,在亚热带森林的研究中发现,降水能显著提高地上凋落物对土壤呼吸的贡献率,且保留地上凋落物处理的土壤呼吸通量在降水天气和晴朗天气具有极显著差异,而去除凋落物的处理土壤呼吸则差异不大[17]。对地中海森林凋落物研究发现,微生物的活性与凋落物水分含量关系密切,在降水丰富的春季和秋季,凋落物分解高于高温少雨的夏季,并且真菌总量和凋落物质量损失率密切相关[63],这也是导致地中海混交栎树林地上凋落物对土壤呼吸贡献率秋季高于夏季的原因之一[46]。在雨热同期的季风气候区,地上凋落物对土壤呼吸的贡献在高温多雨的夏季达到最高,在低温少雨的春季地上凋落物贡献率最低[50],表明在该地区地表凋落物贡献受到温度和水分的共同作用。在我国东北地区,与降水量相比,地上凋落物对土壤呼吸的贡献与温度之间存在更加显著的相关性,尤其是与土壤温度的相关性更加显著[24,55]。例如,王丽丽等[24]研究了三江平原沼泽湿地不同土地利用方式下保留和去除凋落物的土壤呼吸特征,表明小章草甸湿地、人工林地、退耕还湿地3种凋落物对土壤呼吸的贡献均与10 cm的地温呈极显著关系。另外,在高海拔地区,温度或者温度和水分共同影响地上凋落物对土壤呼吸的贡献率[54]。

表1 不同植被地上凋落物对土壤呼吸的贡献率Table 1 Contribution rates of aboveground litters of different plants to soil respiration
其次,地上凋落物对土壤呼吸贡献率也有一定的空间变异规律。Chen等[64]通过对全球成熟林的研究发现,从寒温带、温带到热带,森林地上凋落物对土壤呼吸的贡献在逐渐升高,并且与年平均气温和降水呈正相关。在热带、亚热带森林,水热条件较好,微生物活动强烈[43,45,48],年均凋落物分解率可达到40%以上[45],因此地上凋落物对土壤呼吸的贡献率高(30% ~ 40%)。在温带地区水热条件较差,森林年均凋落物分解率为20% ~ 30%[66],地上凋落物贡献约占土壤呼吸的20%[47,51,67]。在寒温带地区,温度是环境因子中的限制性因素,微生物数量和活性受到制约,地上凋落物分解速率降低,导致地上凋落物贡献低于10%[20,68]。寒带冻原地区植物地上凋落物和腐殖质分解缓慢[69],凋落物对土壤呼吸影响并不大,但是随着全球气候变暖,可以预期冻原地区凋落物(有机质)会加速分解[70],其对土壤CO2的贡献率会有所升高。Raich等[56]研究证明,地上凋落物的贡献率与枯枝落叶输入量相一致。枯枝落叶输入量随着纬度升高逐渐减少[4],这也是造成凋落物对土壤呼吸贡献空间差异的原因。
2.2 地下凋落物对土壤呼吸的贡献率
由于已有的地下凋落物分解实验方法的局限性和地下凋落物分解的复杂性,使研究地下凋落物对土壤呼吸的贡献存在诸多不确定性,也就造成了地下凋落物对土壤呼吸贡献率的研究精度不高,差异较大。根据表2可知,地下凋落物对土壤呼吸的贡献在30% ~80%。
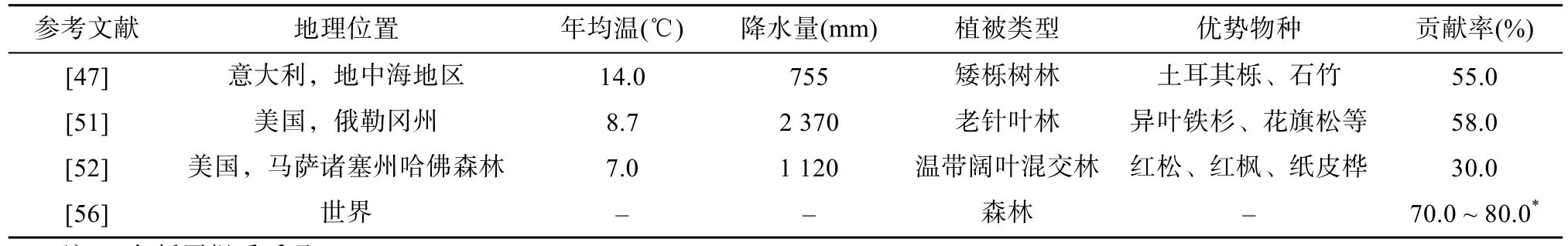
表2 不同植被地下凋落物对土壤呼吸的贡献率Table 2 Contribution rates of belowground litters of different plants to soil respiration
一般认为,土壤养分状况是造成地下凋落物对土壤呼吸贡献差异的主要原因。Bownen等[52]对哈佛森林的研究发现,地下凋落物对土壤呼吸的贡献为30%,低于其他地区的贡献率(表 2),造成差异的原因可能是哈佛森林土壤肥力较好,碳/氮比为23;而喀斯喀特山地区[51]土壤养分状况较差,碳/氮比为35.9,植物碳分配地下部分要高于地上部分[71–72],且根系凋落物周转速率快。植物的生长阶段也会影响地下凋落物的贡献。成熟林地下碳分配约为地上凋落物碳的两倍[56],例如,Davidson等[73]发现,幼龄林的根系呼吸和根系凋落物分解所释放的 CO2对土壤呼吸的贡献高于地上凋落物,这表明幼龄林碳分配更倾向于地下部分,以获取更多的养分和水分促进地上部分的快速生长[74]。菌根作为地下凋落物的来源也是影响地下凋落物分解释放 CO2的重要因素,菌根能扩大根系的吸收面积和吸收范围,增强宿主对养分的吸收利用[75]。Vogt等[76]研究发现,生长在养分较低土壤中的花旗松菌根感染率高达 88%,因此,有些地方地下凋落物输入可能不是直接源于根系,而是菌根[51]。
3 结语与展望
总之,地上凋落物对土壤呼吸的贡献率在10% ~40%,且具有明显的季节性和空间分异,地下凋落物对土壤呼吸的贡献研究较少,目前的研究认为地下凋落物对土壤呼吸的贡献率达30% 以上。凋落物对土壤呼吸的贡献机制包含了一系列的生物学过程,凋落物通过自身分解释放 CO2通量只是对土壤呼吸贡献的一部分,其还可以通过物理、化学和生物过程等间接途径影响土壤呼吸。
虽然凋落物对土壤呼吸的贡献研究已引起生态学界的重视,但目前的许多研究时间较短,实验设计较简单,缺乏长期性、机理性的系统研究。为此,今后的研究重点可围绕如下几个方面开展:
1) 地上/地下凋落物对土壤呼吸协同贡献及其各生态因子交互作用。植物地上生物量和地下生物量是相互联系、密不可分的整体,不同的土壤条件和生长阶段,植物碳分配策略存在差异。因此,应着重研究地上/地下凋落物分解对土壤呼吸的协同贡献;加强土壤水热动态、养分变化、微生物活性等生态因子的交互作用以及对凋落物分解的影响,尤其是明确土壤动物和微生物在凋落物分解中的作用。注意不同尺度的转换,凋落物对土壤呼吸的贡献可能在不同尺度下驱动因素不同。另外,需要关注生态系统演替与碳分配如何影响地上/地下凋落物对土壤呼吸贡献。
2) 开展凋落物–大气–土壤界面研究。凋落物不仅是陆地生态系统地上部分和地下部分物质和能量交换的重要纽带,而且位于土壤和大气的交界面。大气 CO2含量变化、氮沉降等全球变化会影响植物生物量分配,改变凋落物输入量,因此,开展凋落物输入量对全球变化的反馈机制十分必要。另外还有几个问题需要进一步明确,凋落物的“屏蔽效应”是否存在临界值?“屏蔽效应”、“激发效应”和凋落物输入量的是什么关系?
3) 创新研究方法和研究技术。全球变化对凋落物理化性质的影响存在短期应激反应和长期适应的过程。探究凋落物对不同时间尺度如何响应和适应,需要设立定位试验,积累长期数据验证。目前研究手段的局限性,地下凋落物的分解过程,特别是地下凋落物对根系生物量、根际微生物的作用机制还不清楚,需要借助微根窗等技术为根系生长、根系周转、根系分解的研究提供可视化的手段。同时,利用碳同位素示踪法可区分外源性碳输入和土壤原有碳输入。
[1] Schiesinger W H. Evidence from chronosequence studies for a low carbon-storage potential of soils[J].Nature, 1990, 348(6298): 232–234
[2] Dixon R K, Solomon A M, Brown S, et al. Carbon pools and flux of global forest ecosystems[J]. Science, 1994, 263(5144): 185–190
[3] 彭少麟, 李跃林, 任海, 等.全球变化条件下的土壤呼吸效应[J]. 地球科学进展, 2002, 17(5): 705–713
[4] Raich J W,Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate[J]. Tellus,1992, 44(2): 81–99
[5] 刘绍辉, 方精云. 土壤呼吸的影响因素及全球尺度下温度的影响[J]. 生态学报, 1997, 17(5): 469–476
[6] 陈全胜, 李凌浩, 韩兴国, 等. 水分对土壤呼吸的影响及机理[J]. 生态学报, 2003, 23(5): 972–978
[7] Gome-casanovas N, Matamala R, Cook D R, et al. Net ecosystem exchange modifies the relationship between the autotrophic and heterotrophic components of soil respiration with abiotic factors in prairie grasslands[J]. Global Change Biology, 2012, 18(8): 2532–2545
[8] 王忆慧, 龚吉蕊, 刘敏, 等. 草地利用方式对土壤呼吸和凋落物分解的影响[J]. 植物生态学报, 2015, 3(1): 239–248
[9] Zhang Z, Duan J, Wang S, et al. Effects of land use and management on ecosystem respiration in alpine meadow on the Tibetan plateau[J]. Soil & Tillage Research, 2012, 124(4): 161–169
[10] Liski J, Perruchoud D, Karjalainen T. Increasing carbon stocks in the forest soils of western Europe[J]. Forest Ecology & Management, 2002, 169(1/2): 159–175
[11] 张东秋, 石培礼, 张宪洲. 土壤呼吸主要影响因素的研究进展[J]. 地球科学进展, 2005, 20(7): 778–785
[12] 宋新章, 江洪, 张慧玲, 等. 全球环境变化对森林凋落物分解的影响[J]. 生态学报, 2008, 28(9): 4414–4423
[13] 郑元润, 周广胜, 张新时, 等. 中国陆地生态系统对全球变化的敏感性研究[J]. Acta Botanica Sinica, 1997(9): 837–840
[14] Delucia E H, Hanmilton A, Milton J G, et al. Net primary production of a forest ecosystem with experimental CO2enrichmen[J]. Science, 1999, 284(5417): 1177
[15] Schlesinger W H, Anderws J A. Soil respiration and the global carbon cycle[J]. Biogeochemistry, 2000, 48(1): 7–20
[16] Eliasson P E, Mcmurie R E, Pepper D A, et al. The response of heterotrophic CO2flux to soil warming[J]. Global Change Biology, 2005, 11(1): 167–181
[17] 邓琦, 刘世忠, 刘菊秀, 等. 南亚热带森林凋落物对土壤呼吸的贡献及其影响因素[J]. 地球科学进展, 2007, 9(22): 976–986
[18] 骆土寿, 陈步峰, 李意德, 等. 海南岛尖峰岭热带山地雨林土壤和凋落物呼吸研究[J]. 生态学报, 2001, 21(12): 2013–2017
[19] 陈四清, 崔骁勇, 周广胜, 等. 内蒙古锡林河流域大针茅草原土壤呼吸和凋落物分解的 CO2排放速率研究[J].植物学报(英文版), 1999, 41(6): 645–650
[20] Kim H, Hirano T, Koike T, et al. Contribution of litter CO2production to total soil respiration in two different deciduous forest[J]. Phyton, 2005, 45(4): 385–388
[21] 陈宝玉, 刘世荣, 葛剑平, 等. 川西亚高山针叶林土壤呼吸速率与不同土层温度的关系[J]. 应用生态学报, 2007, 18(6): 1219–1224
[22] 王娓, 郭继勋. 东北松嫩平原羊草群落的土壤呼吸与枯枝落叶分解释放CO2贡献量[J]. 生态学报, 2002, 22(5): 655–660
[23] 杨继松, 刘景双, 孙丽娜. 三江平原草甸湿地土壤呼吸和枯落物分解的 CO2释放[J]. 生态学报, 2008, 28(2): 805–810
[24] 王丽丽, 宋长春, 葛瑞娟, 等. 三江平原湿地不同土地利用方式下土壤有机碳储量研究[J]. 中国环境科学, 2009, 29(6): 656–660
[25] Sayer E J, Power J S, Tanner E V. Increased litterfall in tropical forests boosts the transfer of soil CO2to the atmosphere[J]. PLoS ONE, 2007, 2: e1299
[26] Sayer E J, Tanner E V, Chesman A. Increased litterfall changes fine root distribution in a moist tropical fores[J]. Plant and Soil, 2006, 281(1): 5–13
[27] 郭剑芬, 杨玉盛, 陈光水, 等. 森林凋落物分解研究进展[J]. 林业科学, 2006, 42(4): 93–100
[28] Aerts R, Caluwe H D. Nutritional and plant—mediated controls on leaf litter decomposition of Carex species[J]. Ecology, 1997, 78(1): 244–260
[29] Kuzyakov Y. Priming effects: Interactions between living and dead organic matter[J]. Soil Biology & Biochemistry, 2010, 42(9): 1363–1371
[30] Sayer E J, Heard M S, Grant H K, et al. Soil carbon release enhanced by increased tropical forest litterfall[J]. Nature Climate Change, 2011, 1(6): 304–307
[31] Taylor B R, Parkinson D, Parsons W F. Nitrogen and Lignin content as predictors of litter decay rates: a microcosm tes[J]. Ecology, 1989, 70(1): 97–104
[32] Moore T, Trofymow J, Taylor B R, et al. Litter decomposition rates in Canadian forests[J]. Global Change Biology, 1999, 5(1): 75–82
[33] Cotrufo M, Ineson P. Elevated CO2reduces field decomposition rates of Betula pendula (Roth.) leaf litter[J]. Oecologia,1996,106(4): 525–530
[34] Frey S D, Knorr M, Parrent J L, et al. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests[J]. Forest Ecology Management, 2004, 196: 159–171
[35] Waldrop M P, Firestone M K. Microbial community utilization of recalcitrant and simple carbon compounds: Impact of oak-woodland plant communities[J]. Oecologia, 2004, 138(2): 275–84
[36] Phillips R L, Zak D R, Holmes W E, et al. Microbial community composition and function beneath temperate trees exposed to elevated atmospheric carbon dioxide and ozone[J]. Oecologia, 2002, 131(2): 236–244
[37] 张池, 陈旭飞, 周波, 等. 蚓粪施用对土壤微生物特征以及酶活性的影响[J]. 土壤, 2014(1): 70–75
[38] Brant J B, Sulzman E W, Myrold D. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation[J]. Soil Biology Biochemistry, 2006, 38(8): 2219–2232
[39] Nadelhoffer K J, Boone R D, Canary J D, et al. The DIRT experiment: litter and root influences on forest soil organic matter stocks and function[M]//Foster D, Aber J. Forests in time: The environmental consequences of 1000 years of change in New England. Newhaven and London: Yale University Press, 2004: 300–315
[40] Subke J A, Volker H, Giovanna B, et al. Feedback interactions between litter decomposition and rhizosphere activity[J]. Oecologia, 2004, 139(4): 551–559
[41] Li Y, Xu M, Sun O J, et al. Effects of root and litter exclusion on soil CO2efflux and microbial biomass in wet tropical forests[J]. Soil Biology Biochemistry, 2004, 36(12): 2111–2114
[42] Vasconcelos S S, Zarin D J, Capanu M, et al. Moisture and substrate availability constrain soil trace gas fluxes in an eastern Amazonian regrowth forest[J]. Global Biogeochemical Cycles, 2004,18(2): 193–204
[43] Zimmermann M, Meir P, Bird M, et al. Litter contribution to diurnal and annual soil respiration in a tropical montane cloud forest[J]. Soil Biology Biochemistry, 2009, 41(6): 1338–1340
[44] 王春燕, 陈秋波, 袁坤, 等. 橡胶林土壤呼吸速率及其与土壤温湿度的关系[J]. 土壤学报, 2013, 50(5): 974–982 [45] 周存宇, 周国逸, 张德强, 等. 鼎湖山森林地表CO2通量及其影响因子的研究[J]. 中国科学 D 辑, 2004(Z2): 175–182
[46] 田祥宇, 涂利华, 胡庭兴, 等. 华西雨屏区苦竹人工林土壤呼吸各组分特征及其温度敏感性[J]. 应用生态学报, 2012, 23(2): 293–300
[47] Rey A, Pegoraro E, Tedeschi V, et al.Annual variation in soil respiration and its components in a coppice oak forest in Central Italy[J]. Global Change Biology, 2002, 8(9): 851–866
[48] 刘尚华, 吕世海, 冯朝阳, 等. 京西百花山区六种植物群落凋落物及土壤呼吸特性研究[J]. 中国草地学报, 2008, 30(1): 78–86
[49] Jerome N, Daniel E, Claude B, et al. Estimating the contribution of leaf litter decomposition to soil CO2efflux in a beech forest using13C-depleted litte[J]. Global Change Biology, 2005, 11(10): 1768–1776
[50] 周小刚, 郭胜利, 车升国, 等. 黄土高原刺槐人工林地表凋落物对土壤呼吸的贡献[J]. 生态学报, 2012, 32(7): 2150–2157
[51] Sulzman E W, Brant J B, Bowden R D, et al. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2efflux in an old growth coniferous forest[J]. Biogeochemistry, 2005, 73(1): 231–256
[52] Bowden R D, Nadelhoffer K J, Boone R D, et al. Contributions of aboveground litter, belowground litter, and root respiration to total soil respiration in a temperate mixed hardwood forest[J]. Canadian Journal Forest Research, 2011, 23(7): 1402–1407
[53] Atarashi M, Korarashi J, shizuka A S, et al.Seasonal patterns and control factors of CO2effluxes from surface litter,soil organic carbon and root-derived carbon estimated using radiocarbon signatures[J]. Agricultural and Forest Meteorology, 2012, 152(1): 149–158
[54] 熊莉, 徐振锋, 杨万勤, 等. 川西亚高山粗枝云杉人工林地上凋落物对土壤呼吸的贡献[J]. 生态学报, 2015(14): 4678–4686
[55] 陆彬, 王淑华, 毛子军, 等. 小兴安岭4种原始红松林群落类型生长季土壤呼吸特征[J]. 生态学报, 2010, 30(15): 4065–4074
[56] Raich J W, Nadelhoffer K J. Belowground carbon allocation in forest ecosystems—Global trends[J]. Ecology, 1989, 70(5): 1346–1354
[57] 屠梦照, 姚文华. 鼎湖山南亚热带常绿阔叶林凋落物的特征[J]. 土壤学报, 1993(1): 34–42
[58] Schiesinger W H. Carbon balance in terrestrial detritus[J]. Annual Review Ecology System, 1977, 8(4): 51–81
[59] 杨万勤, 王开运. 森林土壤酶的研究进展[J]. 林业科学, 2004, 40(2): 152–159
[60] Wang Y, Hu Y, Ji B, et al. An investigation on the relationship between emission/uptake of greenhouse gases and environmental factors in semiarid grassland[J]. Advances in Atmospheric Sciences , 2003, 20(1): 119–127 [61] Schjonning P, Thomsen I K, Moldrup P, et al. Linking soil microbial activity to water- and air-phase contents and diffusivities[J]. Soil Science Society of America Journal, 2003, 67: 156–165
[62] Stark J M, Firestone M K. Mechanisms for soil-moisture effects on activity of nitrifying bacteria[J]. Applied and Environmental Microbiology, 1995, 61: 218–221
[63] Fioretto A, Papa S, Curcio E, et al. Enzyme dynamics on decomposing leaf litter of Cistus incanus, and Myrtus communis, in a Mediterranean ecosystem[J]. Soil Biology & Biochemistry, 2000, 32(13): 1847–1855
[64] Chen G S, Yang Y S, Guo J F, et al. Relationships between carbon allocation and partitioning of soil respiration across world mature forests[J]. Plant Ecology, 2010, 212(2): 195–206
[65] 田兴军, 立石贵浩. 亚高山针叶林土壤动物和土壤微生物对针叶分解的作用[J]. 植物生态学报, 2002, 26(3):257–263
[66] 沈海龙, 沈国舫. 樟子松人工林下针阔叶凋落物分解动态[J]. 林业科学, 1996(5): 393–402
[67] 林丽莎, 韩士杰, 王跃思, 等. 长白山四种林分土壤CO2释放通量的研究[J]. 生态学杂志, 2004, 23(5): 42–45
[68] Liang N, Hirano T, Zheng Z M, et al. Soil CO2efflux of a larch forest in northern Japan[J]. Biogeosciences & Discussions, 2010, 7(11): 3447–3457
[69] 孙轶, 魏晶, 吴钢, 等. 长白山高山冻原土壤呼吸及其影响因子分析[J]. 生态学杂志, 2005, 24(6): 603–606
[70] Kirschbaum M U F. The temperature dependence of soil organic matter decomposition, and the effect of global warming on soil organic C storage[J]. Soil Biology Biochemistry, 1995, 27(6): 753–760
[71] Hendricks J J, Nadelhoffer K J, Aber J D. Assessing the role of fine roots in carbon and nutrient cycling[J]. Trends Ecology Evolution, 1993, 8(5): 174–178
[72] Klopatek J M. Belowground carbon pools and processes in different age stands of Douglas-fir[J]. Tree Physiology, 2002, 22(2/3): 197–204
[73] Davidson E A, Savage K, Bolstad P. Belowground carbon allocation in forests estimated from litterfall and IRGA-based soil respiration measurements[J]. Agricultural & Forest Meteorology, 2002, 113(1): 39–51.
[74] Jipp P H, Nepstad D C, Cassel D K, et al. Deep soil moisture storage and transpiration in forests and pastures of seasonally-dry Amazonia[J]. Climatic Change, 1998, 39(2): 395–412
[75] 杨国亭, 宋关玲, 高兴喜. 外生菌根在森林生态系统中的重要性(Ⅰ)──外生菌根对宿主树木的影响[J]. 东北林业大学学报, 1999(6): 72–42
[76] Vogt K A, Moore E E, Vogt D J, et al. Conifer fine root and mycorrhizal root biomass within the forest floors of Douglas-fir stands of different ages and site productivities[J]. Canadian Journal of Forest Research, 1983, 13(3): 429–437
Contribution of Litters to Soil Respiration : A Review
LV Fucheng1,2, WANG Xiaodan1*
(1 Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China)
Soil respiration is the main approach for the output of soil carbon pool, and litter is an important factor to affect soil respiration. It is vital to understand the effect of litter decomposition process on soil respiration, because carbon budget among plant-soil-atmosphere carbon pool plays a key role in terrestrial carbon circle. This paper illuminated the contribution mechanism of litter to soil respiration, reviewed the contribution rates of aboveground litters and belowground litters of different plants to soil respiration. In addition, it also pointed out that the future study fields for understanding further the responses of terrestrial ecosystem to global climate change.
Litter; Soil respiration; Carbon cycle
S154.4
A
10.13758/j.cnki.tr.2017.02.003
国家自然科学基金项目(41571205)资助。
* 通讯作者(wxd@imde.ac.cn)
吕富成(1991—),男,河南安阳人,硕士研究生,主要从事土壤生态学研究。E-mail: lvfc2010@163.com
